WWW.BGBUNICT.IT
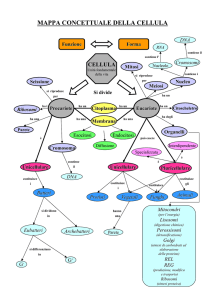
ORGANIZZAZIONE CELLULARE
PROCARIOTICA
ED EUCARIOTICA
TEORIA CELLULARE
(Virchow
Virchow;; Schleiden
Schleiden;; Schwann
Schwann.. 1850)
1) Tutti gli organismi sono formati da una o più cellule;
2) La cellula è la più piccola unità che presenta le
proprietà tipiche della vita;
3) Le cellule si moltiplicano solo in seguito a crescita e
divisione di cellule preesistenti.
A FRONTE DI UNA NOTEVOLE
VARIETA’ DI FORME E FUNZIONI,
DIFFERENTI TIPI DI CELLULE
SONO ACCOMUNATI DAL
POSSEDERE TUTTE:
1) CITOPLASMA
CITOPLASMA;;
2) NUCELOIDE O NUCLEO;
NUCLEO;
3) MEMBRANA PLASMATICA
LA MICROSCOPIA
SE SI RADDOPPIA IL DIAMETRO DI UNA CELLULA, IL SUO VOLUME DIVENTERA’ 8 VOLTE
MAGGIORE, MENTRE LA SUPERFICIE AUMENTERA’ DI APPENA 4 VOLTE!
VOLUME DELLA CELLULA = CONTIENE TUTTE LE SOSTANZE NECESSARIE PER FAR AVVENIRE
LE REAZIONI BIOCHIMICHE ALL’INTERNO DELLA STESSA CELLULA.
SUPERFICIE DELLA CELLULA = CONSENTE LO SCAMBIO DI SOSTANZE TRA AMBIENTE EST. ED
INT. E VICEVERSA.
CONCLUSIONE:
LE CELLULE DEVONO MANTENERE LE DIMENSIONI DEL LORO DIAMETRO
ENTRO UN CERTO LIMITE, ALTRIMENTI CI SAREBBE UNA ECCESSIVA DISCREPANZA
TRA SUPERFICIE (QUANTITA’ DI SOSTANZE CHE POSSONO SCAMBIARSI) E VOLUME
(QUANTITA’ DI REAZIONI BIOCHIMICHE CHE POSSONO SVOLGERSI)!
CELLULA
PROCARIOTICA
DOMINIO
PROCARIOTI
SOTTODOMINI
EUBATTERI
GRAM +
GRAM -
ARCHEOBATTERI
3 mm
EUBATTERI
PEPTIDOGLICANO: MATRICE DI CARBOIDRATI
LEGATI TRASVERSALMENTE DA CORTE UNITà
POLIPEPTIDICHE
INVAGINAZIONE DELLA MEMBRANA
IL FLAGELLO
CARATTERISTICHE PRINCIPALI DEGLI EUBATTERI
1) DIMENSIONI RELATIVAMENTE PICCOLE (POCHI mm DI LUNGHEZZA);
2) PARETE CELLULARE;
3) CAPSULA;
4) MEMBRANA PLASMATICA;
5) CROMOSOMA (ED EVENTUALMENTE UNO O Più PLASMIDI);
6) RIBOSOMI;
7) MESOSOMI;
8) FIMBRIE, PILI, FLAGELLI;
9) TALORA POSSONO SPORULARE.
IMP:: NON HANNO UN SISTEMA ENDOMEMBRANOSO!
IMP
ARCHEA
MANCANO DI PEPTIDOGLICNI
NELLA PARETE CELL
CELL.,
., LA MEMBR.
CELL. E’ COSTITUITA DA LIPIDI
DIFFERENTI RISPETTO A QUELLI
DEGLI EUBATTERI, LA STRUTTURA
DEI LORO GENI, NONCHE’ LA
SEQUENZA DI MOLTI LORO
RNA E PROTEINE E’ PIU’ SIMILE
A QUELLA DEGLI EUCARIOTI
PIUTTOSTO CHE A QUELLA DEGLI
EUBATTERI.
Qualcuno guarda alla
possibilità di usare sostanze
sintetizzate dagli Archea, gli
archeobatteri, che insieme ai
comuni procarioti e agli
eubatteri costituiscono una
delle classi principali di
organismi più primitivi
CELLULA
EUCARIOTICA
QUALI SONO GLI ORGANISMI AD
ORGANIZZAZIONE CELLULARE EUCARIOTICA?
COMPARTIMENTALIZZAZIONE
TUTTI GLI ORGANISMI
MULTICELLULARI, (COMPRESI
VEGETALI, FUNGHI ED
ANIMALI) SONO COSTITUITI
DA CELLULE EUCARIOTICHE.
ESISTONO CMQ. ALCUNI
ORGANISMI EUCARIOTI
UNICELLULARI (PROTOZOI,
AMEBE E LIEVITI)
COMPARAZIONE TRA ALCUNE CARATTERISTICHE DELLE CELL.
PROCARIOTICHE ED EUCARIOTICHE
CARATTERISTICA
PROCARIOTI
EUCARIOTI
Dimensione
Pochi mm
Decine di mm*
Nucleo delimitato da
Membrana
No
Si
Organuli
No
Si
Microtubuli
No
Si
Microfilamenti
No
Si
Filamenti intermedi
No
Si
Esocitosi ed Endocitosi
No
Si
Modal. Divisione Cell.
Scissione
Mitosi e Meiosi
Informazione Genetica
DNA + Proteine
DNA + Proteine
(Cromosomi)
Maturazione dell’RNA
Scarsa
Elevata
Ribosomi
Piccoli (70S)
Grandi (80S)
•* Le cellule eucariotiche sono ca. 10 volte più grandi delle procariotiche come
•Dimensioni lineari e 1000 volte maggiori come volume.
Membrane Biologiche
5-8 nm
LA MEMBRANA PLASMATICA DEGLI EUCARIOTI MOSTRA STRUTTURA E
FUNZIONI PARAGONABILI ALLE MEMBRANE CHE DELIMITANO I VARI
ORGANULI CELLULARI, A MENO DI FINI DIFFERENZE NELLA COMPOSIZIONE
CHIMICA (PROTEICA SOPRATTUTTO).
LA STORIA DELLA SCOPERTA
DELLA STRUTTURA DELLA MC
MODELLO A MOSAICO FLUIDO
(Singer e Nicolson
Nicolson,, 1972)
LA COMPONENTE LIPIDICA DI MEMBRANA
I PIU’ COMUNI FOSFOGLICERIDI
DI MEMBRANA
COINVOLTO IN MECCANISMI DI TRASDUZIONE
DEL SEGNALE
GLI SFINGOLIPIDI
SFINGOSINA AL POSTO DEL GLICEROLO!
SFINGOSINA E CERAMIDI SONO GLI UNICI SFINGOLIPIDI
DI MEMBRANA CHE NON CONTENGONO CARBOIDRATI
CEREBROSIDI E GANGLIOSIDI SONO ABBONDANTI
NELLE MAMBRANE DELLE CELLULE NERVOSE
DEGLI ANIMALI
NELLE MEMBRANE CELLULARI DI
BATTERI E CIANOBATTERI GLI STEROLI
SONO SOSTITUITI DA MOLECOLE SIMILI:
GLI OPANOIDI
FLIPPASI
DIMOSTRAZIONE SPERIMENTALE DEI MOVIMENTI LATERALI
DELLE PROTEINE E DEI LIPIDI
FRYE L.D. AND EDININ M.A. (1970)
I RAFT NON INVAGINATI E LE CAVEOLE
DELLA MEMBRANA PLASMATICA
MICRODOMINI LIPIDICI
MENO FLUIDI.
SONO PRESENTI ALTE
CONCENTRAZIONI DI
COLESTEROLO,
GLICOSFINGOLIPIDI(PRESENTANO
CODE DEGLI ACIDI
GRASSI Più LUNGHE E
SATURE).
SONO ASSOCIATE ALLA
PRESENZA DI RECETTORI DI
MEMBRANA
LA FASE LIQUIDO-ORDINATA (LO), INTERMEDIA TRA LA FASE CRISTALLINA
E LIQUIDO-CRISTALLINA
I RAFT LIPIDICI SVOLGONO
RUOLI CHIAVE IN:
i)
ii)
iii)
Traffico di membrana;
Definizione di polarità
cellulare;
Trasduzione del segnale
LA COMPONENTE PROTEICA DI MEMBRANA
i) Proteine integrali (spesso canali)
ii) Proteine periferiche (spesso proteine della MEC o trasduttori segnale)
NOTA: LEGAMI DEBOLI TRA PROTEINE PERIFERICHE (ESTRINSECHE
E TESTE POLARI DEI FOSFOLIPIDI)
iii) Proteine ancorate a lipidi (spesso implicate nella trasduzione del segnale. Es.:Ras e Src
Src))
LA COMPONENTE GLICIDICA DI MEMBRANA
I GLUCIDI DI MEMBRANA SONO RAPPRESENTATI DA CATENE
OLIGOSACCARIDICHE (DA 2 A 60 RESIDUI).
I GLUCIDI POSSONO LEGARSI COVALENTEMENTE SIA A PROTEINE
(GLICOPROTEINE), SIA A LIPIDI (GLICOLIPIDI).
SONO COINVOLTI IN MECCANISMI DI RICONOSCIMENTO
RECETTORE – LIGANDO E DI INTERAZIONE CELLULA-CELLULA
GLICOPROTEINE E GLICOLIPIDI
FORMANO UN RIVESTIMENTO
GLUCIDICO DETTO GLICOCALICE
GLICOCALICE,,
PARTCOLARMENTE EVIDENTE NEI
MICROVILLI INTESTINALI
ASIMMETRIA DELLA MEMBRANA PLASMATICA
IL VERSANTE CITOPLASMATICO
DELLA MEMBRANA HA
UNA CARICA NETTA NEGATIVA
GRAZIE ALLA MASSICCIA PRESENZA
DI FS
L’ASIMMETRIA DELLA MEMBRANA
VIENE STABILITA AL MOMENTO DELLA
SUA SINTESI, GRAZIE ALL’ATTIVITà
ALL’ATTIVITà
DELLE FLIPPASI
FLIPPASI..
Nucleo
DESCRITTO PER LA PRIMA VOLTA NEL 1831 DA ROBERT
BROWN. E’ L’ORGANULO CELL. PIU’ GRANDE E CARATTE=
RISTICO DELLE CELL EUCARIOTICHE. LA SUA DIMEN=
SIONE E’ DI POCHI mm.
Il poro nucleare
Passano dal complesso del poro:
RNA, proteine, e ribosomi (questi ultimi per trasporto attivo)
La lamina nucleare
LAMINA NUCLEARE:
NUCLEARE: sottile e densa rete di proteine adesa al
versante nucleoplasmatico della membrana interna del
nucleo. Svolge funzione di sostegno ed è il sito di attacco
per i telomeri dei cromosomi durante la meiosi.
LAMìNE A E C SONO OMOLOGHE AI FILAMENTI INTERMEDI
DEL CITOSCHELETRO. SI AGGREGANO IN LUNGHE FIBRE
A COSTITUIRE I FILAMENTI DELLA LAMINA.
MUTAZIONI A CARICO DEI GENI DELLE LAMìNE POSSONO
PROVOCARE MALATTIE EREDITARIE COME UNA RARA
FORMA DI DISTROFIA MUSCOLARE.
La matrice nucleare
Matrice fibronucleare insolubile
che costituisce il nucleoscheletro.
Coinvolta nel mantenimento della
forma del nucleo.
Costituisce l’impalcatura a cui si
associano le fibre di cromatina e
funge da sistema di ancoraggio
per alcuni complessi molecolari
coinvolti nella duplicazione e nella
trascrizione.
Cromatina e Cromosomi
RIF.. A LEZIONE SPECIFICA
RIF
Nucleolo (Robert Perry, inizi 1960)
REGIONE DI POCHI μm DI DIAMETRO
SPESSO DI FORMA SFERICA
SEDE “SPECIALIZZATA” DEL NUCLEO,
CHE COMPRENDE rDNA E RISPETTIVI
TRASCRITTI, NONCHE’ SEDE DI
BIOGENESI DEI RIBOSOMI
REGIONE FIBRILLARE (rDNA o organizzatore
nucleolare + RNA POL I + TFs
TFs))
REGIONE GRANULARE (particelle
ribosomali a vari stadi di assemblaggio)
NOR (ORGANIZZATORE NUCLEOLARE)
The synthesis of ribosomal RNA is stimulated and after a
period of time, the DNA from the nucleolar organizing region
begins to look like a Christmas tree. The top of the tree is the
start site. As you move down the "tree", the branches appear
longer. Each branch is a growing strand of ribosomal RNA.
Taken from Bloom and Fawcett, A Textbook of Histology, Chapman
and Hall, 1994, Figure 1-17
NUCLEOPLASMA
IL NUCLEOPLASMA E’ ORGANIZZATO IN DOMINI FUNZIONALI DEFINITI
SPECKLE: CLUSTER DI GRANULI DI INTERCROMATINA, CONTENENTI
SPECKLE:
AGGREGATIDI COMPLESSI RIBONUCLEOPROTEICI ED ALTRI FATTORI
COINVOLTI NELLA MATURAZIONE DELL’mRNA
DELL’mRNA
I CORPI DI CAJAL
SONO STRUTTURE
COMPATTE, FUNZIONALMENTE
ASSOCIATE
ALLA MATURAZIONE DEI
TRASCRITTI ED AD ALTRE
ATTIVITA’ DEL DNA
NUCLEARE, ADIACENTI AGLI
SPECKLE
I componenti del corpo di Cajal sono fluttuanti e possono
fungere da sito di assemblaggio per i fattori di trascrizione e di
elaborazione dell'RNA.
Reticolo Endoplasmatico (1950/60)
Estesa rete interconnessa di canali membranosi e vescicole chiamate cisterne. Ogni
singola cisterna, racchiuso da un’unica membrana, contiene il lume della cisterna.
Reticolo Endoplasmatico Ruvido (RER)
RE ASSOCIATO A RIBOSOMI.
RIBOSOMI.
PRINCIPALE SEDE DI SINTESI DI PROTEINE DI SECREZIONE
SECREZIONE..
CELLULE AD INTENSA ATTIVITA’ SECRETORIA HANNO UN RER MOLTO ESTESO
(ES: CELLULE DEL PANCREAS ENDOCRINO).
LA MEMBR ESTERNA DEL RER E’ STRETTAMENTE CORRELATA, SIA STRUTTU=
RALMENTE CHE FUNZIONALMENTE CON L’INVOLUCRO NUCLEARE.
P.S.: I RIBOSOMI LIBERI NEL CITOSOL FONDAMENTAMENTALMENTE SINTETIZ=
ZANO PROTEINE CHE RIMANGONO ALL’INTERNO DELLA CELLULA.
LE PROTEINE RILASCIATE NEL LUME
DELLE CISTERNE SUBISCONO DELLE
MODIFICAZIONI POST-TRADUZIONALI.
N-GLICOSILAZIONE
IDROSSILAZIONE
ANCORAGGIO AI GLICOLIPIDI DI
MEMBRANA
Reticolo Endoplasmatico Liscio (REL)
LE SUE MEMBRANE NON SONO
ASSOCIATE AI RIBOSOMI.
SVOLGE ESSENZIALMENTE LA FUNZIONE
DI SINTETIZZARE LIPIDI CHE DIVENTE=
RANNO PARTE DELLE MEMBRANE
CELLULARI.
È ABBONDANTE NELLE CELLULE
ENDOCRINE CHE SINTETIZZANO ORMONI
STEROIDEI (TESTICOLO, FEGATO,
CELLULE INTESTINALI). IL REL E’
PERALTRO COINVOLTO IN MECCANISMI
DI DETOSSIFICAZIONE DA FARMACI
QUALI BARBITURICI, ANFETAMINE,
MORFINA, PESTICIDI, ERBICIDI,
ETC… CIO’ AVVIENE MEDIANTE
IDROSSILAZIONE DELLE MOLECOLE
XENOBIOTICHE
REL NELLE CELLULE MUSCOLARI: IL RETICOLO
SARCOPLASMATICO
NELLE CELLULE MUSCOLARI,
PER LA NECESSITA’ DI
ACCUMULARE E RILASCIARE
IL Ca++ SI è SVILUPPATO
UN ESTESO REL CHE PRENDE
NOME DI RETICOLO
SARCOPLASMATICO
REL E METABOLISMO DEI CARBOIDRATI: IL
CASO DELL’EPATOCITA
COINVOLTO NEL METABOLISMO
DEI CARBOIDRATI (FEGATO).
L’ENZIMA GLUCOSIOGLUCOSIO-6FOSFATASI CHE SI TROVA NELLE
MEMBRANE RE LISCIO
TRASFORMA IL GLUCOSIOGLUCOSIO-6FOSFATO (NON PUÒ
ATTRAVERSARE LA MEMBRANA
PLASMATICA ED ESSERE
RILASCIATO NEL CIRCOLO
SANGUIGNO) IN GLUCOSIO
(PUÒ ESSERE TRASPORTATO
FUORI DALLA CELLULA)
REL E DETOSSIFICAZIONE: IL CYT PP-450
EFFETTI “COLLATERALI” DELL’USO DEI
BARBITURICI
I RIBOSOMI (PALADE et al., 1960)
RIBOSOMI LIBERI: PROTEINE CHE
RIMANGONO NEL CITOPLASMA;
RIBOSOMI LEGATI ALLA MEMBRANA
DEL RER: PROTEINE CHE VENGONO
RACCHIUSE IN VESCICOLE (CHE
RIMANGONO NEL CITOSOL O CHE
VENGONO SECRETE ALL’ESTERNO
DELLA CELLULA)
Mitocondri (Rudolf Kolliker
Kolliker,, 1850 – Palade
Palade,,
1953)
SI TROVANO IN TUTTE LE CELLULE EUCARIOTICHE
(PROTISTI, FUNGHI, ANIMALI E VEGETALI)
Palade G,
The Anatomical Record
Volume 114, Issue
3, pages 427–
451, November 1952
Al loro interno avviene la Respirazione Cellulare.
Processo attraverso cui molecole “carburante”
(Glucidi, Lipidi) sono degradate da reazioni in parte
citosoliche ed in parte mitocondriali a CO2 ed
H2O con rilascio di energia, per circa il 43%
catturata dalle molecole di ATP.
Le molecole “carburante” vengono “bruciate”
grazie all’O2, ecco perché si parla di
respirazione!
Ciclo degli acidi tricarbossilici (matrice)
Fosforilazione ossidativa (membrana interna)
mtDNA UMANO: ca. 16.5 Kb
TUTTI I MITOCONDRI DI UNA SINGOLA CELLULA EUCARIOTICA SONO PRODOTTI
PER DIVISIONE DI MITOCONDRI PRE-ESISTENTI
IN BASE ALLE FUNZIONI CELLULARI, IL
NUMERO DEI MITOCONDRI PUÒ ESSERE
MOLTO ELEVATO, OPPURE POSSONO ESSERE
DISPOSTI IN MANIERA STRATEGICA
ALL’INTERNO DELLA CELLULA PER
ASSOLVERE AL FABBISOGNO DI ATP.
I mt POSSONO ESSERE ORGANIZZATI A FORMARE
UN’UNICA ESTESA RETE MITOCONDRIALE DINAMICA
TEORIA DELL’ENDOSIMBIONTE (Umberto Pierantoni
Pierantoni,, 1909;
Lynn Margulius
Margulius)) – 1,5 MLD. DI ANNI FA SI PASSAVA
DA UN’ATMOSFERA RIDUCENTE AD UNA OSSIDANTE
NASCITA DEI MITOCONDRI
PROVE A SOSTEGNO DELLA TEORIA DELL’ENDOSIMBIONTE
I)
II)
III)
IV)
V)
VI)
VII)
Doppia membrana;
Dimensioni;
Struttura e dimensione dei ribosomi mt
mt;;
Materiale genetico autonomo;
Divisione per scissione semplice;
Modalità di divisione del DNA, simile ai procarioti;
Sequenze nt degli mt rRNA simili a batteri
MITOCONDRI E APOPTOSI
PATOLOGIE MITOCONDRIALI
PRINCIPALMENTE ENCEFALOMIOPATIE (MUTAZIONI PRINCIPALMENTE A CARICO DELL’mtDNA
DELL’mtDNA))
Complesso del Golgi (Camillo Golgi, 1890)
Pila di cisterne membranose (da 3 a 8) cui non sono associati ribosomi.
Generalmente è posizionato in prossimità di complessi di membrane del RER,
in particolare tra questo e la membrana citoplasmatica.
All’interno del complesso avvengono delle modifiche post
post-traduzionali (aggiunta
di piccoli gruppi funzionali o di lipidi o di glucidi) delle proteine sintetizzate
dal RER.
Una volte modificate, le proteine gemmano dalla faccia del Golgi rivolta
verso la membrana cellulare.
cellulare.
Nelle cellule vegetali è noto come dittiosoma.
Le molecole che passano da un
fronte all’altro del Golgi vengono
modificate: glicosilazione
glicosilazione,,
solfatazione,, acetilazione
solfatazione
acetilazione,,
deamminazione (sintesi di
sfingomielina e glicolipidi).
glicolipidi). Nel
Golgi vengono sintetizzati
polisaccaridi complessi come i
glucosoamminoglicani (acido
ialuronico, si trova nella matrice
extracellulare delle cellule
animali)
i) MODELLO DELLE CISTERNE STAZIONARIE;
ii) MODELLO DI MATURAZIONE DELLE CISTERNE
Lisosomi (De Duve
Duve,, 1950)
Vescicole contenenti più di 30 enzimi idrolitici
necessari per la digestione di molte
molecole complesse (prot
(prot,, lipidi, ac. nucleici,
polisaccaridi).
Si originano per gemmazione di vescicole dal
complesso del Golgi.
Il pH al loro interno è di ca. 5.
Es. di loro funzionamento: le molecole alimentari che
entrano all’interno della
cellula mediante endocitosi. Le vescicole endocitiche
possono quindi fondersi
con il lisosoma che inizierà a degradare il contenuto
della vescicola.
Altri processi:
1) Autofagia (Rimozione di organelli difettosi);
2) Fagocitosi (Eliminazione di batteri o particelle
cellulari ad opera dei fagociti).
MALATTIE DA ACCUMULO LISOSOMALE (Es.
TayTay
-Sachs
Sachs))
IDROLASI ACIDE (AGISCONO PER LO Più A pH 5)
POMPA H+ (ATP
(ATP--DIPENDENTE)
POMPA DI CONTINUO H+ DAL
CITOSOL ALL’INTERNO DEL LISOSOMA
I LISOSOMI INATTIVI (LISOSOMI
(LISOSOMI
PRIMARI)) NON HANNO pH
PRIMARI
ACIDO FINO A QUANDO NON SI
FONDONO CON UN VACUOLO
ALIMENTARE. IN QUESTO CASO, LA
POMPA PROTONICA DIMINUISCE IL
pH (LISOSOMA
(LISOSOMA SECONDARIO,
SECONDARIO, ATTIVO).
I LISOSOMI INTERVENGONO IN
PROCESSI DI ETEROFAGIA E
AUTOFAGIA
MACROFAGI E NEUTROFILI UTILIZZANO I
LISOSOMI PER NEUTRALIZZARE BATTERI O
RESIDUI CELLULARI
IN RARISSIMI CASI SONO COINVOLTI IN
PROCESSI DI DIGESTIONE EXTRACELLULARE
(ACROSOMA SPERMATOZOI)
MALATTIE LISOSOMIALI DA “ACCUMULO”
PATOLOGIE RESPIRATORIE CAUSATE DALL’INALAZIONE DI
PARTICELLE DI SILICE (SILICOSI
SILICOSI) O ASBESTO (ASBESTOSI
ASBESTOSI) : ROTTURA
DEI LISOSOMI, RILASCIO DEGLI ENZIMI IDROLITICI, MORTE DELLA
CELLULA FAGOCITANTE. A CAUSA DELLA MORTE DI UN GRAN
NUMERO DI FAGOCITI, I FIBROBLASTI DEPOSITANO COLLAGENE E
RENDONO I POLMONI MENO ELASTICI, CAUSANDO COSÌ
INSUFFICIENZA RESPIRATORIA.
PEROSSISOMI (De Duve
Duve,, 1983)
I PEROSSISOMI SONO
“POSITIVI” ALLA REAZIONE
CON DAB
(DIAMMINOBENZIDINA).
CONTENGONO AL LORO
INTERNO ENZIMI
OSSIDATIVI IN GRADO DI
EFFETTUARE βOSSIDAZIONE DEGLI AC.
GRASSI A LUNGA CATENA ;
ENZIMI QUALI LA
LUCIFERASI
(CHEMIOLUMINESCENZA).
SONO ABBONDANTI NEL FEGATO E
NEI RENI DOVE SVOLGONO
UN’AZIONE DETOSSIFICANTE (ALCOL,
FORMALDEIDE, FENOLI, ECC….)
TALI ENZIMI FORMANO IONI
PEROSSIDO MEDIANTE
AGGIUNTA DI H AD H20 – LA
CATALASI A SUA VOLTA
NEUTRALIZZA H2O2
CONVERTENDOLA IN H20 E
O2
I PEROSSISOMI SI ORIGINANO
DA PEROSSISOMI PREPRE-ESISTENTI
PER SCISSIONE
SCISSIONE,, PREVIA LORO
ACCRESCIMENTO.
PEROSSISOMI “COLLABORANO” CON MITOCONDRI
E CLOROPLASTI NELLE CELLULE VEGETALI PER
LA FOTORESPIRAZIONE
FOTORESPIRAZIONE:
FOTORESPIRAZIONE:
ASSORBIMENTO FOTODIPENDENTE
DI O2 E RILASCIO DI CO2
DIFETTI NEGLI ENZIMI DEI PEROSSISOMI
SINDROME DI ZELLWEGER
ZELLWEGER:: PEROSSISOMI PRIVI DI ENZIMI, PUR
ESSENSO SINTETIZZATI NEL CITOSOL NON VENGONO
TRASPORTATI ALL’INTERNO DEGLI ORGANULI PER UN DIFETTO DI
RECETTORI O DI QUALCHE COMPONENTE DEL SISTEMA DI
TRASPORTO. (ANOMALIE NEUROLOGICHE, VISIVE E DEL FEGATO).
ADRENOLEUCODISTROFIA (ALD):
(ALD): MANCATA IMPORTAZIONE DI ACIDI
GRASSI A CATENA LUNGA DENTRO I PEROSSISOMI CHE SI
ACCUMULANO NEL SANGUE E NEI TESSUTI. NELLE CELLULE
NERVOSE PORTA AD UNA DISTRUZIONE DELLA GUAINA MIELINICA
ED ALLA ALTERAZIONE DEGLI IMPULSI NERVOSI
RICAPITOLANDO
Il Citoscheletro
Il citosch
citosch.. È un sistema di filamenti
e tubuli proteici interconnessi tra loro
Che si estende ovunque nel citoplasma
delle cell
cell.. eucariotiche
eucariotiche..
Molto importnate per dare sostegno e
forma alle cellule euc
euc.. Animali, molto
meno imp. Nelle cellule vegetali dove
la Parete Cellulare svolge un ruolo
chiave nel mantenimento della forma
e della strutt
strutt.. Cell
Cell..
LE FIBRE CITOSCHELETRICHE SONO DEI POLIMERI CHE SI COSTITUISCONO
A PARTIRE DA SUBUNITà PROTEICHE
MICROFILAMENTI DI ACTINA, FILAMENTI INTERMEDI, MICROTUBULI
OLTRE A QUESTE TRE STRUTTURE BASE, FANNO
PARTE DEL CITOSCHELETRO ANCHE ALTRE PROTEINE
(CA
CA.. 20), TRA LE QUALI LA MIOSINA DEL MUSCOLO
MICROFILAMENTI DI ACTINA
I MICROFILAMENTI SONO RESPONSABILI
DEL MANTENIMENTO E DEL
CAMBIAMENTO DELLA FORMA DELLA
CELLULA. RESISTONO ALLA
DEFORMAZIONE, ALLA TENSIONE O
ALLE FORZE DI TRAZIONE;
TRASMETTONO FORZA
E LIMITANO LA DIFFUSIONE DEGLI
ORGANELLI. ASSIEME AD ALTRE
PROTEINE COSTITUISCONO IL CORTEX,
UN RETICOLO AL DI SOTTO DELLA
MEMBRANA PLASMATICA
CHE RISULTA COSÌ RINFORZATA.
CIASCUNA MOLECOLA DI ACTINA
(MONOMERICA) LEGA UNA
MOLECOLA DI ATP CHE VIENE
IDROLIZZATO AD ADP DURANTE
LA POLIMERIZZAZIONE.
DIVERSE FORME DI ACTINA:
4 α-ACTINA MUSCOLO SPECIFICHE
2 β-ACTINA E 2 ϒ-ACTINA (IN CELLULE
DIFFERENTI)
POLIMERIZZAZIONE DEI MICROFILAMENTI DI ACTINA
VIDEO SINTESI F-ACTINA
MACROFAGO
lamellipodi sono delle ampie
strutture laminari;
I filopodi sono sottili proiezioni
citoplasmatiche simili ai
lamellipodi che si estendono dal
fronte di avanzamento della
cellule migranti.
Lo pseudopodio è
l'estroflessione
mobile di citoplasma
osservabile
in alcuni organismi unicellulari
(amebe, sporozoi, plasmodi,
ecc.)
Gli pseudopodi cellulari sono
estroflessioni della membrana
plasmatica tipici delle cellule
fagocitiche come macrofagi e
cellule dendritiche.
I MICROFILAMENTI
SUPPORTANO I MICROVILLI E
CONTRIBUISCONO ALLA
FORMAZIONE DEGLI
PSEUDOPODI.
L’ACTINA CONTRIBUISCE AL MOVIMENTO
IN DUE MODI:
i) POLIMERIZZANDO/DEPOLIMERIZZANDO LA
RETE DI FILAMENTI CORTICALI AL DI SOTTO
DELLA MEBRANA;
ii) ESTENDENDO FILOPODI, LAMELLIPODI E
PSEUDOPODI
VIDEO MOVIMENTO DENTRO LA CELLULA
MICROTUBULI
LEGAMI NON COVALENTI TRA I MONOMERI
αEβ
ESITONO DIVERSE FORME DI TUBULINA
α,β, δ,ε,ζ,η. OGNI MONOMERO DI TUBULINA
LEGA UNA MOLECOLA DI GTP CHE VIENE
IDROLIZZATA IN SEGUITO A
POLIMERIZZAZIONE
IN GENERE 13 PROTOFILAMENTI SI
DISPONGONO L’UNO ACCANTO ALL’ALTRO
PER FORMARE UN CILINDRETTO
MICROTUBULARE
POLARITA’
I microtubuli si formano a partire dal centro
della cellula irradiandosi verso la periferia.
L’estremità minus è ancorata al centro di
organizzazione del microtubulo
(centrosoma).
IL MOVIMENTO INTRACELLULARE È
ASSICURATO DA PROTEINE MOTRICI CHE
PERMETTONO IL MOVIMENTO DI COMPONENTI
SUBCELLULARI LUNGO LE STRUTTURE DEL
CITOSCHELETRO.
MOVIMENTO ANTEROGRADO (VERSO L’ESTREMITà
L’ESTREMITà ASSONE): KINESINE
MOVIMENTO RETROGRADO (VERSO IL CORPO CELLULARE): DINEINE
CIGLIA E FLAGELLI
FLAGELLI DI SOLITO: LUNGHI; SINGOLI (AL MAX IN COPPIA); MOVIMENTO
ONDULATORIO
CIGLIA DI SOLITO: CORTE; NUMEROSE; MOVIMENTO A BATTITO
UNIDIREZIONALE (BATTITO A REMO)
MOVIMENTO AMEBOIDE IMPORTANTE PER LA DIAPEDESI DEI GLOBULI
BIANCHI, DURANTE EVENTI INFETTIVI ED INFIAMMATORI
FILAMENTI INTERMEDI
DATA LA LORO TESSUTOTESSUTO-SPECIFICITà SONO IMPORTANTI
COME STRUMENTO CLINICOCLINICO-DIAGNOSTICO
OGNI SPECIFICO TIPO DI CELLULA POSSIEDE UNO DEI VARI
TIPI DI FILAMENTI INTERMEDI: LE CHERATINE (CELLULE
EPITELIALI), VIMENTINA (TESSUTO CONNETTIVO), DESMINA
(MUSCOLI), NEUROFILAMENTI (NEURONI).