BOTANICA
Le piante si sono evolute quasi sicuramente da alghe verdi del gruppo delle caroficee: si definisce
pianta un organismo eucariote, pluricellulare, fotosintetico adattato a vivere prevalentemente sulla
terraferma.
Quindi rispetto ad alcuni gruppi di alghe c'è una grande differenza soltanto per quanto riguarda il
substrato dove vivono. È per questo che certuni classificano le alghe nel regno Plantae.
MINI GLOSSARIO
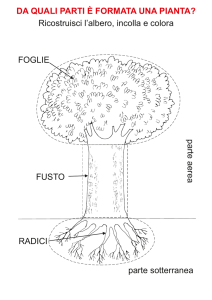
sistema radicale: il complesso normalmente ipogeo della pianta;
sistema aereo: il complesso normalmente epigeo della pianta, fusto foglie etc.
radici: funzione di ancoraggio e assorbimento di acqua e sali minerali;
cuticola: strato ceroso delle foglie con compito di evitare la traspirazione;
stomi: pori della pagina inferiore delle foglie deputati agli scambi gassosi;
tessuto vascolare: tessuti specializzati nel trasporto di acqua, minerali e linfa elaborata;
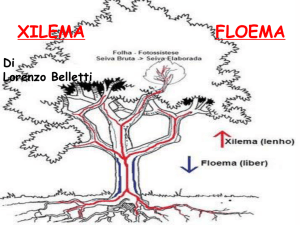
xilema: tessuto vascolare specializzato nel trasporto di acqua e sali minerali dalle radici alla
parte aere, costituito da cellule morte;
floema: tessuto vascolare specializzato nel trasporto di linfa elaborata dalla parte aerea
fotosintetizzante a tutti gli altri distretti, costituito da cellule vive;
gametangi: strutture formate da cellule idonee a proteggere le cellule che produrranno i
gameti;
embriofite: sono definite così le piante poiché esiste uno stadio embrionale pluricellulare
all'interno della pianta madre.
CLASSIFICAZIONE DELLE PIANTE
Briofite, licopodi e pteridofite → crittogame (a nozze nascoste);
Spermatofite: piante vascolari con semi nudi (gimnosperme), piante vascolari con semi
racchiusi in ovario (angiosperme) → fanerogame (a nozze manifeste).
Le denominazioni crittogame e fanerogame non hanno alcuna valenza sistematica, ma si usa per
comodità didattica.
Briofite: piante senza veri e propri tessuti vascolari. Sono probabilmente il termine di
passaggi dalle alghe alle piante. Crescono in ambienti umidi, costituiscono la vegetazione
tipica del bioma tundra, Il tallo (è il corpo delle briofite, è costituito quasi esclusivamente
dal gametofito, ha attività fotosintetica e misura in altezza non più di qualche centimetro) è
costituito da strutture, prive di tessuti di sostegno e di sistema vascolare, dette filloidi,
cauloidi e rizoidi. Il ciclo biologico presenta una evidente alternanza di generazioni con lo
stato di gametofito aploide dominante sullo sporofito diploide. Lo sporofito è costituito da
uno stelo da una capsula e da un piede che si inserisce nel gametofito e da esso dipende per
la nutrizione. Nella capsula si originano per meiosi le spore che germinano e producono il
protonema (primo gametofito) da cui per successive proliferazioni si ottiene il gametofito.
Su quest'ultimo si formano i gametangi maschili e femminili che portano i gameti. Dalla
fusione dei gameti deriva lo zigote quindi lo sporofito. Si classificano in: muschi, epatiche,
antocerotali.
Muschi: comprendono veri muschi e sfagni, i secondi molto frequenti nelle zone
paludose e spesso danno colorazioni rosse alle acque. I corpi degli sfagni quando
muoiono vanno a formare le torbe.
Epatiche: il nome deriva dal fatto che nel medioevo le proprietà medicamentose delle
essenze si potesse riconoscere dalla forma, in questo caso la forma del gametofito
pare un fegato e quindi ecco epatiche. Ci sono epatiche a foglie (tipiche delle zone
subtropicali) e epatiche a tallo, le più comuni appartengono al gen. Marchantia.
Licopodi e pteridofite: lo sporofito è un cormo (corpo della pianta) in cui si distinguono
radice, fusto e foglie. La foglia non ha solo il compito di fare la fotosintesi ma anche di
portare gli sporangi contenenti spore. Nel ciclo biologico è evidente l'alternanza di
riproduzione con netta predominanza dello stato di sporofito diploide sul gametofito. Dalla
germinazione delle spore si forma un gametofito laminare detto protallo sul quale si
producono i gametangi maschili (anteridi) e femminili (archegoni). I gameti si fondono
originano lo zigote, dal quale si origina l'embrione che prima riceve nutrimento dal
gametofito, poi quando si sviluppano le radici e le foglie diviene autosufficiente e il
gametofito degenera. I licopodi sono tipici di ambienti caldi tropicali. Le pteridofite si
suddividono in equiseti e felci.
Equiseti: si trovano facilmente nei fossi, nei terreni incolti, nei terreni paludosi.
Hanno stelo tubulare, e ramificazioni inserite ad anello intorno al fusto a più livelli,
separati da tratti senza ramificazioni, le cellule epidermiche sono fortemente
impregnate di silice e sono anche utilizzate per pulire i metalli (coda cavallina o
Equisetum arvense).
Felci: spontanee nei terreni ombreggiati dei climi temperati, ai tropici raggiungono i
15 m hanno foglie frondose espanse con venatura che si innalzano da fusti
sotterranei. I giacimenti di carbone odierni sono originati dalle antiche foreste di felci
arboree.
CICLI VITALI DELLE PIANTE
Le piante hanno alternanza di generazioni quindi si trovano individui diploidi detti sporofiti e
aploidi detti gametofiti. I gametofiti per mitosi produrranno i gameti maschili e femminili. Dalla
fecondazione verrà uno zigote diploide che dividendosi per mitosi produce lo sporofito.
Successivamente lo sporofito per meiosi produce spore aploidi che per mitosi danno il gametofito
aploide.
Nei muschi è dominante la fase del gametofito, nelle felci è dominante la fase dello sporofito, come
poi avviene per tutte le piante ancora più evolute.
Gimnosperme: sono piante legnose, caratterizzate da grande diversità morfologica. Possono essere
di varie dimensioni, da arbusti ad alberi giganteschi. Il legno omoxilo è costituito da elementi
chiamati tracheidi areolate che hanno funzione di trasporto e di sostegno. Hanno radici molto
robuste e profonde che presentano una ramificazione monopodiale, in cui l’asse principale è
superiore a quelli laterali. Hanno foglie di varia forma, a volte estese, più spesso aghiformi, con
abbondante cuticola, coriacee. Presentano un fiore sempre unisessuale. I microsporofilli (maschili),
in genere squame legnose che sostengono gli ovuli, generano per meiosi i gametofiti maschili o
granuli pollinici, in genere anemofili.
Da rilevare i tempi lunghi tra impollinazione e fecondazione e dispersione dei semi, dalla durata di
circa 2 anni.
Delle gimnosperme fanno parte 4 divisioni: Cycadophyta (palme), Ginkgophyta (Ginkgo biloba),
Coniferophyta e Gnetophyta (sono quelle più evolute, forse sono il termine di passaggio per le
angiosperme, hanno tessuti meccanici diversi da quelli conduttori, i fiori presentano accenni
rudimentali di calice e infiorescenze bisessuali, l'impollinazione avviene spesso ad opera di insetti,
presentano poche specie che vivono in ambienti tropicali).
Angiosperme: sono piante con fiore. Le specie classificate fino ad oggi sono circa 235.000.
Anche nelle angiosperme lo sporofito diploide domina il ciclo vitale. Questo sporofito protegge e
nutre il gametofito. I semi prodotti sono racchiusi in frutti che hanno la funzione di proteggerli e
favorirne la dispersione nell'ambiente. I fiori hanno strutture riproduttive particolari che si sono
evolute insieme agli animali impollinatori, co-evoluzione frutto di una azione selettiva basata su
reciproci vantaggi.
Il fiore è in realtà un fusto modificato munito di quattro serie di foglie modificate:
sepali: nella parte più bassa del fiore, in genere verdi;
petali: posti superiormente ai sepali, sono la parte appariscente e colorata, fungono in genere
da attrattori per animali impollinatori;
stame: struttura riproduttiva maschile costituita da un filamento che porta una sacca
chiamata antera che contiene i microsporangi con le sacche polliniche;
carpello: struttura riproduttiva femminile composta da stilo alla cui base si trova l'ovario che
contiene uno o più ovuli (megasporangi), all'apice dello stilo si trova lo stimma che cattura il
polline.
Il ciclo vitale di una angiosperma può essere così schematizzato:
Formazione per meiosi di spore aploidi che per mitosi originano i gametofiti maschili
(polline) e femminili (ovuli).
Impollinazione zoofila o anemofila: un granulo pollinico si deposita sulla estremità
appiccicosa dello stimma;
Formazione del tubulo pollinico che giunge fino alla cellula uovo e formazione dei due
nuclei spermatici;
Fecondazione del gamete maschile (un nucleo spermatico) con cellula uovo con formazione
dello zigote; l'altro nucleo spermatico si unisce al nucleo endospermico da cui si origina un
tessuto detto endosperma secondario con funzione di nutrire l'embrione; questo endosperma
è triploide poiché proviene dalla fusione di un nucleo spermatico e dai due nuclei polari
dell'ovario (in totale 8 nuclei: una oosfera, due sinergidi ai lati del polo micropilare, due
nuclei polari al centro, 3 antipodi al poco calazale).
Sviluppo del seme contenente un embrione diploide, quindi sporofito, circondato da tessuti
di riserva e da un tegumento;
Formazione del frutto per ispessimento ed evoluzione dei tessuti dell'ovario, in genere in
relazione alla dispersione e diffusione;
Germinazione del seme in condizioni idonee.
I motivi del successo evolutivo delle angiosperme sono:
relazioni di reciproca dipendenza tra animali impollinatori e piante;
rapidità di riproduzione rispetto alle gimnosperme (poche ore o giorni contro mesi o anni).
TIPI DI IMPOLLINAZIONE
Anemofila o anemogama: le specie che adottano questo mezzo, gimnosperme e molte
angiosperme, producono polline leggero e abbondante fiorendo a fine inverno, inizio primavera,
quando ancora la copertura fogliare è minima. Queste piante presentano fiori ridotti alle parti
essenziali o configurati in modo tale da disperdere facilmente il polline.
In generale le piante ad impollinazione anemofila sono caratterizzate da una elevata attitudine
pollinifera.
L'ulivo (Olea europaea), il noce (Juglans regia), il nocciolo (Corylus avellana), il pino (Pinus
spp.), la palma da dattero (Phoenix dactylifera), sono tipiche specie ad impollinazione anemofila
che producono non solo una grande quantità di fiori, ma anche un grande numero di granuli
pollinici, rispetto alle specie ad impollinazione entomofila, quali, ad esempio, le pomacee. Mentre
infatti una antera di pero contiene mille duemila granuli pollinici, una antera di ulivo ne può
contenere circa 10 mila ed una di nocciolo fino a 20 mila.
Nelle specie ad impollinazione anemofila, le infiorescenze maschili presentano una conformazione
idonea a facilitare la dispersione del polline nell’aria, mentre quelle femminili di solito prive di
organi vessillari (petali), hanno stimmi molto sviluppati per favorire la captazione del polline.
Nelle specie ad impollinazione anemofila il trasporto del polline può avere luogo anche a vari
chilometri del punto di provenienza, a seconda dell'intensità dei venti e di altri fattori influenti
(esempio piogge).
Idrofila o idrogama È l’impollinazione ad opera dell’acqua. È tipica delle piante acquatiche. Il
polline delle ninfee (Nymphaea alba), ad esempio, è liberato sotto il pelo dell’acqua, quello della
elodea (Elodea densa) galleggia sulla superficie, ma in entrambi i casi sono le correnti acquatiche
che lo trasportano fino ad incontrare lo stimma di un altro fiore della stessa specie.
Zoofila o zoogama: È l'impollinazione operata da animali come piccoli uccelli (ad es. colibrì) e
soprattutto insetti appartenenti a vari ordini (lepidotteri, coleotteri, ditteri e menotteri).
L’impollinazione ad opera degli insetti, entomogamia, è la più diffusa nel mondo ed esclusiva delle
zone temperate. Le piante entomofile hanno sviluppato strategie atte ad attrarre insetti specifici. In
genere le piante che presentano zoogamia hanno fiori profumati, colorati e dotati di nettare;
producono poco polline spesso appiccicoso e fioriscono in periodi stagionali o in momenti della
giornata favorevoli ai pronubi specifici.
Come detto nelle specie ad impollinazione entomofila il trasporto del polline è operato da numerosi
insetti, in primo luogo dalle api (Apis mellifera) che però possono essere affiancate anche da vari
altri pronubi. Il raggio medio di azione di tali pronubi si gira, in condizioni favorevoli, sui 700-900
metri, ma quello utile ai fini di una efficiente azione è di circa 400 metri. Nelle loro visite ai fiori le
api compiono un lavoro altamente specializzato: alcune operaie provvedono infatti a raccogliere il
polline che viene trasportato, agglomerato in grosse masse sugli arti posteriori, mentre altre operaie
raccolgono il nettare secreto dai fiori (il nettare è secreto al solo scopo di attirare gli impollinatori).
Visitando i fiori le api si coprono di polline (anche più di un milione di granuli) che trasportano
nelle successive visite sugli stimmi di altri fiori. Ogni ape visita in media, per ogni volo, una
settantina di fiori; poiché ogni colonia è composta da varie migliaia di api bottinatrici essa è così in
grado di assicurare l'impollinazione di un imponente numero di fiori.
Un'altra importante caratteristica che rende le api particolarmente preziose quali agenti pronubi, è
rappresentata dalla cosiddetta costanza fiorale. Di solito le api che iniziano la loro attività sui fiori
di una determinata specie, continuano a visitare solo i fiori di tale specie fino al termine della antesi,
per passare poi su fiori di altre specie; in tal modo si realizza una provvidenziale economia
nell'attività dei pronubi che limitano il loro campo d'azione alle piante appartenenti alla medesima
specie e quindi potenzialmente suscettibili di avvantaggiarsi della impollinazione incrociata.
D’altra parte,quando differenti specie fioriscono contemporaneamente i pronubi sono di preferenza
attratti da talune di esse soprattutto in rapporto al grado zuccherino del nettare secreto dai loro fiori,
così che ad esempio i fiori di ciliegio sono preferiti a quelli di pero (Pyrus communis).
Le condizioni climatiche che si verificano nel periodo utile per la impollinazione influiscono
notevolmente sulla attività delle api, che raggiungono la loro massima efficienza nelle giornate
soleggiate, con una temperatura media di 20-22°C. Al di sotto di 14°C non è giudicata proficua ai
fini della impollinazione. Quando il vento raggiunge una velocità di 20 km/h i voli si riducono
sensibilmente per cessare del tutto quando la velocità giunga a 35-40 km/h. I voli sono anche
notevolmente ostacolati dalle piogge.
FRUTTIFICAZIONE, MATURAZIONE DEI FRUTTI, DISSEMINAZIONE
Un altro processo che avviene in seguito alla fecondazione della cellula uovo, che si realizza
simultaneamente alla formazione del seme, è la formazione del frutto.
Il frutto comincia a formarsi (fase di accrescimento) quando la parete dell'ovario si ingrossa in
seguito all'impollinazione. Piccole quantità di auxine, portate dal polline, stimolano l'inizio della
crescita e con rapide divisioni cellulari che aumentano il numero di cellule nei tessuti ovarici.
Quando le mitosi cessano, il numero delle cellule resta costante e la crescita del frutto avviene
quindi per distensione cellulare. Alcune cellule moltiplicano il proprio volume anche più di una
decina di volte, altre vengono schiacciate e scompaiono. Durante questo periodo il frutto è duro, di
sapore di sgradevole per la presenza di tannini, acidi, alcaloidi, è di colore verde, coperto spesso da
peli e da squame: queste caratteristiche hanno la funzione di proteggere i frutti quando i semi sono
immaturi.
All’accrescimento prosegue la fase di maturazione accompagnata da notevoli mutamenti
biochimici. Quando i frutti cominciano a maturare compare un ormone, l'etilene che scatena una
serie di processi: l'amido viene degradato in zuccheri più semplici, la clorofilla viene sostituita da
pigmenti, come i carotenoidi o le antocianine; questi conferiscono al frutto colori attraenti quali
rosso arancio, viola; i peli e le squame cadono. Scompaiono quindi gli acidi e le sostanze di sapore
sgradevole; il pH aumenta, mentre si liberano sostanze aromatiche che conferiscono il profumo
caratteristico. Si verifica anche un rammollimento della polpa, dovuto alla degradazione delle
sostanze pectiche della parete cellulare e allo sviluppo di spazi intercellulari. La finalità intrinseca
della maturazione è quella di rendere frutto attraente agli animali, i quali, cibandosene, favoriscono
la dispersione e la diffusione dei semi. I cambiamenti visibili fanno seguito ad un incremento
improvviso del tasso respiratorio (respirazione climaterica), la velocità respiratoria aumenta fino ad
un massimo detto picco climaterico, dopodiché diminuisce.
Nella maturazione naturale si verifica la abscissione, cioè il distacco del frutto dalla pianta madre e
a questo punto avvengono ulteriori trasformazioni, tra le quali la fermentazione alcoolica che attira
gli animali che si cibano di frutti. Questa, chiamata fase di post-maturazione, è di ostacolo alla
prolungata conservazione dei frutti ed è utile in natura poiché così si verifica la completa
disorganizzazione del frutto e la liberazione del seme dai tegumenti che, avvolgendolo, ne
impediscono la maturazione. Un nuovo ciclo può quindi iniziare.
Le piante sono organismi immobili quindi, per diffondere i propri semi (disseminazione) lontano e
garantire la sopravvivenza della specie, attuano meccanismi diversi servendosi di agenti diversi,
come vento, acqua, animali.
Questi agenti, già visti nel ruolo di impollinatori, sono, in genere, anche disseminatori delle stesse
piante, e consentono la dispersione autonoma per mezzo di particolari dispositivi di lancio
(autodisseminazione). L’energia di lancio può essere dovuta a pressione di liquidi interni al frutto,
come nel caso del cocomero asinino (Ecballium elaterium), oppure a tensioni igroscopiche, come in
molte specie dei luoghi aridi.
La disseminazione anemocora è operata dal vento. I semi sono leggeri o di piccole
dimensioni, dotati di strutture atte al volo come ali o pappi.
La disseminazione idrocora avviene per mezzo dell’acqua, che trasporta semi galleggianti
per distanze notevoli.
La disseminazione zoocora è ad opera di animali ed avviene in due modi:
L’animale divora il frutto, in genere appetibile e colorato; il seme ben protetto, passa
indenne attraverso i succhi gastrici e viene eliminato con le feci.
L’animale trasporta il frutto, munito di uncini od altri elementi adesivanti, come
quelli del Gallium aparine attaccati al vello. L’abbandono del seme può avvenire in
luoghi lontani da quelli di partenza.
STRUTTURA E FUNZIONI DELLE PIANTE
Le angiosperme si possono classificare in due gruppi, in base al numero di foglioline embrionali e
ad altri caratteri:
Monocotiledoni:
Dicotiledoni:
foglie parallelinervie;
fasci vascolari a disposizione sparsa all'interno del fusto;
radici fascicolate.
foglie con nervature ramificate;
fasci vascolari con disposizione ad anello;
radici fittonanti.
TESSUTI MERISTEMATICI
I tessuti meristematici o meristemi sono tessuti privi di spazi intercellulari: sono formati da
cellule che hanno la capacità di dividersi in senso periclinale (parallelamente alla superficie) e in
senso anticlinale (perpendicolarmente alla superficie). La pianta si accresce in lunghezza grazie ai
meristemi primari o apicali e in larghezza grazie ai meristemi secondari o laterali
MERISTEMI PRIMARI O APICALI
Sono situati negli apici vegetativi, punti estremi del fusto e delle radici, e sono formati da cellule
poco differenziate, abbastanza piccole, con parete cellulare sottile, citoplasma denso e prive del
vacuolo centrale. Nell'apice dall'esterno verso l'interno si individua:
il protoderma, primo strato di cellule che darà origine al sistema tegumentale;
il procambio, formato da numerosi strati di cellule allungate, in posizione centrale, che
daranno origine al sistema conduttore;
il meristema fondamentale, costituito da alcuni strati di cellule di dimensioni più ampie, da
cui si origineranno i sistemi dei tessuti fondamentali.
Si dicono tessuti primari quelli derivati dal meristema primario.
MERISTEMI SECONDARI O LATERALI
Iniziano la loro attività dipleurica (verso l'interno e verso l'esterno) dopo che i tessuti primari sono
maturati. Si distinguono in:
cambio cribro-vascolare o cambio: è costituito da cellule allungate con pareti sottili, ma
più spesse di quelle del meristema apicale, e si evolve originando xilema verso l'interno e
floema verso l'esterno;
cambio subero fellodermico o Fellogeno: produce verso l'esterno sughero e verso l'interno
un tessuto parenchimatico chiamato felloderma.
Sono tessuti secondari quelli che derivano da meristemi secondari.
TESSUTI DEFINITIVI
I tessuti definitivi sono formati da cellule che hanno perduto la capacità di dividersi e si sono
specializzate. Sono presenti in tutti gli organi dove si formano tre sistemi unitari che mostrano la
continuità del corpo vegetale.
TESSUTI FONDAMENTALI
Il sistema dei tessuti fondamentali è costituito da tessuti semplici, ovvero tessuti formati da cellule
più o meno simili come caratteristiche strutturali, come:
Parenchima: si trova un po' in tutti gli organi, da solo o combinato con altri tessuti più
specializzati, ed è considerato un tessuto base sia da un punto di vista morfologico che
fisiologico. È costituito da cellule sempre vive anche a maturità, con nucleo, vacuolo e
parete cellulare formata da cellulosa e pectine è sottile. Il contatto tra una cellula e l'altra
avviene solo in poche zone e quindi si formano numerosi spazi intercellulari.
I tessuti parenchimatici si distinguono in: parenchimi clorofilliani, aeriferi, acquiferi, di
riserva.
Parenchima clorofilliano di assimilazione: presente nella foglia, nei fusti verdi e
nello strato sottocorticale del tronco e dei rami, possiede cellule ricche di cloroplasti
ed è perciò il tessuto in cui si svolge la fotosintesi. Si distingue in:
parenchima clorofilliano a palizzata con cellule allungate, a perfetto contatto fra
loro, disposte una sull'altra in modo ordinato;
parenchima clorofilliano lacunoso con cellule di forma varia disposte a formare
una rete con spazi intercellulari molto evidenti fino a raggiungere il 70% del
volume totale.
Parenchima aerifero: si trova nelle piante che hanno uno scarso ricambio d'aria come
ad esempio le palustri. Al di sotto dell'epidermide una rete di cellule con maglie
molto ampie, entro cui circola l'aria, favorisce così il trasporto di ossigeno in
profondità.
Parenchima acquifero: è situato in profondità nel fusto e nelle foglie delle piante che
vivono in ambienti aridi e hanno quindi bisogno di trattenere acqua da utilizzare nei
periodi di siccità. Ha cellule provviste di un grosso vacuolo ricco di acqua.
Parenchima di riserva: ne sono dotati semi, fusti e radici, ovvero gli organi della
pianta che accumulano sostanze di riserva. Possiede cellule grandi, con parete sottile,
e vacuoli ricchi di oli o amidi.
Parenchima di trasporto: formato da cellule transfer, associate ai tessuti conduttori;
queste hanno pareti con introflessioni e si trovano nei cotiledoni, nelle foglie di
piante erbacee, nelle strutture riproduttive e in quelle ghiandolari, svolgendo un ruolo
importante nel trasferimento dei soluti a piccole distanze.
Collenchima: si trova soprattutto nei fusti giovani e nelle foglie. È formato da cellule vive,
con citoplasma ricco di cloroplasti, un grosso vacuolo centrale e pareti cellulari ispessite di
cellulosa.
Sclerenchima: si trova nelle zone mature della pianta che hanno terminato l'accrescimento.
Ha cellule morte poiché gli strati di lignina e cellulosa abbinati, occupano tutto lo spazio
citoplasmatico. L'ispessimento può avvenire in diverse direzioni: se viene in tutte e tre le
dimensioni dello spazio, si hanno le sclereidi, se lo sviluppo maggiore avviene in un'unica
direzione, si formano le fibre che, essendo allungate, possono venire utilizzate nell'industria
tessile. Alcune cellule sclerose di forma varia, dette idioblasti, servono a impedire lo
schiacciamento del parenchima aerifero e danno forma agli spazi intercellulari.
Sia il collenchima che lo sclerenchima hanno la specifica funzione di sostenere la pianta e
di proteggerne gli organi; sono pertanto considerati tessuti meccanici. Si formano per
progressivo ispessimento delle pareti cellulari del parenchima iniziale; entrambi sono
caratterizzati da cellule isodiametriche.
TESSUTI CONDUTTORI
Il sistema dei tessuti conduttori o vascolari è formato da tessuti complessi, ovvero tessuti formati
da cellule che, pur essendo differenti, compiono la stessa funzione.
I tessuti conduttori derivano dal procambio nelle strutture primarie e dal cambio nelle strutture
secondarie e consentono il trasporto delle soluzioni provenienti dall'ambiente esterno e di quelle
elaborate dalla pianta.
Dalle radici alle foglie l'acqua scorre nei vasi legnosi, mentre per mezzo dei vasi cribrosi avviene la
distribuzione delle sostanze elaborate a tutti gli organi della pianta.
Vasi legnosi (o xilema): sono costituiti da cellule morte, prive di citoplasma, allungate, di
forma cilindrica e sovrapposte. Si distinguono in trachee, se il setto mediano divisorio fra
cellula e cellula si dissolve totalmente, e tracheidi, se fra cellula e cellula rimangono degli
ispessimenti di lignina. I vasi legnosi nelle gimnosperme sono tutte tracheidi (legno
omoxilo). Le tracheidi estive assumono anche funzione di sostegno e prendono il nome di
fibro-tracheidi. Nelle angiosperme sono presenti sia trachee sia tracheidi (legno eteroxilo).
Per impedire eventuale risalita dell'acqua durante il riposo vegetativo, alcune estroflessioni
delle cellule parenchimatiche vicino al vaso, i tilli, ne obliterano il lume.
Vasi cribrosi (o floema): sono formati da cellule vive con setto divisorio unteggiato da tanti
forellini, i cribri, che quando la pianta entra in riposo vegetativo, vengono chiusi da una
sostanza, il callosio, secreta dalle cellule stesse. Nella primavera successiva, un enzima
presente nel citoplasma, la scioglie.
FASCI CRIBROVASCOLARI
Nella struttura primaria l'insieme dei tessuti conduttori e delle fibre costituisce i fasci
cribrovascolari che, a seconda della disposizione dello xilema e del floema possono essere
ulteriormente distinti in:
Fasci raggiati: xilema e floema alternati a raggiera;
Fasci concentrici: xilema e floema formano due anelli concentrici, in genere perifloematici
(floema all'esterno), perixilematici (xilema all'esterno, per alcune monocotiledoni e in alcuni
fusti sotterranei e fusti aerei, oppure nei tessuti differenziati);
Fasci collaterali: i fasci xilematici si contrappongono a quelli floematici; si dicono chiusi se
i due tessuti sono a contatto (monocotiledoni), aperti se sono inframezzati dal cambio
(dicotiledoni e gimnosperme); in genere questi tipi di fasci sono nelle zone di
differenziazione;
Fasci bicollaterali: sempre aperti e xilema al centro di due cordoni di floema; si trovano
solo in alcune dicotiledoni (Cucurbitacee, Solanacee, Convolvolacee).
TESSUTI TEGUMENTALI
Il sistema dei tessuti tegumentali è formato da tessuti complessi. I tessuti tegumentali costituiscono
gli strati cellulari più esterni della pianta, hanno il compito di proteggerne le porzioni interne e sono
esposti ad urti ambientali e a variazioni. Questi strati di cellule sono anche quelli che, più degli altri,
mostrano specializzazioni ed adattamenti particolari.
I tessuti tegumentali sono costituiti dall’epidermide nella struttura primaria, dal periderma e dai
tessuti secretori nella struttura secondaria.
Epidermide: è un tessuto di protezione formato da un unico strato pavimentoso, provvisto
di uno strato impermeabile, la cuticola, che impedisce la dispersione dell'acqua.
L'epidermide ha origine dal protoderma e riveste foglie, frutti parti fiorali e organi giovani
della pianta. Questo tipo di tessuto prende il nome rizoderma se riguardante la radice.
Un esempio di specializzazione si ha nelle piante definite "d'appartamento", dove questo
strato è pluristratificato e svolge anche la funzione di riserva acquifera; nelle piante che
vivono in ambienti aridi, e devono quindi ridurre il più possibile la dispersione, si hanno
invece ispessimenti di cutina e secrezioni di cera.
Le cellule dell'epidermide, sempre vive, hanno forma variabile, sono strettamente unite le
une alle altre, contengono poco citoplasma e ampi vacuoli. Sono tutte trasparenti perché
prive di cloroplasti, tranne gli stomi, cellule modificate allo scopo di permettere gli scambi
gassosi.
Nelle piante che vivono in ambienti umidi talora, accanto agli stomi, sono presenti gli stomi
acquiferi o idatodi, aperture rigide poste sul bordo della foglia, attraverso le quali fuoriesce
acqua allo stato liquido. Al contrario, nelle piante che vivono in ambienti aridi, gli stomi
sono affossati in cripte, talvolta coperte da peli.
Estroflessioni unicellulari o pluricellulari delle cellule, i peli o tricomi, limitano la
traspirazione, ma svolgono anche azione protettiva (come nel caso dell'ortica) o secernono
oli essenziali.
Nei petali l'epidermide è costituita dalle papille coniche, cellule leggermente allungate e
rialzate, con citoplasma ricco di cromoplasti che conferiscono il caratteristico aspetto
vellutato. I peli radicali, che hanno il compito di assorbire la soluzione nutritizia del terreno,
sono estroflessioni, prive di cutina, delle cellule dell'epidermide dell'apice della radice.
Periderma: si trova nelle strutture secondarie della pianta e riveste fusti e radici. È formato
da tre tessuti: un parenchima, il felloderma; un meristema, il fellogeno; uno strato di cellule
morte, il sughero. Gli scambi gassosi con l'ambiente esterno avvengono attraverso le
lenticelle, aperture rigide.
Tessuti secretori: sono i tessuti capaci di elaborare sostanze di grande importanza biologica
ed ecologica. Si dividono in tessuti secretori esterni ed interni.
Tessuti secretori esterni: nel citoplasma delle cellule dell'epidermide vengono secrete
le essenze. Esse diffondono attraversi la parete di cellulosa, provocando il distacco e
la distensione della cuticola che, toccata appena o dilatata dal calore del Sole, si
rompe e permette alla sostanza volatile di diffondere nell'aria.
Cellule secretrici possono trovarsi anche alla sommità dei tricomi. Allo stesso modo,
sostanze viscose contenenti enzimi proteolitici sono secrete dalle cellule delle foglie
di piante carnivore, che vivono in terreni poveri di azoto.
Cellule secretrici esistono nei frutti, in alcune foglie e soprattutto nella corteccia
degli alberi. Esse producono tannini, contenuti nei vacuoli e nelle pareti cellulari,
che hanno funzione antisettica.
Tessuti secretori interni: sono racchiusi nelle zone più profonde e interne degli
organi. A volte sono formati da masse compatte di cellule, più spesso versano i
prodotti elaborati in cavità interne. Le cellule si allontanano le une dalle altre dando
origine a un canale.
I canali resiniferi delle foglie e dei fusti delle Conifere sono capaci di secernere
resine con funzione di protezione e difesa, circondati all'esterno da un tessuto
sclerenchimatico che conferisce rigidità e li mantiene sempre aperti.
Altre cavità di diversa origine sono tipiche del periderma degli agrumi, derivano dal
dissolvimento delle cellule secretrici dopo che hanno elaborato le loro essenze.
Alcuni tessuti secretori presentano strutture particolari come i tubuli laticiferi,
cellule allungate e ramificate chi percorrono tutto il vegetale e producono il lattice,
una soluzione bianca e viscosa contenente varie sostanze tra cui alcuni alcaloidi.
APPARATO RADICALE: MORFOLOGIA E ANATOMIA
La prima radice è detta radice primaria, può persistere tutta la vita della pianta o degenerare. Il
sistema radicale si può sviluppare in due modi: a fittone o fascicolato.
Nel fittone c'è una radice principale conica, con radici laterali, esili e progressive. Nel fascicolato
vi sono molte radici ramificate.
A volte le radici non si originano dalla radice primaria, ma dal fusto o dalle foglie: in questo caso si
parla di radici avventizie. Molte piante che in condizioni normali non utilizzano questa possibilità,
in certe situazioni sono in grado di emetterle; questa proprietà viene sfruttata per la propagazione
agamica per talea e sistemi simili.
ZONE DELLA RADICE
Nella radice delle piante possono essere individuate alcune zone:
Zona apicale: è formata da una cuffia o caliptra e da una zona di accrescimento per
divisione;
Cuffia: è lo strato più esterno formato da cellule epidermiche a forma di guaina, che
ha il compito di proteggere l'apice, rispondere allo stimolo della gravita
(geotropismo), favorire la penetrazione della radice fra le particelle del terreno. A
questo scopo nelle vescicole dell'Apparato del Golgi viene secreta una sostanza che
agisce da lubrificante. Man mano che la radice cresce, la cuffia viene Spinta in avanti
e le cellule che a contato con il terreno si sfaldano sono sostituite da altre prodotte da
un tessuto meristematico chiamato caliptrogeno.
Zona di accrescimento per divisione: formata da tessuto meristematico che
dividendosi per mitosi presiede all'allungamento dell'organo. Solo le cellule terminali
non possiedono attività mitotica, ma costituiscono la zona quiescente, sede della
elaborazione della maggior parte degli ormoni.
Zona liscia o di accrescimento per distensione: si tratta della porzione immediatamente
sopra l'apice, ha lunghezza da 1 a10 mm e termina dove i tessuti sono completamente te
differenziati. Poche sono le mitosi e l'accrescimento cellulare avviene in senso longitudinale.
Zona di struttura primaria: è posta successivamente alla zona di accrescimento per
distensione, è costituita da cellule mature in grado di espletare la loro funzione. Osservando
in senso longitudinale, si riconoscono due zone distinte, la zona pilifera e la zona suberosa.
Osservando in senso trasversale, si distinguono vari strati:
Zona pilifera:
rizoderma: si origina dal protoderma e ha cellule in stretto contatto. Alcune di esse
si allungano a formare peli unicellulari adatti ad assorbire acqua e sali minerali.
corteccia o cilindro corticale: si origina dal meristema fondamentale. Le cellule,
prive di cloroplasti, sono connesse da plasmodesmi e hanno funzione di riserva di
amido e altre sostanze.
endoderma o cilindro centrale: è lo strato più interno della corteccia e presenta
nella parete primaria un ispessimento nastriforme noto come banda del Caspary,
impermeabile all'acqua perché impregnato di suberina o lignina. Nelle
dicotiledoni la banda del Caspary interessa solo le pareti radiali e trasversali come
un tubo. Nelle monocotiledoni la banda del Caspary ha un ispessimento anche
nella parete tangenziale interna e assume la forma di una U o di un tubo a fondo
chiuso. Solo alcune cellule dell'endoderma, dette punti di permeazione, non
ispessiscono la parete tangenziale interna.
periciclo: delimita il cilindro centrale; da questo derivano il cambio cribro
vascolare e le radici laterali.
cilindro vascolare: contiene i tessuti conduttori legnosi (xilema) e cribrosi
(floema) disposti fianco a fianco su raggi distinti, detti raggi midollari, che
partono dal centro del midollo centrale. Esso è sempre presente nelle
monocotiledoni, mentre è spesso assente nelle dicotiledoni.
Zona suberosa: presenta un tessuto epidermico resistente detto esoderma, con cellule
allungate impregnate di suberina. I punti di permeabilità ai liquidi sono costituiti da
alcune cellule vive e ricche di plasma. In questa zona hanno origine le radici
secondarie o laterali, secondo il seguente processo.
Le cellule del periciclo cominciano a dividersi, formando l'apice della nuova
radichetta;
Contemporaneamente anche l'endoderma comincia ad accrescere con divisioni
anticlinali e origina uno strato detto strato digerente perché secerne un enzima, la
pectidasi, che idrolizza la pectina e apre la strada alla radichetta fra le cellule della
corteccia;
Inizialmente non vi è connessione fra i tessuti conduttori della radice madre e
quelli della radichetta laterale, il contatto avviene in un secondo tempo ad opera di
cellule derivate dal periciclo;
Una volta raggiunto il terreno intorno all'apice, si ha la formazione della cuffia e
la radichetta fuoriesce.
Zona di struttura secondaria: questa zona non si trova nelle radici delle monocotiledoni, il
cui accrescimento diametrale avviene ad opera delle cellule del periciclo. Nelle
gimnosperme e nelle dicotiledoni, invece, l'accrescimento in spessore è operato da un
meristema secondario, il cambio.
Il cambio si origina dalle cellule parenchimatiche dei raggi midollari e si inserisce fra la
parte esterna del legno e la parte interna del libro, assumendo un andamento sinusoidale.
Poiché svolge attività dipleurica, producendo verso l'interno legno secondario e verso
l'esterno libro secondario, si originano fasci collaterali aperti.
Quando il cambio raggiunge il periciclo, anche questo acquista attività meristematica e il
tessuto meristematico diviene, da sinusoidale, circolare.
SISTEMA DEL GERMOGLIO
Il germoglio comprende fusto e foglie e si origina dalla plumula embrionale, formata da un asse
(epicotìle), da una o più foglioline immature (primordi fogliari) e da un meristema apicale.
La gemma è un germoglio non ancora sviluppato presente in tutti i tipi di fusti. Essa continua
l'accrescimento in lunghezza del fusto (gemma terminale o apicale) o si evolve all'ascella della
foglia in un primordio di ramo (gemma laterale o ascellare).
Generalmente la gemma, formata da tessuto meristematico circondato e protetto da un cappuccio
embricato di foglioline appressate a causa di un diseguale accrescimento delle due facce, si
differenzia durante l'estate. In inverno viene ricoperta da squame protettive di consistenza coriacea,
dette perule, ed entra in un periodo di quiescenza (gemma dormiente o ibernante). In base alla loro
diversa attività, possono essere distinte da esse le gemme avventizie. In primavera, le squame
cadono lasciando cicatrici nel punto cui si staccano e comincia la crescita della pianta. Spesso le
cellule del meristema apicale secernono sostanze inibitrici che bloccano lo sviluppo delle gemme
laterali (dominanza apicale), il cui sviluppo avviene solo quando, con l'accrescimento della pianta,
aumenta la loro distanza dall'apice.
La gemma apicale rimane costantemente attiva nel fusto monopodiale cosicché l'asse principale si
allunga per tutta la vita, mentre si arresta dopo un certo tempo in quello simpodiale, per cui
l'allungamento del fusto prosegue attraverso le gemme laterali. Nella ramificazione dicotomica sono
le gemme laterali che si sviluppano con conseguente biforcazione dell'asse principale e dei rami.
Le gemme possono contenere solo il primordio di un asse vegetativo (gemme a legno), solo gli
abbozzi di un fiore (gemme a fiore) o un primordio di un asse vegetativo con un fiore (gemme
miste).
IL FUSTO: MORFOLOGIA E ANATOMIA
Il fusto di una pianta ha la funzione si sostenere foglie, fiori, frutti, di orientarli nello spazio e di
trasportare acqua e sostanze nutritive; in certi casi svolge anche la fotosintesi e può avere funzioni
di riserva.
La disposizione del fusto e dei rami condizionano il portamento della pianta; il fusto si collega alle
radici in una zona detta colletto e si collega alle foglie in zone ingrossate dette nodi; i segmenti tra i
nodi sono detti internodi.
Si trovano fusti erbacei e legnosi. Si hanno anche fusti metamorfosati con funzioni di riserva,
nutrimento, difesa, trasporto di ossigeno e fotosintesi (esempi: rizoma – gamigna; tubero – patata;
bulbo – cipolla; bulbo-tubero – tulipano; stolone - fragola, cladodi; fogliforme – pungitopo; fusti
sgerici colonnari – cactus; viticci - vite, spine - biancospino).
A partire dall'apice del germoglio, è possibile distinguere varie zone del fusto:
Zona embrionale o di accrescimento: è l'apice del fusto, misura poche centinaia di micron
ed ha la caratteristica di essere in attiva divisione mitotica.
La zona apicale è costituita dal cono vegetativo o dall’apice del germoglio; le
protuberanze che in successione acropeta compaiono sui fianchi del cono vegetativo sono
dette bozze fogliari; altre protuberanze sono chiamate primordi dei rami. A tutto questo
complesso si da il nome di gemma.
La zona distale dell'apice viene detta promeristema da cui derivano i tre meristemi
apicali del fusto: protoderma, procambio e meristema fondamentale.
Zona di differenziazione: questa zona segue la zona embrionale; le cellule sospendono
l'attività meristematica e si differenziano nei tessuti adulti. Dal protoderma si differenzia
l'epidermide, dal procambio il sistema vascolare (xilema primario e floema primario), dal
meristema fondamentale i tessuti parenchimatici. La differenziazione è un processo lento e
non sincrono.
L'evento comune che caratterizza le cellule di questa zona è l'accrescimento per distensione,
attraverso il quale gli internodi si allungano e le foglie si distanziano nel fusto. Questo
accrescimento è regolato da ormoni, tra i quali il più importante è l'auxina, la cui azione è
profondamente influenzata dalla luce.
Zona di struttura primaria: si definisce così l'insieme dei tessuti adulti primari originatisi
dai meristemi apicali. Nelle dicotiledoni arboree e nelle gimnosperme questa zona si estende
per alcuni millimetri o pochi centimetri; nelle monocotiledoni è in genere la struttura
definitiva; per le dicotiledoni erbacee è molto estesa e si formano poche zone in struttura
secondaria.
Come nella radice si individuano 3 regioni: epidermide (tessuto tegumentale), corteccia
(tessuti parenchimatici e meccanici), cilindro centrale (tessuto vascolare e parenchimatico).
Epidermide: formato da unico strato di cellule con compito di protezione (tessuto
tegumentale) e di contrastare le perdite di acqua grazie ad un rivestimento ceroso che
impedisce la traspirazione (cuticola),
Cilindro corticale o corteccia: costituito da tessuto parenchimatico pluristratificato, i
cui strati esterni sono di tipo clorofilliano, quelli più interni sono ricchi di leucoplasti.
Lo strato finale, detto guaina amilifera, è ancora più ricco di amido. Sotto
l'epidermide, in posizione superficiale, si trovano anche cordoni di collenchima
(tessuti meccanici).
Cilindro centrale o stele: è la parte più interna, che deriva dal meristema
fondamentale (parenchima midollare) e in parte dal procambio (tessuto vascolare).
Lo stato periferico che confina con la corteccia si chiama periciclo.
I fasci cribro-vascolari delle dicotiledoni e delle gimnosperme sono, in genere, di
tipo collaterale aperto e risultano distribuiti in una cerchia regolare. Tali fasci sono
immersi in un tessuto di natura parenchimatica, dove si distingue una parte centrale
chiamata midollo e porzioni interposta tra i fasci conduttori chiamati raggi
midollari. Questo tipo di struttura si dice eustelica. Spesso ai fasci conduttori sono
associati tessuti meccanici (sclerenchima), più spesso sul lato esterno floematico.
I fasci cribro-vascolari delle monocotiledoni sono, in genere, sparsi in tutto il suo
spessore; tali fasci sono di tipo collaterale chiuso senza cellule procambiali tra il
metaxilema (interno) e metafloema (esterno). In genere i fasci sono di alcune decine
o, nel caso delle palme, alcune centinaia.
Zone di struttura secondaria: nel fusto delle dicotiledoni e delle gimnosperme compaiono
abbastanza presto due meristemi laterali secondari: il cambio cribro-vascolare e il cambio
subero-fellodermico. Questi meristemi producono tessuti secondari che vanno ad alterare
profondamente la struttura primaria. In seguito alla formazione di tessuti secondari il fusto si
accresce in spessore. Il cambio cribro-vascolare determina l'accrescimento in spessore del
cilindro centrale, quello subero-fellodermico quello della corteccia. In genere il contributo
del cambio subero-fellodermico all'aumento in spessore del fusto è limitato, solo in pochi
casi è rilevante (quercia da sughero).
Cambio cribro-vascolare: si origina dalle cellule procambiali dei fasci collaterali
aperti della struttura primaria e si estende alle cellule parenchimatiche interposte tra
i fasci che riacquistano caratteri meristematici. Il cambio ha attività dipleurica (sui
due lati): le cellule del lato esterno differenziandosi formeranno il floema secondario,
quelle del lato interno lo xilema secondario. Il cambio delle piante di zone temperate
è attivo dall'inizio della primavera alla fine dell'estate/autunno. L'attività è sospesa
durante il periodo invernale. I meccanismi che sovrintendono a questa sospensione
sono molto complessi e non ancora del tutto chiari. Il cambio continua ad essere
attivo per tutta la vita della pianta e anno genera xilema secondario e floema
secondario: ne consegue che anche la cerchia cambiale deve aumentare di
circonferenza. Anche il floema secondario per essere posto all'esterno del cambio è
soggetto a sollecitazioni per allargarsi e forma quindi un parenchima detto di
dilatazione.
Si formano dalle cellule del cambio anche i raggi parenchimatici o raggi midollari
che assicurano il trasporto radiale.
Xilema secondario: nel fusto delle dicotiledoni svolge funzioni di conduzione (vasi),
riserva (parenchima) e sostegno (fibrotracheidi e fibre libriformi). Il legno che si
viene a formare da questi elementi eterogenei viene detto legno eteroxilo. In una
sezione del fusto di dicotiledone con qualche anno di età si rilevano le cerchie
annuali (zone concentriche di diametro progressivamente più grande), dove si può
distinguere il legno primaverile (con grandi vasi, molto parenchima e poche fibre) e
quello estivo (pochi vasi e piccoli, poco parenchima molte fibre).
Nel fusto delle gimnosperme ha una struttura più semplice ed omogenea ed è
costituito da un solo tipo di elementi, le tracheidi (nel legno estivo sono dette
fibrotracheidi, poiché sono più efficienti a svolgere la funzione meccanica). Queste
sono quasi tutte uguali e sono allineate secondo file radiali. Questa regolare
disposizione determina la denominazione di legno omoxilo. Nelle gimnosperme
spesso si trovano i canali resiniferi.
Floema secondario o libro: nelle dicotiledoni, il sistema assiale è costituito da
elementi di conduzione (tubi cribrosi), da elementi meccanici (fibre) e da cellule
parenchimatiche; il sistema radiale è formato da cellule parenchimatiche che
formano i raggi parenchimatici: questi sono una diretta continuazione di quelli del
legno ed in genere aumentano di spessore nel libro poiché formano anche il
cosiddetto parenchima di dilatazione. Questo parenchima è necessario per impedire
che il libro subisca lacerazioni dal successivo accrescimento annuale. In genere i tubi
cribrosi funzionano per una sola stagione, raramente per due (vite). La quantità di
libro che si accumula dipende dalla posizione dove si differenzia il fellogeno e se si
differenzia tutti gli anni oppure ad intervalli più lunghi.
Nelle gimnosperme si hanno cellule cribrose, cellule parenchimatiche e fibre
(mancanti nelle pinacee). Ci sono anche i canali resiniferi che proseguono dallo
xilema.
Cambio subero-fellodermico o fellogeno: è un tessuto meristematico che si
differenzia da cellule di tipo epidermico, parenchiamtiche, collenchimatiche e
floematiche. Anch'esso ha attività dipleurica: verso l'esterno produce il sughero e
verso l'interno il felloderma, con attività più intensa comunque verso l'esterno. Il
fellogeno è attivo solo per un anno: ogni anno si differenzia un nuovo fellogeno in
posizione sempre più interna rispetto a quello precedente. Dato che il fellogeno si
differenzia ogni anno in uno strato più interno, tutti gli strati esterni precedenti
rimangono isolati dalla pianta (esterni al sughero). Questi sono tessuti morti e vanno
a costituire la scorza o ritidoma. Continuando, anno dopo anno, iniziano a
differenziarsi per il fellogeno cellule che erano del libro e quindi via via gli strati del
libro si troveranno nel ritidoma e si perderanno: ecco perché l'accrescimento in
spessore è dovuto solo allo xilema, ed il libro è presente solo in piccole quantità.
LA FOGLIA
La foglia è il laboratorio principale della pianta: essa svolge diverse funzioni tra cui, la più
importante, la fotosintesi clorofilliana. Inoltre, ha altre attività metaboliche e sintetizza, oltre agli
zuccheri, ormoni, vitamine, aminoacidi. È la sede principale della traspirazione.
Le foglie sono appendici laterali del fusto ed in genere si inseriscono a livello dei nodi, in modo da
poter ottimizzare la superficie di ciascuna foglia esposta alla luce. La distribuzione delle foglie sul
fusto è nominata tassia fogliare o fillotassi.
Nella foglia delle angiosperme, tipicamente, si distingue una lamina fogliare e un picciolo: questo
può anche mancare e allora le foglie si dicono sessili.
A volte la foglia è provvista della guaina che rappresenta una espansione della base del picciolo (in
genere è presente nelle monocotiledoni), alla base della foglia possono esserci delle appendici di
varia forma e grandezza dette stipole.
Le foglie si distinguono per la forma e la consistenza della lamina, per il margine e per la
disposizione delle nervature. Vi sono molte foglie modificate nella forma e nella funzione: gli
esempi più rappresentativi sono gli embriofilli ( foglie cotiledonari), gli ipsofilli (brattee fiorali con
funzione vessillare per fiori poco appariscenti), gli antofilli (sepali e petali), gli sporofilli (stami e
carpelli).
Struttura della lamina fogliare
Anche la foglia è costituita da un sistema tegumentale (epidermide), un sistema fondamentale
(parenchimi) un sistema vascolare (fasci cribro-vascolari).
Una tipica foglia di dicotiledone, con orientamento plagiotropo (foglia dorsoventrale), presenta due
facce distinte, una superiore colpita dai raggi del sole ed una inferiore rivolta verso il terreno. Il
lembo fogliare su entrambe le facce è rivestito da epidermide. I tessuti compresi tra l'epidermide
superiore e inferiore costituiscono il mesofillo fogliare, composto da due porzioni il tessuto a
palizzata (vicino alla epidermide superiore) e il tessuto lacunoso (vicino a quella inferiore).
Epidermide: ha funzione di protezione. La parete cellulare delle cellule epidermiche non è
omogenea su tutta la superficie: la parete rivolta verso l'esterno è ispessita ed impregnata di
cutina (acidi grassi idrossilati e polimerizati, quindi idrofoba), a volte può essere ricoperta
da uno strato più o meno spesso di cutina a cui viene dato il nome di cuticola. La cuticola è
più o meno sviluppata a seconda della necessità della pianta di limitare o meno le perdite di
acqua. Al di sopra della cuticola, più spesso sulla faccia superiore, si possono trovare le
cere. I rivestimenti cerosi danno un aspetto vellutato (es. tulipani, iris). A volte la parete può
essere impregnata di silice (culmo delle graminacee con margini taglienti). Le cellule
epidermiche, ad eccezione delle cellule di guardia degli apparati stomatici, non sono dotate
di cloroplasti ben differenziati, ma rudimentali. Presenti spesso nella pagina inferiore i peli o
tricomi unicellulari o pluricellulari: in genere sono morti e ripieni di aria. Una funzione dei
peli è quella di mantenere uno strato di aria circostante saturo di vapore e determina quindi
un rallentamento del processo di evaporazione all'interno della foglia.
Nelle foglie dorsoventrali, sulla pagina inferiore sono presenti gli stomi, in numero variabile
da poche decine a centinaia per mm2, mediamente tra 200 e 400. Gli stomi (dal greco stoma
= bocca) permettono gli scambi gassosi e ne permettono la regolazione.
Volendo tracciare lo schema di costituzione di uno stoma, osservando frontalmente possiamo
riconoscere due cellule reniformi dette cellule di guardia, affacciate l'una contro l'altra a
delimitare una apertura detta rima stomatica, di lunghezza variabile da 5 a 40 micrometri.
Le cellule di guardia sono dotate di cloroplasti. Accanto alle cellule di guardia vi sono
spesso altre due cellule dette cellule annesse. La rima stomatica è in comunicazione con un
ampio spazio intercellulare sottostante detto camera sottostomatica. La parete delle cellule
di guardia è più sottile nella parte che guarda le cellule annesse e più spessa nella parte verso
la rima stomatica. Quando nelle cellule di guardia vi sono variazioni del turgore, queste
subiscono variazioni di volume e di forma dovute anche al diverso inspessimento e quindi
diversa estensibilità della parete. Quindi il risultato è un restringimento o allargamento della
rima stomatica. Quando sono turgide le cellule di guardia tendono a incurvarsi e quindi la
rima è più aperta, quando sono meno turgide si afflosciano e la rima arriva anche a
chiudersi. In genere gli stomi sono aperti di giorno e chiusi di notte. Il meccanismo di
apertura e chiusura degli stomi non è ancora del tutto chiaro, potrebbe dipendere dalla luce,
dalla concentrazione di zuccheri, dal tenore di anidride carbonica, da variazioni del pH,
oppure dalla maggiore o minore concentrazione di ioni K+; probabilmente esiste un
meccanismo di regolazione complesso a feed back.
Mesofillo fogliare: comprende i tessuti compresi tra l'epidermide superiore e quella inferiore
ed è formato da tessuti parenchimatici dove scorrono anche i vasi conduttori. Nelle foglie
bifacciali è ordinariamente suddiviso in tessuto a palizzata e lacunoso. Il tessuto a palizzata
è costituito da cellule allungate a forma cilindroide orientate perpendicolarmente alla
superficie della foglia, disposte in modo ordinato l'una vicina all'altra. Sono disposte in un
numero di strati variabile da 1 a 3: il numero di strati è influenzato dalla luce solare. Le
cellule del tessuto a palizzata sono dotate di numerosi cloroplasti ed è un tessuto
particolarmente atto allo svolgimento della fotosintesi. In genere si trova subito sotto
l'epidermide superiore.
Al di sotto del tessuto a palizzata, ed esteso fino alla epidermide inferiore, si trova il tessuto
lacunoso, anch'esso di tipo parenchimatico, con cellule di forma irregolare inframmezzate
da ampi spazi intercellulari. Le cellule del tessuto lacunoso sono dotate di un minor numero
di cloroplasti e con un sistema lamellare meno sviluppato. La funzione è quella di assicurare
un grande ricambio di aria necessario per un efficiente svolgimento della fotosintesi.
Sistema vascolare: Le nervature che si notano nelle foglie altro non sono che il
prolungamento dei fasci vascolari del fusto. Le nervature possono essere retinervie con
nervature principali e secondarie e numerose anastomosi, invece nelle parallelinervie ci
sono rare anastomosi con scarsa capacità di conduzione. In genere sono fasci cribrovascolari chiusi, con xilema verso l'alto e floema verso il basso.
Sistema meccanico: per le foglie più grandi è necessaria la presenza di tessuti meccanici di
tipo sclerenchimatico che sono disposti attorno ai fasci cribro-vascolari, molto abbondanti
per es. nell'agave e nella sanseveria. Spesso sono presenti anche i tessuti collenchimatici,
sempre in prossimità delle nervature e del picciolo.
ASSORBIMENTO E TRASPORTO DELLA LINFA
ASSORBIMENTO
Le piante assorbono anidride carbonica dall'aria e per il 96% sono costituite da composti
sintetizzati a partire dal carbonio della CO2; inoltre le piante assorbono dal suolo acqua, ioni
inorganici e una certa quantità di ossigeno.
Le piante con la fotosintesi producono zuccheri che poi riutilizzano per fini energetici e strutturali,
combinano e trasformano questi con gli ioni inorganici. Attraverso la respirazione cellulare
demoliscono gli zuccheri per produrre energia, ricaricando ADP.
La radice è l'organo preposto all'assorbimento di acqua e sali minerali dal terreno. L'acqua e i soluti
devono attraversare l'epidermide e la corteccia della radice per arrivare nei vasi conduttori
xilematici: poiché le membrane cellulari sono selettivamente permeabili, solo determinati soluti
sono in grado di entrare nello xilema.
La penetrazione delle sostanze nella radice può seguire una via intracellulare, in cui acqua e soluti
attraversano la parete cellulare e la membrana delle cellule dell'epidermide a livello di un pelo
radicale. Poiché le cellule sono collegate grazie ai plasmodesmi, la soluzione acquosa può, passando
di cellula in cellula, arrivare fino all'endoderma e da qui essere ammessa allo xilema. Esiste anche
una via extracellulare, cioè la soluzione si insinua tra le pareti cellulari senza penetrare nelle cellule,
e non trova alcun impedimento fino all'endoderma. Qui trova la banda impermeabile del Caspary
che impedisce l'ulteriore avanzamento. Gli ioni sono quindi sottoposti alla selezione, da parte della
membrana, di una cellula endodermica per poter accedere allo xilema (diffusione, trasporto
facilitato, trasporto attivo).
TRASPORTO DI ACQUA E IONI
Conosciamo già che le cellule di trachee e tracheidi a maturità sono morte. Il trasporto avviene con
la combinazione due meccanismi:
Pressione radicale: gli ioni accumulati nello xilema richiamano per osmosi acqua a cui
quindi viene impressa una certa spinta verso l'alto. Questo meccanismo ha una efficacia non
irrilevante (fenomeno della guttazione e del pianto nella vite, betulla, acero; se si provoca
una ferita in primavera prima della schiusura delle gemme, il liquido emesso ricco di glucidi
[idrolisi sostanze di riserva] è pari a 1 litro/giorno per la vite fino a 5/6 litri giorno per
betulla). Via via che ci sono le foglie diminuisce l'importanza della pressione radicale.
Traspirazione: una molecola di acqua evapora attraverso uno stoma o attraverso la cuticola
(solo il 15% della traspirazione è cuticolare) perché la concentrazione idrica è superiore
all'interno che all'esterno della foglia. Le cellule a ridosso della cavità stomatica quindi
perdono acqua a causa dell'evaporazione e quindi diminuisce la loro pressione di turgore e si
instaura una tensione di assorbimento. Il fenomeno a catena si propaga alla cellula
confinante da cui viene richiamata acqua fino a giungere allo xilema. La continua rimozione
di acqua dai vasi per opera delle cellule traspiranti del mesofillo determina nei vasi una
pressione negativa inferiore a quella atmosferica. Quindi nella sequenza di trachee e
tracheidi sottostanti si stabilisce un gradiente di pressione che fa si che l'acqua si diriga nelle
zone a pressione più bassa, quelle terminali. Questo meccanismo si propaga dai vasi della
foglia a quelli del fusto fino alla radice. Suolo, pianta e atmosfera costituiscono quindi un
unico sistema nel quale l'acqua si muove nella direzione verso cui il potenziale dell'acqua è
più basso. La traspirazione funziona così bene anche grazie alle forze di coesione tra le
molecole d'acqua (ponti H) e per le forze di adesione tra acqua e cellulosa o lignina delle
pareti dei vasi (ponti H). Con questo modello per spiegare la risalita dell'acqua si afferma
anche che il trasporto dell'acqua contro la forza di gravità non costa energia alla cellula.
La funzione principale della traspirazione è quella di far entrare per gradiente di diffusione
l'anidride carbonica nel tessuto lacunoso e quindi gli stomi devono stare aperti il più a lungo
possibile affinché possa essere organicato il carbonio con la fotosintesi. Inoltre, la
traspirazione permette anche l'ingresso di ossigeno, regola la temperatura e fa salire i
minerali necessari.
TRASPORTO DELLA LINFA ELABORATA
È il floema che trasporta la linfa elaborata. I vasi cribrosi sono formati da catene di cellule che
hanno le pareti cellulari con fori nelle placche e quindi la soluzione si può muovere liberamente.
L'acqua va solo verso l'alto, la linfa elaborata va dall'alto verso il basso, ma anche in altre direzioni.
La linfa elaborata contiene principalmente saccarosio ma anche ormoni, aminoacidi e ioni.
Le zone dove avviene la produzione di zuccheri sono dette sorgenti. Il floema trasporta gli zuccheri
dalle parti verdi fino alle cellule non fotosintetiche. Si dicono pozzi le zone dove gli zuccheri
vengono immagazzinati o consumati. I tuberi, i bulbi all'inizio della stagione vegetativa si invertono
e da pozzi diventano sorgenti.
La linfa sembra essere trasportata senza spesa energetica attraverso il meccanismo dettato dal
modello detto flusso di pressione. La linfa scorre alla velocità di circa 1 m/h. Dalle cellule che
producono gli zuccheri questi passano ai tubi cribrosi per trasporto attivo (spesa energetica),
aumenta la concentrazione di zuccheri nel floema; questa alta concentrazione di soluti richiama per
osmosi acqua dai vasi xilematici e il flusso di acqua fa aumentare la pressione idrostatica nel tubo
cribroso (sorgente). Al pozzo i soluti e l'acqua escono dal tubo cribroso per gradiente di
concentrazione: prima escono gli zuccheri poi li segue l'acqua per osmosi. Nel tubo cribroso
diminuisce la pressione idrostatica. È l'aumento della pressione idrostatica alla sorgente e la
diminuzione nel pozzo che fa sì che l'acqua con i soluti scorra dalla sorgente al pozzo.
Successivamente dal pozzo l'acqua esce e ritorna allo xilema per essere trasportata alla sorgente.
GLI ORMONI VEGETALI
Gli ormoni sono sostanze chimiche che coordinano le attività degli organismi animali e vegetali. In
genere sono prodotti in determinate zone e vengono traslocati successivamente nei distretti dove è
necessario, detti organi o tessuti bersaglio. In genere sono attivi a concentrazioni molto basse.
Spesso l'azione non è isolata ma con associazione di più ormoni. Spesso inducono la crescita, la
distensione cellulare, e il differenziamento cellulare. Il loro meccanismo di azione produce
modificazioni dell’ espressione genica, oppure alterando la permeabilità delle membrane.
Gli ormoni vegetali identificati sono 5 classi di sostanze.
Auxina o acido indolacetico: prodotta nel meristema apicale, diffonde verso il basso e il
fusto viene stimolato a crescere per allungamento delle foglie. Questa azione è attiva solo in
un determinato intervallo di concentrazione: se le concentrazioni sono troppo elevate si
ottiene l'effetto opposto, probabilmente per la secrezione da parte del fusto di etilene che
contrasta l'azione dell'auxina. Se le concentrazioni sono molto basse si ripercuote su un
allungamento delle radici che invece viene inibito a concentrazioni dentro l'intervallo. Le
auxine stimolano anche lo sviluppo dei tessuti vascolari e la divisione delle cellule cambiali.
Le auxine hanno anche l'effetto di dominanza apicale.
Citochinine: sono varie sostanze, tra le quali la zeatina, il 6 furfuril-aminopurina e la
benziladenina, prodotte generalmente nelle radici, nei semi e nei frutti che promuovono la
divisione cellulare. Dalle radici si diffondono e possono promuovere la crescita di gemme
laterali, che invece sono inibite dall'effetto auxinico di dominanza apicale.
Considerando che in genere la massa radicale è pari a quella aerea le citochinine potrebbero
avere questo compito di far sviluppare armonicamente le due parti.
Gibberelline: sono composti terpenoidi costituiti da una struttura di base a 5 atomi di
carbonio, l'isoprene. Si conoscono più di 30 diversi tipi di gibberelline. Queste sono prodotte
dai meristemi apicali e sub apicali del fusto, dalle foglioline e dai giovani embrioni. La loro
azione è rivolta verso la distensione cellulare e la divisione cellulare e partecipa inoltre
anche al fenomeno della fioritura e della ripresa embrionale dopo la dormienza del seme.
Acido abscissico o dormina: inizialmente fu scoperto nel cotone nel corso di studi sulla
abscissione delle foglie e prese da li il nome; da altre ricerche fu trovata una sostanza che
induceva la dormienza delle gemme che ha determinato l'altro nome. È prodotto dalle foglie
dai fusti e frutti verdi. La sua azione è in genere quella di indurre dormienza in condizioni
non idonee per la vegetazione. Questa dormienza può verificarsi nelle gemme, ma anche nei
semi che necessitano magari di certe dosi di freddo per eliminare l'ABA in eccesso, oppure
per altri che necessitano di acqua sempre per eliminarlo e quindi trovare le giuste condizioni
per la germinazione. Ha anche azione antistress. Sempre in condizioni ambientali avverse, di
scarsità di acqua, fa chiudere gli stomi rallentando le perdite di acqua.
Etilene: è una molecola molto semplice ed è, unico caso, gassosa. L'etilene è prodotto in
quantità durante la maturazione dei frutti. Provoca, in qualche modo, la scissione enzimatica
delle pareti cellulari e la conversione dell'amido in disaccaridi e monosaccaridi. Nei giovani
frutti la respirazione cellulare è molto attiva; quando raggiungono le dimensioni finale la
respirazione diminuisce. In alcuni frutti, detti climaterici, dopo questa diminuzione si ha un
rialzo di respirazione detto picco climaterico, successivamente decresce ed il frutto entra
nella fase di senescenza. In molti frutti il picco climaterico coincide con le migliori qualità
eduli del frutto medesimo (pera); nella mela e banana precede leggermente l'optimum di
qualità. La fase climaterica, anche sul frutto staccato dalla pianta, coincide con le attività
idrolitiche enzimatiche precedentemente ricordate; il rialzo climaterico coincide con la
produzione di etilene. Il meccanismo di azione non è ancora del tutto chiaro.
Nelle piante caducifogli, all'approssimarsi della stagione meno favorevole, si ha il fenomeno
della abscissione delle foglie che coincide con vari mutamenti di colore delle medesime,
con pigmenti nuovi o con pigmenti che sono scoperti dopo la scomparsa della clorofilla.
Questi fenomeni intervengono quando vengono recepite variazioni di ore luce e temperature
in diminuzione. Questi stimoli provocano variazioni nella produzione di auxine e di etilene:
le auxine diminuiscono e l'etilene viene prodotte dalle cellule della zona di abscissione alla
base del picciolo. Quando l’etilene prevale sulle auxine si ha la caduta della foglia.
TROPISMI
I movimenti delle piante, poco visibili per la loro lentezza, sono detti tropismi. Un tropismo è una
crescita irreversibile che avviene come risposta a determinati stimoli.
Fototropismo: la luce provoca la migrazione della auxina verso il lato oscuro aumentando
così l'allungamento delle cellule di quel lato e facendo così incurvare l'altro lato verso la
luce.
Geotropismo: le piante distinguono alto e basso, così le radici si dirigono verso il basso e i
germogli verso l'alto, anche in caso che siano posti da noi in maniera opposta. Il
meccanismo d'azione non è chiaro: forse organuli in grado di percepire la gravità sono
presenti su granuli di amido.
Tigmotropismo: cambiamento di direzione della crescita come risposta al contatto. Ad
esempio tutti i tipi di viticci che prima crescono in modo diritto e verticale, quando trovano
un appiglio si avvoltolano.
OROLOGI BIOLOGICI
Le piante hanno una specie di orologio biologico, regolato sul giorno solare, che fa si che le foglie
e i fiori assumano determinate posizioni quando c'è la luce e quando non c'è ne assumano altre. Ma
se queste piante vengono tenute al buio mantengono certi comportamenti con ritmi giornalieri anche
se magari perdono un po' il tempo e il ritmo magari si allunga di alcune ore.
Oltre alle attività giornaliera c'è un orologio biologico che regola i cicli stagionali. Fioritura,
germinazione, inizio e fine quiescenza in genere si svolgono in determinati periodi dell'anno a
seconda delle specie. Lo stimolo più importante che riescono a percepire è la diversa durata del
giorno e della notte. Ad esempio, per la fioritura, si trovano piante che sono brevidiurne, ovvero che
fioriscono nelle giornate più corte, e longidiurne che al contrario fioriscono con luce crescente. La
fioritura avviene solo quando le ore di luce sono superiori o inferiori alla soglia specifica.
Sopratutto le brevidiurne percepiscono la notte: se infatti un lungo periodo di buio (14 ore che
dovrebbe far fiorire la pianta) viene interrotto da una luce di modesta entità, anche per pochi minuti,
viene inibita la fioritura. Invece il periodo di luce può essere interrotto da brevi periodi di luce senza
per questo inibire la fioritura. Ogni specie ha quindi un periodo critico di buio.
Il sistema con cui la pianta si accorge se c’è luce o buio sembra essere un pigmento detto fitocromo
che esiste in due forme che si trasformano l'una nell'altra, il Pr e il Pfr. Il primo è la forma che si
trova al buio: con la luce avviene la trasformazione in Pfr; quando ritorna il buio si ritrasforma in Pr.
Se il fitocromo Pfr è presente sempre quando c'è la luce, per indurre la fioritura deve interagire con
l'orologio biologico.
STANZE NUTRITIVE NECESSARIE ALLE PIANTE
L'anidride carbonica è prelevata dall'aria, l'acqua e tutto il resto deve essere prelevato
essenzialmente dal terreno. Con opportune coltivazioni di piante in presenza di sola acqua e alcuni
minerali noti si sono determinati i principali elementi necessari alla vita delle piante.
I minerali per convenzione vengono divisi in due grandi categorie: macrominerali, macroelementi o
macronutrienti e micro minerali, microelementi o micronutrienti. La suddivisione è fatta in base alla
quantità necessaria.
Macronutrienti: C, H, N, S, P e inoltre Ca, K e Mg.
Micronutrienti: Cl, Fe, Mn, B, Zn, Cu, Ni, Mb.
Il calcio regola la permeabilità delle membrane plasmatiche ed è importante anche per la
formazione delle pareti cellulari.
Il potassio è attivatore di enzimi, regola l'osmosi cellulare ed entra nel meccanismo di apertura delle
rime stomatiche.
Il magnesio è il componente della clorofilla e cofattore in diversi enzimi.
I microelementi sono spesso cofattori enzimatici. Il ferro è presente nei citocromi della catena
respiratoria.
Quando mancano alcuni elementi si manifestano sintomi da carenza, riconoscibili spesso a vista
oppure da ricercare in laboratorio con analisi accurate.
Gli elementi che più comunemente sono poco biodisponibili nei terreni e che necessitano di
somministrazioni sono N, P e K.
I concimi e i fertilizzanti che si trovano in commercio sono a base di uno di questi o di due o di tutti
e tre.
Carenza di N: scarso sviluppo vegetativo, colorazione sbiadita delle foglie. L'azoto è
presente nell'aria per il 78%: nel suolo è presente quindi nell'aria che riempie i pori del
terreno. Questo è in una forma non biodisponibile per le piante; l'unica forma biodisponibile
è come ione ammonio e come nitrato.
Carenza di P: crescita lenta, rami sottili e fragili, colorazione violacea sulle foglie. I terreni
hanno grandi quantità di fosforo, ma poco di quello disponibile per le piante, ovvero sotto
forma di ioni fosfato in soluzione.
Carenza di K: poco solubile nella soluzione circolante nel terreno, sintomi localizzati,
foglie gialle con punteggiature e tessuti necrotizzati.
Gli elementi minerali sono spesso assorbiti come ioni positivi cationi (Mg2+, K+, Ca2+): questi
spesso si scambiano con i cationi presenti nei foglietti dei minerali argillosi e da qui possono essere
difficilmente dilavati dalle piogge o dalle acque circolanti nel terreno. Le piante li assorbono
scambiandoli con ioni H+ che rilasciano nella soluzione circolante e sono quindi disponibili per i
peli radicali.
Gli anioni (come i nitrati) invece rimangono liberi e quindi corrono il grande rischio di essere
dilavati e percolare facilmente in profondità e giungere così alle acque superficiali e causare
inquinamenti.
AGRICOLTURA TRADIZIONALE E AGRICOLTURA BIOLOGICA
ORGANISMI OGM (schede pag 800-801)