Lezioneinvasioneemovimento
Movimento caratterizzato
dalla produzione di code di
actina o comete che
generano una forza
propulsiva per il movimento.
.
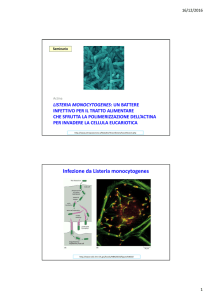
Listeria monocytogenes
• Listeria utilizza la proteina ACTA
per formare le comete. ACTA
agisce come mimo strutturale e
funzionale della proteina N-WASP,
attivando il complesso Arp2/3.
• Arp2/3 promuove l’assemblaggio
delle code di actina e la loro
reticolazione. Il dominio centrale di
ACTA ricco di ripetizioni di prolina
consente il legame con VASP.
• VASP recluta la profilina per
aggiungere monomeri di actina ai
filamenti in crescita.
• Nella fase finale le proteine
capping terminano la
polimerizzazione delle comete e le
cofiline iniziano la
depolimerizzazione.
Shigella flexneri
Shigella utilizza la proteina
IcsA per formare le comete.
IcsA non agisce da mimo
strutturale o funzionale di
NPFs eucariotici , ma si
lega direttamente ad NWASP per attivare il
complesso Arp2/3.
IcsA si comporta da mimo
funzionale di cdc42 per
potersi legare ad N-WASP.
il legame con VASP è
mediato dalla vinculina.
Fine polimerizzazione e
inizio depolimerizzazione
sono omologhe a Listeria.
• Rickettsia utilizza la proteina
RickA nella fase precoce del
movimento actina-dipendente.
• Si tratta di un mimo strutturale
e funzionale di N-WASP che
attiva il complesso Arp2/3.
• Nella fase tardiva del
movimento, RickA viene
sostituita da SCA2,una
formin-like protein, che agisce
direttamente come nucleatore
dell’actina.
Il movimento diventa indipendente dal complesso, infatti SCA2 da sola
funziona da NPF promuovendo l’assemblaggio delle comete, grazie
anche ai suoi domini ricchi di prolina.
•
Una sola proteina batterica ACTA
•
Un unico percorso di
•
ACTA agisce da mimo strutturale e funzionale di N-WASP
•
Attivazione complesso Arp2/3
•
Legame ACTA-VASP (proline)
•
Code di actina corte e reticolate
•
Una sola proteina batterica IcsA
•
IcsA non agisce da mimo strutturale o funzionale di NPFs eucariotici
ma si lega direttamente ad N-WASP (potremmo considerarlo un mimo
funzionale di Cdc42)
•
Attivazione del complesso Arp2/3
•
Legame IcsA-VASP (vinculine)
•
Code di actina corte e reticolate
•
Proteine capping e cofilina
•
Due proteine batteriche RickA e SCA2
•
Due distinte fasi di movimento e polimerizzazione delle comete
•
RickA agisce da mimo di N-WASP attivando il complesso Arp2/3
•
SCA2 agisce da mimo delle formins eucariotiche, da solo conduce
l’assemblaggio delle comete senza l’attivazione del complesso Arp2/3
polimerizzazione per la formazione delle comete
•
Legame SCA2-VASP (proline)
•
Code di actina lunghe e lineari
•
Proteine capping e cofilina
Dall’ABM alle protrusioni batteriche .…. il movimento intercellulare!
Protrusioni batteriche
Le protrusioni batteriche sono create alla
periferia cellulare quando il batterio, grazie
all’ABM, giunge alla membrana plasmatica
dell’ospite. La formazione delle protrusioni non è
una conseguenza passiva dell’ABM ma
coinvolge specifici fattori proteici.
Paracytophagy
Alla formazione delle protrusioni, segue il
processo di paracytophagy, attraverso il quale
una cellula avvolge e interiorizza le protrusioni
batteriche che si estendono dalle cellule
adiacenti.
Attraverso la sintesi di enzimi litici, l’agente
patogeno può liberarsi dal vacuolo e venire a
contatto con il citoplasma di una nuova cellula
ospite.
Questo processo consente ai batteri patogeni
di poter diffondere direttamente all’ interno di
una nuova cellula ospite , sfuggendo al sistema
immunitario e di conseguenza alla distruzione.
In seguito alla fagocitazione della protrusione, il
batterio risulta avvolto da un vacuolo con
doppia membrana.
Il fattore proteico responsabile della formazione
delle protrusioni in Listeria è InlC. Esso si lega al
dominio SH3 di tuba al posto di N-WASP allentando
la tensione corticale provocata dalle giunzioni
cellulari. Le proteine ERM aumentano la rigidità
delle protrusioni. La risoluzione delle protrusione e
la loro interiorizzazione nelle cellule adiacenti sono
ancora poco chiare ( probabile fattore coinvolto
Ck1-alfa)
Il fattore proteico responsabile delle formazione
delle protrusioni in Shigella è la Myosin x o
Myo10.
Myo10 aumenta la rigidità delle protrusioni e ne
favorisce l’allungamento.
Altre proteine coinvolte sono DIA1 e DIA2.
La risoluzione delle protrusione e la loro
interiorizzazione nelle cellule adiacenti avviene
tramite un’insolita endocitosi clatrina
dipendente, che coinvolge anche altri fattori
proteici quali Pi3k, epsin1 e dynamin2
La diffusione intercellulare non si
verifica in siti casuali di contatto
fra le cellule, piuttosto le
protrusioni batteriche tendono a
penetrare in angoli specifici.
In Listeria prevale la diffusione
attraverso le bTj o giunzioni
bicellulari, mentre in Shigella
prevale la diffusione attraverso le
tTj o giunzioni tricellulari.