METABOLISMO DEGLI
ACIDI GRASSI
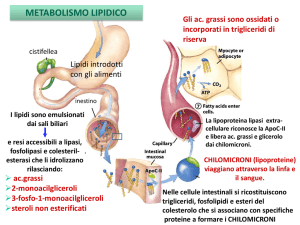
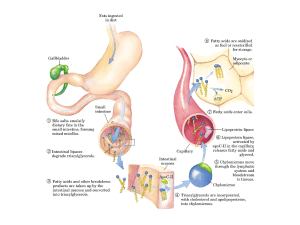
Lipidi introdotti
con gli alimenti
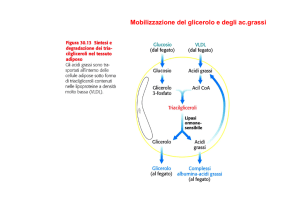
Mobilizzazione dei
trigliceridi di riserva
IDROLIZZATI DA LIPASI IN
GLICEROLO E ACIDI GRASSI
MOBILIZZAZIONE DEI LIPIDI DI
RISERVA Condizioni di bassa carica
energetica, digiuno (es: durante il
sonno)
Glucagone/adrenalina
Lipidi accumulati in gocce lipidiche:
nucleo di esteri del colesterolo e
trigliceridi, circondato da fosfolipidi
e proteine: le perilipine
in forma defosforilata
impediscono l’accesso alle lipasi
in forma fosforilata
interagiscono con la lipasi ormonesensibile e ne permettono
l’accesso nella goccia lipidica
la PKA fosforila sia le
perilipine sia la lipasi ormonesensibile attivando la
mobilizzazione degli ac. grassi
Glicerolo
Nelson·Cox – I PRINCIPI D BIOCHIMICA DI LEHNINGHER 5/E, Zanichelli Editore S.p.A. Copyright © 2010
Fegato
Fegato, cuore, muscolo, corteccia renale
Trasportati nel circolo
sanguigno attraverso
l’albumina serica
Berg et al. BIOCHIMCA 6/E, ZANICHELLI
EDITORE S.p.A. Copyright 2007
Il 5% dell’energia rilasciata dai trigliceridi deriva dal Glicerolo:
Il GLICEROLO è fosforilato nel fegato dalla Glicerolo chinasi
in GLICEROLO 3-FOSFATO
È ossidato dalla Glicerolo 3-fosfato deidrogenasi (NAD-dipendente)
in DIIDROSSIACETONEFOSFATO
che isomerizzato a GLICERALDEIDE 3-FOSFATO entra nella glicolisi
(o nella gluconeogenesi).
La gran parte dell’energia rilasciata dai trigliceridi deriva
dall’ossidazione degli acidi grassi.
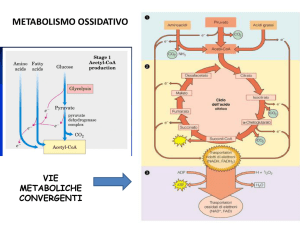
BETA-OSSIDAZIONE DEGLI ACIDI GRASSI
Produce grandi quantità di Acetil-CoA,
che alimenta il ciclo di Krebs e di
conseguenza la fosforilazione
ossidativa.
Produce anche grandi quantità di
NADH e FADH2 che sono incanalati
nella catena respiratoria di trasporto
degli e-
LA PRIMA FASE DELLA βOSSIDAZIONE AVVIENE NEL
CITOSOL
E
SERVE
AD
ATTIVARE L’ACIDO GRASSO
L’acido grasso viene ADENILATO
(Anidride mista)
pirofosfatasi
α
L’acido grasso deve essere
attivato per mezzo della
ACIL-CoA SINTETASI
(famiglia di isozimi specifici
per ac. grassi a catena corta,
media o lunga)
L’AMP dell’acil-denilato è
spiazzato dal CoA-SH e si
forma un legame TIOESTERE
(ACIL-CoA)
AMP + ATP → 2ADP
Sono consumati
in totale 2 ATP
Nelson-Cox - I principi di Biochimica di Lehninger – Zanichelli editore spa Copyright 2014
Gli ac. grassi attivati devono essere trasferiti nel mitocondrio, dove si
trovano tutti gli enzimi deputati alla loro ossidazione. Nelle piante il
sito principale della β-ossidazione degli ac. grassi sono i perossisomi.
Ac. grassi con catene non più lunghe di 12 atomi di carbonio:
attraversano la membrana mitocondriale interna.
Ac. grassi con catene più lunghe necessitano di un sistema
navetta costituito dalla CARNITINA
carnitina
-C=O
l
R
carnitina
aciltrasferasi I
HS-CoA
Acil-carnitina
Il trasferimento degli ac.grassi via carnitina dentro il mitocondrio è la
tappa limitante di tutto il processo di ossidazione degli ac.grassi
I pools di CoA citosolico e mitocondriale sono tenuti separati
Il CoASH citosolico serve per i processi di sintesi (Biosintesi acidi grassi).
Il CoASH mitocondriale per i processi catabolici (decarbossilazione ossidativa del
piruvato; ossidazione ac. grassi e di alcuni aminoacidi)
Nelson-Cox - I principi di Biochimica di Lehninger – Zanichelli editore spa Copyright 2014
β-OSSIDAZIONE di un Ac. Grasso Saturo con n°PARI di atomi di carbonio
Deidrogenazione FAD-dipendente del legame CαCβ per mezzo della Acil-CoA deidrogenasi che è
legata alla membrana mitocondriale interna)
ETF = flavoproteina
trasportatrice di elettroni
Trans-∆2enoil-CoA
Idratazione (introduciamo un gruppo –
OH sul Cβ) per mezzo della enoil-CoA
idratasi
β-idrossi-acil-CoA
D. Voet, J.G. Voet, C.W. Pratt. FONDAMENTI DI BIOCHIMCA 2/E, ZANICHELLI EDITORE S.p.A. Copyright 2007
Ossidazione del Cβ per mezzo della
β-idrossiacil-CoA deidrogenasi NADdipendente. Si forma NADH.
al Complesso I
α
CoASH
Il Cα si trova fra 2 gruppi C = O, ciò
rende meno stabili i legami C-C del Cα.
La funzione chetonica sul Cβ lo rende
un buon elettrofilo suscettibile
all’attacco del gruppo tiolico del CoASH
S-CoA
al Ciclo di Krebs
Per gli ac. grassi con 12 o più atomi di carbonio le ultime 3 tappe sono catalizzate da un
complesso multienzimatico associato alla membrana mitocondriale interna (PROTEINA
TRIFUNZIONALE –TFP-), che consente un incanalamento dei substrati da un sito attivo all’altro.
Quando la catena si accorcia a < 12 C, il processo è seguito dagli enzimi della matrice.
D. Voet, J.G. Voet, C.W. Pratt. FONDAMENTI DI BIOCHIMCA 2/E, ZANICHELLI EDITORE S.p.A. Copyright 2007
C16
Per ossidare completamente una molecola di
palmitato (16 atomi di C) occorrono 7 cicli di βossidazione e sono rilasciate 8 molecole di AcetilCoA.
Inoltre complessivamente sono prodotti:
7 FADH2 e 7 NADH
se: FADH2/1.5 ATP;
NADH/2.5 ATP
Fosforilazione ossidativa >> 10.5 ATP + 17.5 ATP = 28 ATP
8 Acetil-CoA >> Krebs >> 8 GTP (ATP) + 24 NADH + 8 FADH2
80 ATP
Fosforilazione ossidativa >> 60 ATP + 12 ATP
TOT = 80 + 28 ATP – 2 (consumati per l’attivazione dell’ac. grasso)
106 ATP
(da 1 glucosio se ne ottengono 36 o 38)
β-OSSIDAZIONE DI UN ACIDO GRASSO DISPARI
L’ultimo ciclo rilascia 1 Acetil-CoA e
1 Propionil-CoA (unità tricarboniosa)
Convertito in Succinil-CoA
β
α
Propionil-CoA
carbossilasi
ciclo di Krebs
α
Metilmalonil-CoA
mutasi
(cobalammina)
+ biotina
α
Metilmalonil-CoA
epimerasi
Diventa L
CHETOGENESI (PRODUZIONE DI CORPI CHETONICI)
Bassa glicemia (es.: digiuno) o di Diabete mellito non trattato (insufficiente captazione
insulino-dipendente di glucosio)
La glicolisi è inibita ma
la β-ossidazione è
attiva e continua a
produrre grandi
quantità di acetil-CoA
Accumulo di Acetil-CoA.
Viene accelerata la
gluconeogenesi epatica
e renale che sottrae
ossalacetato al ciclo di
Krebs bloccandolo.
In condizioni di bassa glicemia sotto effetto del
GLUCAGONE attraverso la via cAMP/PKA è
attivata la mobilizzazione dei trigliceridi di riserva
che libera GLICEROLO e ACIDI GRASSI
I corpi chetonici sono molecole carburante
che rilasciate nel torrente circolatorio
riforniscono di energia metabolica le cellule
cerebrali, cardiache e della corteccia renale
Berg et al, BIOCHIMICA 6/E, Zanichelli editore S.p.a. Copyright © 2007
2 molecole di Acetil-CoA sono condensate
liasi
1 Acetil-CoA è
rilasciato
β-chetolasi
α
β
2
1
Deidrogenasi
HMG-CoA
sintasi
β
Una 3a molecola di AcetilCoA è condensata
Sono resi disponibili 2 CoA-SH e 1 NAD+
Acetoacetato e idrossibutirrato entrano
nei mitocondri delle cellule che li
utilizzano, dove vengono convertiti in
Acetil-CoA
Complesso II
(viene riattivata
la fosforilazione
ossidativa)
Ciclo di Krebs
Fumarato
Malato
Ossalacetato
VIENE
RIATTIVATO IL
CICLO DI KREBS
OO
OO
OO-
D. Voet, J.G. Voet, C.W. Pratt. FONDAMENTI DI BIOCHIMCA 2/E, ZANICHELLI EDITORE S.p.A. Copyright 2007
SINTESI DEGLI ACIDI GRASSI
>> AVVIENE NEL CITOSOL. Nei mammiferi principalmente negli
epatociti e negli adipociti.
>> per sintetizzare acidi grassi è necessario avere nel citosol:
Acetil-CoA,
Malonil-CoA
NADPH
ATP
>> l’Acetil-CoA è prodotto nel mitocondrio ma in condizioni di
accumulo energetico viene traslocato fuori dal mitocondrio sotto
forma di citrato con un processo che è associato alla riduzione di
NADP+ a NADPH.
>> Il malonil-CoA è prodotto nel citosol a partire dall’acetil-CoA.
>> NADPH: prodotto in grandi quantità nella via dei pentosio-fosfati
CITRATO
consumo
di ATP
OSSALACETATO + ACETIL-CoA
1a reazione del
ciclo di Krebs
consumo
di ATP
malato
deidrogenasi
citosolica
enzima malico
(riduzione di NADP+)
Berg et al, BIOCHIMICA 6/E, Zanichelli editore S.p.a. Copyright © 2007
Nelson·Cox – I PRINCIPI D BIOCHIMICA DI LEHNINGHER 5/E,
Zanichelli Editore S.p.A. Copyright © 2010
SINTESI DI MALONIL-CoA
biotina
Acetil-CoA
carbossilasi
SONO CONSUMATE COMPLESSIVAMENTE:
• 2 MOLECOLE DI ATP PER OGNI ACETIL-CoA CHE E’ TRASPORTATO
FUORI DAL MITOCONDRIO
• 3 MOLECOLE DI ATP PER OGNI MALONIL-CoA CHE VIENE
SINTETIZZATO DALLA ACETIL-CoA CARBOSSILASI
Un gruppo malonilico e un gruppo acetilico vengono trasferiti sul
complesso enzimatico dell’ACIDO GRASSO SINTASI
nei mammiferi è un polipeptide con 6 domini ciascuno dei quali ha
una sua attività enzimatica
(ER) Enoil-ACP
reduttasi
(DH) βidrossiacil-ACP
deidratasi
(KS) β-chetoacil
-ACP sintasi
(KR) β-chetoacil-ACP
reduttasi
(ACP) proteina
trasportatrice di acili
(MAT) Malonil/acetil-CoA-ACP
transferasi
(ACP) proteina trasportatrice di acili
(KS) β-chetoacil
-ACP sintasi
(MAT) Malonil/acetil-CoA-ACP transferasi
L’Acetile viene legato al
gruppo tiolico di un residuo di
Cys del dominio KS (chetoacilACP sintasi). Il malonile viene
caricato sull’ACP per mezzo del
dominio MAT
Malonil-CoA –ACP
Trasferasi (MAT)
ACP (Acyl Carrier Protein) = coenzima proteico contenente un residuo di
fosfopanteteina, possiede dunque un gruppo reattivo SH con cui forma un
legame tioestere con il malonile
S KS
̶ KS
KS
CONDENSAZIONE
Catalizzata dalla
β-chetoacil-ACP-sintasi
(KS)
CO
KS2
S ̶ ACP
VIENE ELIMINATA
CO2 E SI FORMA
UN β-CHETOACILE
CHE RIMANE
LEGATO ALL’ACP
β-chetobutirril-ACP
β
Viene ridotto il gruppo carbonile β
a spese del NADPH
β-idrossibutirril-ACP
β-chetobutirril-ACP
RIDUZIONE
Catalizzata dalla
β-chetoacil-ACPreduttasi (KR)
β
β
α
H2O
DEIDRATAZIONE
β-idrossiacil-ACP
deidratasi (DH)
trans-Δ2-butenoil-ACP
trans-Δ2-Butenoil-ACP
Viene ridotto il doppio legame
a spese del NADPH
Riduzione catalizzata dalla
Enoil-ACP-reduttasi (ER)
NADPH + H+
NADP+
Butirril-ACP
La catena acilica (4 atomi di
C) viene trasferita dal
braccio mobile dell’ACP sul
gruppo SH del residuo di
cisteina del dominio KS
Il braccio mobile dell’ACP, di nuovo libero, può accogliere una successiva
molecola di malonil-CoA e iniziare un nuovo ciclo di allungamento
L’allungamento, in genere procede sino a 16 atomi di C e il principale
prodotto dell’ACIDO GRASSO SINTASI è il palmitato (16 atomi di C, saturo).
Quando la sintesi è completata l’acido grasso è rilasciato mediante una
tioesterasi.
Il palmitato a sua volta diventa
precursore per la sintesi di ac.
grassi a catena più lunga: il
sistema di allungamento agisce
nel reticolo endoplasmatico liscio
aggiungendo unità acetiliche
Le insaturazioni negli ac. grassi
insaturi sono introdotte da una
serie di DESATURASI specifiche
che agiscono in sequenza
(reticolo endoplasmatico liscio)
Nelson·Cox – I PRINCIPI D BIOCHIMICA DI LEHNINGHER 5/E, Zanichelli
Editore S.p.A. Copyright © 2010
La degradazione degli acidi grassi così come la loro biosintesi è fortemente
regolata
• attiva solo quando la cellula
richiede energia
• è bloccata quando è attiva la biosintesi
1) La mobilizzazione di acidi grassi dalle riserve di trigliceridi è sotto
controllo ormonale (glucagone e adrenalina la stimolano)
2) La produzione di MALONIL-CoA è sotto controllo ormonale, infatti
l’Acetil-CoA Carbossilasi (ACC) è regolata dal sistema PKA/proteinafosfatasi 2A
PKA
Quando l’ACC è attiva (sotto controllo insulinico) viene prodotto
malonil-CoA in gran quantità. Una parte di questo malonil-CoA
funge da INIBITORE della carnitina-Aciltrasferasi I → è inibita la βossidazione degli ac. grassi.
L’Acetil-CoA carbossilasi è regolata anche dal:
- palmitoil-CoA (inibitore retroattivo)
- citrato (attivatore allosterico che ne aumenta la Vmax)
quando la produzione mitocondriale di Acetil-CoA e ATP aumenta
eccessivamente, il citrato è esportato nel citosol.
1) diventa precursore dell’Acetil-CoA citosolico
2) attiva l’Acetil-CoA carbossilasi accelerando la sintesi
di ac.grassi.
3) inibisce la fosfofruttochinasi-1, rallentando la glicolisi