
La regolazione dell’espressione genica
Tutte le cellule di un organismo multicellulare possiedono tutti i geni della propria
specie; tuttavia solo una piccolissima frazione è attiva in ogni cellula; per esempio, in
una cellula delle isole del pancreas è attivo il gene per l’insulina, non quelli per
l’emoglobina (attivi nelle cellule dei midollo osseo, progenitrici dei globuli rossi) o per la
pepsina (attivi nelle cellule della mucosa gastrica)
Durante lo sviluppo embrionale di un organismo multicellulare, le cellule destinate a
costituire i diversi tessuti e organi attivano in momenti precisi i geni che codificano le
proteine che stimolano o inibiscono la proliferazione cellulare e quelle caratteristiche del
tessuto in formazione (per esempio osseina per le ossa, cheratina per la pelle)
Anche gli organismi unicellulari hanno bisogno di attivare alcuni geni solo in particolari
circostanze (per esempio la presenza di un nutrimento che richiede particolari enzimi)
per non sprecare l’energia e le molecole necessarie
Le cellule tumorali hanno perduto, anche in seguito a mutazioni somatiche, la capacità
di regolare l’espressione genica relativa alle proteine che controllano la proliferazione
cellulare
Dunque nelle prossimità dei geni ci devono essere recettori in grado di percepire segnali
provenienti dall’ambiente esterno e veicolati all’interno della cellula; tali recettori
devono quindi segnalare ai geni se trascrivere o no, in funzione dei segnali provenienti
dall’esterno
L’esempio che viene proposto nelle diapositive 2-5 riguarda la regolazione dell’attività
dei geni (detta “espressione genica”) relativa agli enzimi per il metabolismo del
galattosio in Escheirichia coli (sistema lac)
Il sistema lac in E. coli
HOC6’H2
C5’
OH
O
O
H
C4’
H
Lattosio
OH
H
C3’
C2’
H
OH
C1’
C4’
H
H
OH
C2’
OH
H
H
H
OH
C5’
O
HOC6’H2
b-galattosidasi
O
C5’
OH
H
C1’
OH
H
C3’
H
Galattosio
C3’
HOC6’H2
C4’
H
OH
C1’
HOC6’H2
C5’
H
H
H
O
L’enzima b-galattosidasi,
che consente di utilizzare
il lattosio come fonte di
carbonio e di energia,
viene sintetizzato in
presenza di lattosio che
quindi oltre che esserne il
substrato è anche
l’induttore della sua
sintesi
OH
H
C4’
OH
+ H2O
C1’
OH
H
C2’
C3’
C2’
OH
H
OH
Glucosio
H
Dal lattosio è indotta
la sintesi di altri 2
enzimi: permeasi e
transacetilasi; anche la
permeasi è coinvolta
nel metabolismo del
lattosio, poiché facilita
l’ingresso del lattosio
nella cellula.
I geni strutturali e il gene I
I
Z
Y
I+ = allele normale:Z, Y, A trascrivono in
presenza dell’induttore;
I- = mutante costitutivo: Z, Y, A trascrivono
sempre;
Is = allele inibente la risposta all’induttore:
Z, Y, A non trascrivono mai
A
trascrizione
Merodiploidi per definire le relazioni funzionali
tra gli alleli
Un unico mRNA policistronico
I+Z-/I-Z+
traduzione
I-Z-/I+Z+
b-galattosidasi
transacetilasi
I-Z-/I-Z+
Trascrive*
sempre
IsZ+/I+Z+
Non trascrive
mai
IsZ+/I-Z+
Non trascrive
mai
F’(lac)
permeasi
Geni lac
1) Is è dominante su I+ che a sua volta è
dominante su I-
Trascrive* con
l’induttore
Trascrive* con
l’induttore
*un mRNA per un enzima funzionale
2) I+ è dominante su I- indipendentemente dalla posizione in cis o in trans rispetto a Z+
Operatore, promotore e operone
I
P
O
Z
Y
A
operone
O+ = allele normale:Z, Y, A trascrivono in presenza dell’induttore;
Oc = mutante costitutivo: Z, Y, A trascrivono sempre
O+Z+/OcZ+
Trascrive* sempre
O+Z+/OcZ-
Trascrive* con
l’induttore
O+Z-/OcZ+
Trascrive* sempre
IsO+Z+/I+OcZ+
Trascrive* sempre
Il prodotto del gene I è una
proteina diffusibile che, in assenza
dell’induttore, si lega ad O e
impedisce la trascrizione di Z, Y ,
A; in presenza dell’induttore non
si lega ad O e consente la
trascrizione di Z, Y, A
*un mRNA per un enzima funzionale
1) Oc è dominante su O+, ma solo in cis rispetto a Z+ (cis-dominanza)
2) Gli alleli normali di P sono cis-dominanti sui mutanti difettivi di P, che
impediscono incondizionatamente la trascrizione di Z, Y, A, impedendo
l’inserimento della RNA polimerasi
Regolazione dell’espressione
genica nell’operone lac
RNA polimerasi
I
P
O
Z
Y
A
Y
A
induttore
mRNA
I-
I
Is
Oc
P-
P
O
Z
repressore non legato
all’induttore, che si lega
ad O e impedisce la
trascrizione
repressore legato
all’induttore, che
non si lega ad O e non
impedisce la trascrizione
repressore I- senza affinità
per O, con cui non si lega
mai e non impedisce mai
la trascrizione
I
P
O
Z
Y
A
repressore Is senza affinità
per l’induttore, che si lega
sempre ad O e impedisce
sempre la trascrizione
I
P
O
Z
Y
A
operatore senza affinità
per il repressore che non
lega mai e non impedisce
mai la trascrizione
I
P
O
Z
Y
A
promotore senza affinità
per l’RNA polimerasi che
impedisce sempre la
trascrizione
Ingegneria genetica
Le tecniche dell’ingegneria genetica permettono di isolare un gene che codifica per una determinata
proteina e di inserirlo, mediante dei vettori adatti, nel genoma di un altro organismo. Queste
tecniche sono state applicate al campo della medicina per la produzione farmaci, per la diagnosi di
malattie genetiche, per la progettazione di vaccini. In agricoltura si stanno progettando piante e
animali transgenici, per aumentarne la produttività e migliorarne la qualità.
Ma tutto ciò pone problemi nuovi di carattere etico, economico, biologico che devono essere
attentamente valutati per la tutela della vita e dell’ambiente.
Batterio
Cromosoma
batterico
Plasmide isolato
Gene da isolare
Plasmide
DNA ricombinante
(plasmide)
Inserimento del
plasmide nella
cellula batterica
Copie del gene
isolato
Cellula
contenente il
gene da
isolare
DNA purificato
Batterio
ricombinante
duplicazione
Copie delle proteine come
ad esempio insulina, ormone
della crescita
Enzimi di restrizione
Gli enzimi di restrizione sono endonucleasi capaci di tagliare il DNA producendo
rotture a doppia elica in corrispondenza di sequenze specifiche di nucleotidi (siti di
restrizione); di essi si servono i batteri per distruggere i cromosomi virali
endonucleasi
Sito di restrizione
EcoRI
5’GAATTC
3’CTTAAG
HaeIII
5’CC GG
3’GGCC
Formazione di code
5’CCAA
3’GG
GG
TT CC
Tipo di taglio
Sfalsato, adesivo
Tronco, non adesivo
Un taglio è detto adesivo
quando:
1) È sfalsato, o sono
aggiunti nucleotidi
al 3’;
2) I nucleotidi a singolo
filamento, sfalsati o
aggiunti, sono fra
loro complementari
Le estremità di un taglio adesivo si possono unire
mediante i legami idrogeno tra le basi complementari
presenti a singolo filamento sui 2 segmenti di DNA;
successivamente, mediante l’azione di ligasi e, se serve,
di DNA polimerasi, si saldano i filamenti
polinucleotidici (vedere diapositiva 8)
Si possono aggiungere al 3’ di uno dei 2 filamenti di un capo di un taglio tronco, non
“adesivo”, una breve sequenza, ripetizione di un solo nucleotide e al 3’ dell’altro capo la
sequenza complementare, rendendo adesivo il taglio
I siti di restrizione sono brevi sequenze palindromiche: la sequenza posta a un lato
rispetto al centro del sito è complementare alla sequenza inversa rispetto a quella posta al
lato opposto
Mappe di restrizione
Una mappa di restrizione consiste nell’identificazione della successione dei
siti di restrizione per diverse endonucleasi in un segmento di DNA,
misurando la distanza tra i siti come numero di nucleotidi
1) Si esamina un
frammento di DNA,
marcandolo con 32P al 5’
2) Si digerisce il frammento con
diversi enzimi di restrizione
Marcatura con con 32P
3) Si stima la lunghezza dei
frammenti dallo spazio
percorso su un gel
mediante elettroforesi
Siti di restrizione
dell’enzima
Siti di restrizione
dell’enzima
L’elettroforesi consiste nel collocare le macromolecole in un
pozzetto all’estremità di un gel (gelatina) che viene poi sottoposto
un campo elettrico; le molecole si spostano con una velocità
proporzionale alla propria carica elettrica e inversamente
proporzionale lla propria lunghezza; poiché nel DNA la carica è
distribuita uniformemente, la velocità dipende solo dalla lunghezza
del frammento
Il DNA ricombinante
DNA da clonare
vettore
VETTORI
Sono cromosomi
capaci di replicare e
selezionabili, in cui si
possono inserire
segmenti di DNA di
interesse
Digestione con
endonucleasi
DNA polimerasi I +
ligasi
I vettori possono essere :
-plasmidi,
-cromosomi di fagil,
ibridi fago-plasmide (cosmidi),
tutti capaci di replicarsi in E. coli,
-YAC, cioè cromosomi artificiali di
lievito, capaci di replicarsi in lievito
In ogni tipo di vettore si può
inserire un frammento di DNA di
una particolare lunghezza massima
Estremità adesive spontanee o
costruite, fra loro uguali
Estremità adesive
complementari alle precedenti
La clonazione del DNA
Individuazione dei geni
clonati: Southern blotting
Genoma da saggiare
frammentato con
enzimi di restrizione
Cromosomi di
fago l
frammentati
1) Si effettua l’elettroforesi
su gel dei frammenti a
doppia elica
ligasi
Genoteca di DNA
ricombinante
Clonazione
Ibridazione su filtro
con sonda radioattiva
complementare al
DNA da studiare
2) Si denatura il DNA e lo si fa
aderire a singolo filamento,
nelle stesse posizioni, a un
filtro
3) Si ibridizza con sonda
radioattiva complementare al
frammento cercato e si effettua
l’autoradiografia, localizzando
così il gene studiato
Sequenziamento
SEQUENZIAMENTO DEI NUCLEOTIDI
1) Si marca con 32P l’estremità 5’ del frammento da analizzare, quindi si denatura il DNA
2) Si distruggono
selettivamente basi o
loro combinazioni G
(G, G+A, C, C+T),
che rendono più
fragile il filamento
saggiato, che quindi
si rompe in quei
punti
3) Si stima la lunghezza
dei frammenti dallo
spazio percorso in una
corsa elettroforetica
G
C
A
T
G+A
C
C+T
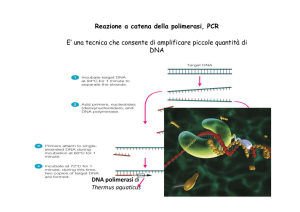
Reazione a catena della polimerasi (PCR)
PCR
Taq polimerasi, resistente al caldo
Primer 1
Regione complementare a
Primer 2
Regione complementare a
Si alternano cicli di denaturazione (a temperatura alta) e di
replicazione ( a temperatura bassa), ciascuno dei quali
porta un raddoppiamento del numero delle molecole
Costruire nuovi geni
SINTESI DI SEQUENZE NON SU STAMPO:
è possibile allungare filamenti di DNA al 5’ aggiungendo nucleotide
dopo nucleotide costruendo oligonucleotidi di sequenza voluta
INDUZIONE DI MUTAZIONI SPECIFICHE
1) Si rompe il DNA
con endonucleasi
2) Si digeriscono singoli
filamenti in direzione 5’
con esonucleasi
3) Si modificano specifici nucleotidi sul filamento
rimasto; dopo la replicazione, anche il nuovo
filamento sarà mutato nello stesso sito




![mutazioni genetiche [al DNA] effetti evolutivi [fetali] effetti tardivi](http://s1.studylibit.com/store/data/004205334_1-d8ada56ee9f5184276979f04a9a248a9-300x300.png)






![(Microsoft PowerPoint - PCR.ppt [modalit\340 compatibilit\340])](http://s1.studylibit.com/store/data/001402582_1-53c8daabdc15032b8943ee23f0a14a13-300x300.png)