MECCANISMI DI
TRASPORTO ATTRAVERSO
LA MEMBRANA
FUNZIONI DELLA MEMBRANA PLASMATICA
1) ricevere
informazione
2) importare ed
esportare molecole
3) capacità di
movimento ed
espansione
FUNZIONI DELLA MEMBRANA PLASMATICA
• Regola il trasporto dei nutrienti all’interno della cellula
• Regola il trasporto dei prodotti di degradazione verso l’esterno
• Mantiene “adeguate” condizioni chimiche nella cellula
• Fornisce un sito per le reazioni che avvengono raramente in ambiente
acquoso
• Contribuisce alla trasduzione del segnale
• Interagisce con altre cellule e con la matrice extracellulare
LIPIDI:
PROTEINE:
- ruolo strutturale
- ruolo funzionale
testa
idrofilica
coda
idrofobica
In acqua le interazioni delle code idrofobiche e
delle teste
idrofiliche generano un doppio strato fosfolipidico. Le teste sono
dirette verso l’esterno, dove interagiscono con l’acqua che le
circonda. Le code sono rivolte verso l’interno.
MODELLO A MOSAICO FLUIDO :
proteine e lipidi
continuazione.
variano
la
loro
disposizione
spaziale
Le proteine sono sospese singolarmente in un doppio strato fluido.
Non tutte le proteine sono capaci di diffondere liberamente
attraverso la membrana.
RUOLI SPECIALIZZATI
in
In alcuni casi , la mobilità delle proteine è limitata dalla loro
associazione con il citoscheletro (es. banda 3 neinglobuli rossi).
Quasi tutte le membrane cellulari sono rinforzate e sostenute da
un’impalcatura
proteica,
fissata
per
mezzo
di
proteine
transmembrana che si organizzano in una trama di proteine fibrose
(lo strato corticale o CORTEX) aderente alla faccia citosolica. Il
cortex determina la forma cellulare e le proprietà meccaniche della
membrana plasmatica.
LIPIDI DI MEMBRANA
1)
FOSFOGLICERIDI
(FOSFOLIPIDI:
molecole
anfipatiche,
rappresentano i principali lipidi delle membrane biologiche)
- colina
- glicerolo
- etanolammina
- inositolo
- serina
- treonina
-SRUTTURALI: es. fosfatidil etanolammina
glicerolo
-FUNZIONALI: es. fosfatidil-inositolo
Fosfolipasi C
acido grasso
fosfato
INOSITOLO-3P + DIACIL GLICEROLO
2) GLICOLIPIDI DI MEMBRANA
batteri e piante:
glicolipidi basati sul diacil glicerolo
gal
gal
glu
acido grasso
glicerolo
animali:
SFINGOLIPIDI, costituiti da un alcool a lunga catena denominato
SFINGOSINA
colina
glu
fosfato
SFINGOSINA
SFINGOMIELINA
serina
acido grasso
serina
acido grasso
acido grasso
serina
GLICOSFINGOLIPIDI
- particolarmente abbondanti nel sistema nervoso centrale nelle
membrane dei neuroni
- fanno legami fortissimi altamente ramificati tra loro, quindi
servono a stabilizzare la struttura delle membrane
- difetti nel loro metabolismo sono causa di malattia genetica con
paralisi e ritardo mentale
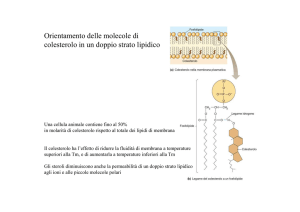
3)STEROLI
Il COLESTEROLO serve a mantenere fluide le membrane (anche
a basse T):
- impedisce il “congelamento” delle membrane;
- aumenta la flessibilità e la stabilità meccanica
La struttura e la funzione di tutte le membrane cellulari dipende
fondamentalmente dai fosfolipidi e da derivati degli steroidi.
Le specifiche funzioni di ciascuna membrana dipendono dal tipo di
proteine presenti su quella specifica membrana.
LE PROTEINE DI MEMBRANA
1) PROTEINE
PERIFERICHE:
proteine che si staccano dalla membrana in
seguito a trattamento con reagenti polari che
non distruggono il doppio strato fosfolipidico.
Sono associate alla membrana tramite
interazioni non covalenti PROTEINAPROTEINA.
2) PROTEINE
INTEGRALI:
proteine che possono essere rilasciate dalla
membrana solo in seguito a trattamenti che
distruggono il doppio strato fosfolipidico.
-TRANSMEMBRANA
-ASSOCIATE ALLA MEMBRANA
-LEGATE AI LIPIDI
Le proteine di membrana, come quelle citoplasmatiche, contengono
circa il 50% di amminoacidi idrofobici.
Gli amminoacidi idrofobici nelle proteine di membrana sono organizzati
in DOMINI TRANSMEMBRANA
NH2
dominio transmembrana
idrofobico
COOH
20-25 aa.
Struttura ad -elica
COOH
NH2
domini transmembrana
Pochi foglietti b attraversano le membrane
Le porzioni extracellulari delle proteine della membrana plasmatica
sono generalmente glicosilate.
-GLICOPROTEINE:
legano brevi catene di zuccheri. Sono
abbondanti nella membrana plasmatica,
scarse nelle membrane interne (di solito si
trovano nel lume di RE e Golgi)
Glucide legato a NH2 di ASN
legame
N-GLICOSIDICO
Glucide legato a OH di SER/THR
legame N-GLICOSIDICO
-PROTEOGLICANI:
legano una o più catene saccaridiche lunghe.
Tutti i carboidrati delle glicoproteine si trovano nello stesso lato
della membrana, quello non citosolico, e contribuiscono a formare
un involucro glucidico detto GLICOCALICE.
-stabilità alle membrane
-inibizione delle proteasi extracellulari
-conformazione 3D delle membrane
-siti di riconoscimento
I glucidi sulle proteine possono legarsi in maniera lineare o ramificata: le strutture
dell’asse centrale dei gruppi glucidici vengono aggiunti nel RE, le ramificazioni nel Golgi.
rivestimento
glucidico
membrana plasmatica
vescicole secretorie
complesso di Golgi
vescicole di transizione
reticolo endoplasmico ruvido (RER)
glicoproteina di membrana
gruppo glucidico
polipeptide
MECCANISMI DI TRASPORTO
Le membrane cellulari regolano il passaggio di tutte le sostanze in
transito fuori e dentro la cellula e all’interno dei diversi compartimenti
intracellulari.
La membrana
plasmatica è una
barriera selettiva
DIFFUSIONE SEMPLICE
DIFFUSIONE FACILITATA
o
TRASPORTO PASSIVO
TRASPORTO ATTIVO
X GRADIENTE DI
CONCENTRAZIONE
(non richiede energia)
CONTRO GRADIENTE
(consumo di energia)
Avvengono su tutte le membrane cellulari: plasmatiche, mitocondriali,
nucleari, Golgi, RE
DIFFUSIONE SEMPLICE:
100
molecole
10
molecole
55
molecole
55
molecole
EQUILIBRIO
-non selettiva
-non saturabile
-porta all’equilibrio
-passano molecole che si
dissolvere in membrana
possono
-gas (CO2 e O2)
-molecole idrofobiche (benzene)
-piccole molecole polari prive di
carica (H2O e etanolo)
DIFFUSIONE FACILITATA o TRASPORTO PASSIVO
come la DIFFUSIONE SEMPLICE + CARRIER
Le molecole trasportate non si sciolgono nel doppio strato
fosfolipidico. Il loro passaggio è mediato da proteine che gli
consentono di attraversare la membrana senza interagire con
l’interno idrofobico.
Molecole polari e
amminoacidi e ioni)
con
carica
(carboidrati,
nucleosidi,
I CARRIERS sono proteine integrali di membrana
- saturabili
- specifiche
1) PROTEINE TRASPORTATRICI
(zuccheri, aa e nucleosidi)
2) PROTEINE CANALE
(ioni e piccole molecole)
TIPI PRINCIPALI DI PROTEINE DI TRASPORTO
CANALI: trasportano H2O e specifici tipi di ioni secondo gradiente di
concentrazione. Le proteine che li compongono formano dei canali
che attraversano la membrana. Sono di solito regolati da stimoli
specifici.
TRASPORTATORI: legano ioni o molecole specifiche. Il legame con la
molecola trasportata provoca un cambiamento conformazionale e
quindi il passaggio.
- Uniporto - Simporto - Antiporto
POMPE: usano l’energia da idrolisi dell’ATP per spostare ioni contro
gradiente di concentrazione
1) PROTEINE TRASPORTATRICI: es. trasp del GLUCOSIO
(uniporto GLUT1)
Il trasporto avviene secondo gradiente:
- nella maggior parte delle cellule il glucosio viene trasportato dentro
la cellula
- nelle cellule epatiche il glucosio viene esportato
Una volta entrato nella cellula il Glu viene fosforilato e non è più in
grado di attraversare la membrana.
L’uniporto accelera una reazione comunque termodinamicamente
favorita (meccanismo simile a quello degli enzimi)
Tre principali caratteristiche distinguono l’uniporto dalla diffusione
passiva:
•la velocità della diffusione facilitata è molto maggiore di quella della
diffusione passiva
•il trasporto è specifico
•il trasporto avviene attraverso un numero limitato di “porte”
2) CANALI IONICI:
-estrema rapidità (>106ioni/s)
-selettività
-azione regolabile (apertura
chiusura mediate)
e
3 TIPI DI CANALE AD ACCESSO CONTROLLATO:
canale a CONTROLLO DI POTENZIALE (variazioni di voltaggio di
membrana)
canale a CONTROLLO DI LIGANDO (es. ormoni,
neurotrasmettitori)
canale a CONTROLLO MECCANICO (stimolazioni meccaniche)
CANALE A CONTROLLO DI POTENZIALE
(voltaggio dipendenti)
I canali di K+, Na+ e Ca2+ appartengono ad una famiglia di proteine correlate.
Il canale del K+ è formato dall’associazione di quattro subunità identiche. Il
canale del Na+ consta di una singola catena polipeptidica contenente quattro
domini ripetuti, ognuno dei quali è simile ad una subunità del canale del K+.
Ogni subunità o dominio contiene 6 -eliche che attraversano la membrana.
L’-elica 4 contiene molti aa carichi + e agisce come sensore di voltaggio che
media l’apertura del canale in risposta a variazioni del potenziale di
membrana.
CANALE A CONTROLLO DI LIGANDO
Es. recettore nicotinico dell’acetilcolina muscolare: proteina
transmembrana costituita da 5 subunità(2, 1b, 1 g e 1 d). In seguito al
legame di due molecole di aceticolina alle subunità si ha una
modificazione conformazionale con conseguente apertura del canale.
Il flusso degli ioni attraverso i canali di membrana dipende dallo
stabilirsi di un gradiente ionico attraverso la membrana plasmatica.
Tutte le cellule possiedono una pompa ionica che usa l’E dell’ATP per
trasportare attivamente gli ioni attraverso la membrana plasmatica.
Come conseguenza, la composizione ionica del citoplasma è
sostanzialmente diversa da quella dei fluidi extracellulari.
SPAZIO
EXTRACELLULARE
ESTERNO
[Na+] ELEVATO
CITOSOL
INTERNO
[K+] ELEVATO
Poiché gli ioni sono carichi elettricamente, il loro trasporto risulta
nello stabilirsi di un gradiente elettrico attraverso la membrana
plasmatica: il POTENZIALE DI MEMBRANA
La membrana plasmatica contiene: - pompa che trasporta K+ nel citosol
(Na+/K+ ATPasi)
- canali a perdita di K+ (leak channels)
SPAZIO
EXTRACELLULA
RE ESTERNO
K+
EFFETTO CONCENTRAZIONE
+++++++++++++
+
+++++++++++++
+
-------------CITOSOL
-INTERNO
--------------
K-+-
K+
K+
K+
K+
K+
EQUILIBRIO: –60mV
(potenziale a riposo)
EFFETTO CARICA
K+ tende ad uscire per GRADIENTE DI CONCENTRAZIONE
eccesso di carica all’esterno
K+ tende a rientrare per GRADIENTE ELETTROCHIMICO
Le cellule nervose e muscolari sono eccitabili, cioè il loro potenziale
di membrana può variare in risposta a diversi stimoli
TRASMISSIONE DI UN SEGNALE
Questà propietà è dovuta alla presenza, nella membrana plasmatica,
di canali del Na+ e del K+ a controllo di potenziale. Quando questi
canali si aprono, gli ioni si muovono secondo il loro gradiente di
concentrazione, portando ad un’alterazione del potenziale a riposo
che assume un andamento caratteristico definito POTENZIALE
D’AZIONE.
canali voltaggio dipendenti per il Na+
2 tipi di canali K+
sempre
aperti
voltaggio
dipendenti
mantengono potenziale
a –60mV
apertura dopo
depolarizzazione
POTENZIALE D’AZIONE
viene scatenato da uno stimolo che depolarizza la membrana
plasmatica di un valore sufficiente a superare il livello soglia di
apertura dei canali voltaggio-dipendenti per il Na+
direzione di propagazione
----- +++++
+++++ ----assone
La regione negativa alla superficie della
membrana attira gli ioni positivi dalle
regioni adiacenti a riposo, che hanno
carica positiva.
+++++ --------- +++++
segmento
attivato
Allo stesso modo, gli ioni positivi all’interno della membrana in
corrispondenza della regione attivata scorrono sotto la membrana
in direzione delle regioni a riposo a carica negativa dell’assone.
L’effetto netto di questi flussi riduce la differenza di potenziale
attraverso la membrana nelle regioni adiacenti, facendo sì che
queste regioni superino la soglia di generazione di un potenziale
L’arrivo di un impulso nervoso alla fine di un neurone provoca la
fusione di vescicole sinaptiche con la membrana plasmatica,
consentendo il rilascio di neurotrasmettitore dalla cellula
presinaptica nello spazio sinaptico. Il neurotrasmettitore si lega al
recettore a controllo di ligando sulla membrana plasmatica della
cellula bersaglio.
TRASPORTO ATTIVO
Pompe ATPasiche
Contro gradiente di concentrazione
Richiede ENERGIA
normalmente ATP
Le proteine del trasporto attivo sono:
-saturabili
-specifiche
trasporto attivo DIPENDENZA DIRETTA:
-usa ATP
-serve per tutti gli ioni positivi
-crea un gradiente di concentrazione
trasporto attivo DIPENDENZA INDIRETTA:
-usa il gradiente di concentrazione formatosi nella dipendenza diretta
-veicola sostanze organiche (zuccheri e aminoacidi)
-altri ioni positivi e/o negativi
POMPE DI TIPO P a trasporto attivo diretto
Legano temporaneamente, durante il ciclo, un gruppo fosforico
sottratto dall’ATP.
1) Pompa H+ ATPasi
2) Pompa Ca2+ ATPasi
3) Pompa Na+/K+ ATPasi
x H+
x Ca2+ fuori dalla cellula o dentro il
RE
Na+ verso esterno
K+ verso interno
Proteine transmembranarie in cui un unico polipeptide svolge tutte le
funzioni della pompa:
-idrolisi ATP
-legame del fosfato
-trasporto dello ione
1) H+ ATPasi
-
nelle membrane plasmatiche di batteri, lieviti e piante
trasportano H+ fuori dalla cellula.
nelle cellule che delimitano lo stomaco pompano H+ all’esterno
generando acidità dei succhi gastrici
trasporto di H+ nei lisosomi e negli endosomi
sintesi di ATP in cloroplasti e mitocondri (ATP-sintasi)
2) Ca2+ ATPasi
-
mantiene basse le [Ca2+ ] nella cellula
pompa Ca2+ fuori dalla cellula, nel RE e/o nel
mitocondrio
Ca2+ ESTERNO
INTERNO
2.5-5 mM
0.0001 mM
3) Na+/K+ ATPasi
-
3 Na+ all’interno
2 K+ all’inerno
-
Genera il potenziale d’azione di cellule nervose e muscolari
Consuma circa il 25% dell’ATP prodotto dalla cellula
Svolge un ruolo critico nel mantenimento dell’EQUILIBRIO OSMOTICO
e del VOLUME CELLULARE
Citoplasma è ricco di macromolecole
ACCUMULA CARICA POSITIVA
ALL’ESTERNO
ZUCCHERI
AMMINOACICI
NUCLEOTIDI
RICHIAMO DI H2O
x OSMOSI
3 Na+
2K+
MANTENGONO EQUILIBRIO OSMOTICO
e PREVENGONO AUMENTO DEL VOLUME
CELLULARE X RICHIAMO DI H2O
MECCANISMO D’AZIONE DELLA Na/K ATPasi
TRASPORTATORI ABC (trasporto attivo a dipendenza diretta)
ABC
ATP Binding Cassette: dominio che lega ATP conservato
in questa famiglia di
trasportatori
trasportano: IONI
ZUCCHERI
AMINOACIDI
FIBROSI CISTICA
Alterazione di un gene che codifica x un
trasportatore ABC.
Il prodotto di questo gene (regolatore della
conduttanza transmembrana della fibrosi cistica,
o CFTR) funziona come canale del Cl- nelle cellule
epiteliali
trasporto attivo DIPENDENZA INDIRETTA
Es. trasporto del GLUCOSIO “2-Na+/1-glucosio”
MECCANISMO D’AZIONE DEL TRASPORTATORE
La transizione conformazionale è resa possibile grazie
all’energia liberata dal passaggio del Na+ che si sposta
secondo il gradiente di concentrazione
GLUCOSIO PERMEA
sia x
DIFFUSIONE FACILITATA
(secondo gradiente)
TRASPORTO ATTIVO
(contro gradiente)
ALTRI TIPI DI TRASPORTO
FAGOCITOSI:
ingestione di grosse
particelle (batteri)
PINOCITOSI
o
ENDOCITOSI
MEDIATA
DA
RECETTORE:
assorbimento di fluidi o di
macromolecole in piccole
vescicole
ENDOCITOSI:
ESOCITOSI
FAGOCITOSI:
- amebe: catturare cibo
- organismi pluricellulari: meccanismo di difesa contro
microrganismi ed eliminazione di cellule vecchie o danneggiate.
(LEUCOCITI, MACROFAGI e NEUTROFILI)
PINOCITOSI o ENDOCITOSI MEDIATA DA RECETTORE:
Le macromolecole che devono essere internalizzate si legano a specifici
recettori sulla membrana cellulare. Questi recettori sono concentrati in
regioni specifiche della membrana plasmatica, le FOSSETTE
RIVESTITE DI CLATRINA
ESOCITOSI