La regolazione dell’espressione genica
Tutte le cellule di un organismo multicellulare possiedono tutti i geni della propria
specie; tuttavia solo una piccolissima frazione è attiva in ogni cellula; per esempio, in
una cellula delle isole del pancreas è attivo il gene per l’insulina, non quelli per
l’emoglobina (attivi nelle cellule dei midollo osseo, progenitrici dei globuli rossi) o per la
pepsina (attivi nelle cellule della mucosa gastrica)
Durante lo sviluppo embrionale di un organismo multicellulare, le cellule destinate a
costituire i diversi tessuti e organi attivano in momenti precisi i geni che codificano le
proteine che stimolano o inibiscono la proliferazione cellulare e quelle caratteristiche del
tessuto in formazione (per esempio osseina per le ossa, cheratina per la pelle)
Anche gli organismi unicellulari hanno bisogno di attivare alcuni geni solo in particolari
circostanze (per esempio la presenza di un nutrimento che richiede particolari enzimi)
per non sprecare l’energia e le molecole necessarie
Le cellule tumorali hanno perduto, anche in seguito a mutazioni somatiche, la capacità
di regolare l’espressione genica relativa alle proteine che controllano la proliferazione
cellulare
Dunque nelle prossimità dei geni ci devono essere recettori in grado di percepire segnali
provenienti dall’ambiente esterno e veicolati all’interno della cellula; tali recettori
devono quindi segnalare ai geni se trascrivere o no, in funzione dei segnali provenienti
dall’esterno
Il sistema lac in E. coli
HOC6’H2
C5’
OH
O
O
H
C4’
H
Lattosio
OH
H
C3’
C2’
H
OH
C1’
C4’
H
H
OH
C2’
OH
H
H
H
OH
C5’
O
HOC6’H2
b-galattosidasi
O
C5’
OH
H
C1’
OH
H
C3’
H
Galattosio
C3’
HOC6’H2
C4’
H
OH
C1’
HOC6’H2
C5’
H
H
H
O
L’enzima b-galattosidasi,
che consente di utilizzare
il lattosio come fonte di
carbonio e di energia,
viene sintetizzato in
presenza di lattosio che
quindi oltre che esserne il
substrato è anche
l’induttore della sua
sintesi
OH
H
C4’
OH
+ H2O
C1’
OH
H
C2’
C3’
C2’
OH
H
OH
Glucosio
H
Dal lattosio è indotta
la sintesi di altri 2
enzimi: permeasi e
transacetilasi; anche la
permeasi è coinvolta
nel metabolismo del
lattosio, poiché facilita
l’ingresso del lattosio
nella cellula.
I geni strutturali e il gene I
I
Z
Y
I+ = allele normale:Z, Y, A trascrivono in
presenza dell’induttore;
I- = mutante costitutivo: Z, Y, A trascrivono
sempre;
Is = allele inibente la risposta all’induttore:
Z, Y, A non trascrivono mai
A
trascrizione
Merodiploidi per definire le relazioni funzionali
tra gli alleli
Un unico mRNA policistronico
I+Z-/I-Z+
traduzione
I-Z-/I+Z+
b-galattosidasi
transacetilasi
I-Z-/I-Z+
Trascrive*
sempre
IsZ+/I+Z+
Non trascrive
mai
IsZ+/I-Z+
Non trascrive
mai
F’(lac)
permeasi
Geni lac
1) Is è dominante su I+ che a sua volta è
dominante su I-
Trascrive* con
l’induttore
Trascrive* con
l’induttore
*un mRNA per un enzima funzionale
2) I+ è dominante su I- indipendentemente dalla posizione in cis o in trans rispetto a Z+
Operatore, promotore e operone
I
P
O
Z
Y
A
operone
O+ = allele normale:Z, Y, A trascrivono in presenza dell’induttore;
Oc = mutante costitutivo: Z, Y, A trascrivono sempre
O+Z+/OcZ+
Trascrive* sempre
O+Z+/OcZ-
Trascrive* con
l’induttore
O+Z-/OcZ+
Trascrive* sempre
IsO+Z+/I+OcZ+
Trascrive* sempre
Il prodotto del gene I è una
proteina diffusibile che, in assenza
dell’induttore, si lega ad O e
impedisce la trascrizione di Z, Y ,
A; in presenza dell’induttore non
si lega ad O e consente la
trascrizione di Z, Y, A
*un mRNA per un enzima funzionale
1) Oc è dominante su O+, ma solo in cis rispetto a Z+ (cis-dominanza)
2) Gli alleli normali di P sono cis-dominanti sui mutanti difettivi di P, che
impediscono incondizionatamente la trascrizione di Z, Y, A, impedendo
l’inserimento della RNA polimerasi
Controllo negativo dell’espressione genica
nell’operone lac
RNA polimerasi
I
P
O
Z
Y
A
Y
A
induttore
mRNA
I-
I
Is
Oc
P-
P
O
Z
repressore non legato
all’induttore, che si lega
ad O e impedisce la
trascrizione
repressore legato
all’induttore, che
non si lega ad O e non
impedisce la trascrizione
repressore I- senza affinità
per O, con cui non si lega
mai e non impedisce mai
la trascrizione
I
P
O
Z
Y
A
repressore Is senza affinità
per l’induttore, che si lega
sempre ad O e impedisce
sempre la trascrizione
I
P
O
Z
Y
A
operatore senza affinità
per il repressore che non
lega mai e non impedisce
mai la trascrizione
I
P
O
Z
Y
A
promotore senza affinità
per l’RNA polimerasi che
impedisce sempre la
trascrizione
Controllo positivo dell’espressione genica nell’operone lac
cAMP si lega a
una proteina
attivatrice
(CAP).
P
O
Z
RNA polimerasi
CAP
Y
A basse
concentrazioni di
glucosio aumenta la
concentrazione di
AMP ciclico (cAMP).
A
mRNA
Il complesso CAP-cAMP si lega
al promotore e ne aumenta
l’affinità per l’RNA
polimerasi…
…e, in presenza di lattosio, che
“blocca” il repressore,…
cAMP
…intensifica la trascrizione di
Z, Y ed A.
Controllo negativo dell’espressione genica nell’operone trp
P
O
trpE
trpD
triptofano
RNA polimerasi
trpC
trpB
trpA
repressore non legato al
triptofano, che non si
lega ad O e consente la
trascrizione
mRNA
repressore legato al
triptofano, che si lega ad
O e non consente la
trascrizione
Organizzazione della cromatina e controllo
della trascrizione negli eucarioti
Nei nuclei in interfase i
cromosomi hanno domini
separati, che si possono
vedere con sonde specifiche
Le regioni che
devono
trascrivere si
spostano alla
superficie dei
domini
L’acetilazione degli istoni, mediata dalle
proteine attivatrici e da trans-acetilasi, allenta
il legame con il DNA, che diventa accessibile ai
fattori di trascrizione e alla RNA polimerasi
attivatore
transacetilasi
deacetilasi
HO
La metilazione degli
istoni compatta la
cromatina e rende il
DNA inaccessibile
C H3
metile
C
CH2
acetile
O
Terminata la trascrizione, gli istoni sono
deacetilati con l’azione di una deacetilasi e si
ripristina il legame DNA-istoni nei nucleosomi,
tornando alle condizioni di riposo.
Regolazione della trascrizione negli eucarioti:
promotore e intensificatori
Regioni regolatrici dei geni eucariotici
Regioni
Sequenze
,
TATA box
Funzioni
Posizioni
Legame con fattori di trascrizione,
RNA polimerasi II
25-30 nucleotidi a
monte
Promotore (altre regioni)
CAAT box
e/o GC box
Legame con fattori di trascrizione
(generali e specifici)
Distanze varie, a
monte
Intensificatori
Varie
Legame con proteine attivatrici,
modificazione della cromatina
Varia
Regione centrale del promotore
Attivatori
Complesso di fattori
di trascrizione
Intensificatore
RNA polimerasi II
CAAT
box
TATA
box
Inattivazione a lungo termine della trascrizione negli
eucarioti: eterocromatina costitutiva e facoltativa
Le regioni cromosomiche con la
cromatina compatta sono visibili
nei nuclei in interfase come zolle
più colorate e costituiscono
l’eterocromatina
Il compattamento avviene
con l’avvolgimento dei
nucleosomi a formare il
solenoide…
…e con il ripiegamento
della cromatina sulla
matrice.
Le regioni con DNA satellite non hanno
geni, non trascrivono mai e sono sempre
compatte (eterocromatina “costitutiva”)
Il cromosoma X inattivo rimane condensato
in interfase (eterocromatina “facoltativa”),
ove è visibile come “corpo di Barr”.
HH
H
H N
Durante le fasi precoci dell’embriogenesi uno dei 2 cromosomi X
di femmine di mammifero è inattivato a caso in ciascuna cellula
somatica e l’inattivazione è ereditata dalla discendenza cellulare;
i tessuti somatici femminili sono mosaici clonali “epigenetici”,
cioè d’espressione genica.
C H
H
C C
C H
N
C N
C
O
C
La metilazione
della citosina nel
DNA in sequenze
5’CG3’ è connessa
con l’inattivazione
del cromosoma X
Inattivazione a lungo termine della trascrizione
negli eucarioti: l’imprinting genomico
Durante la gametogenesi sono inattivati
alcuni geni specifici, che sono diversi
nelle cellule germinali maschili e
femminili (“imprinting genomico”).
Disomia
uniparentale
geni inattivati nei gameti maschili
geni inattivati nei gametifemminili
Nelle fasi precoci dello sviluppo embrionale
l’inattivazione viene rimossa nella linea germinale
La metilazione della
citosina nel DNA in
sequenze 5’CG3’ è
connessa con
l’imprinting genomico
La metilazione della citosina nel
DNA in sequenze 5’CG3’ è connessa
infine con l’inattivazione di geni ad
azine tessuto-specifica
Zigote bilanciato
geneticamente (tutti i geni in 2
copie) ed epigeneticamente
(almeno una copia attiva di
tutti i geni).
Zigote bilanciato geneticamente
(tutti i geni in 2 copie) ma
sbilanciato epigeneticamente
(almeno ungene non ha una
copia attiva).
Il controllo dopo la trascrizione negli eucarioti
Controllo quantitativo: l’interferenza da RNA
Complesso RISC
Complesso DICER
siRNA
trascrizione
1) Un breve (70 nucleotidi) RNA a doppio
filamento viene tagliato da DICER
2) Un dei 2 filamenti complementari
residui (21 nucleotidi), detto siRNA,
si lega a RISC
3) siRNA-RISC si appaia a una
regione complementare di uno
specifico mRNA e lo frammenta
mRNA
4) I frammenti dell’mRNA sono
degradati
miRNA
mRNA
1) Un breve (70-130 nucleotidi) RNA a
forcina viene tagliato da DICER
3) Un dei 2 filamenti complementari residui (19-24
nucleotidi), detto miRNA, si appaia a una regione
complementare all’estremità 3’ non tradotta di uno
specifico mRNA e ne blocca la traduzione
Il controllo della traduzione negli eucarioti
b-tubulina
….
….
…..-ArgaanGlu-Ile
-aa5- aa6-Met5’
AUG
AAAAAAAA 3’
In assenza/bassa concentrazione di b-tubulina, la traduzione dell’mRNA della b-tubulina procede fino
al completamento e il rilascio del polipeptide.
b-tubulina
….
….
Met-Arg-Glu-Ile
5’
AUG
AAAAAAAA 3’
RNasi
In presenza di concentrazioni alte di b-tubulina, la traduzione dell’mRNA della b-tubulina è bloccata
dal legame che si crea tra alcuni aminoacidi centrali della b-tubulina già presente nel citoplasma con i
primi 4 aminoacidi della b-tubulina nascente; tale complesso induce l’azione di una Rnasi che taglia e
degrada l’mRNA legato al ribosoma.
Un gene più catene polipeptidiche?
“editing” dell’mRNA
Cambiamenti nella sequenza degli mRNA
possono avvenire per inserzione/delezione di
base (p. es mRNA mitocondrali)
UUUU
mRNA
Cambiamenti nella sequenza degli mRNA possono
avvenire per isostituzione di base (p. es mRNA
dell’apolipoproteina B nell’intestino umano)
Esoni “costitutivi”
mRNA
U AA
C
“splicing” alternativo
pre-mRNA
Introni
Esoni “facoltativi”
mRNA alternativi
Al gene SLO nell’uomo sono attribuiti 500
possibili mRNA nelle cellule della coclea
Al gene Dscam in Drosophila sono attribuiti
più di 30.000 possibili mRNA nei neuroni
Splicing alternativo: determinazione
del sesso in Drosophila
X Y
Sxl
X X
Tra
AUG
UAG
AUG
UGA
SXL
UAG
UGA
pre-mRNA
Polipetide tradotto
TRA
TRA-
Dsx
AUG
UGA
UAG
AUG
UGA
pre-mRNA
Polipetide tradotto
DSX m
Differenziamento maschile
DSX f
Differenziamento femminile
UAG
Ingegneria genetica
Le tecniche dell’ingegneria genetica permettono di isolare un gene che codifica per una determinata
proteina e di inserirlo, mediante dei vettori adatti, nel genoma di un altro organismo. Queste
tecniche sono state applicate al campo della medicina per la produzione farmaci, per la diagnosi di
malattie genetiche, per la progettazione di vaccini. In agricoltura si stanno progettando piante e
animali transgenici, per aumentarne la produttività e migliorarne la qualità.
Ma tutto ciò pone problemi nuovi di carattere etico, economico, biologico che devono essere
attentamente valutati per la tutela della vita e dell’ambiente.
Batterio
Cromosoma
batterico
Plasmide isolato
Gene da isolare
Plasmide
DNA ricombinante
(plasmide)
Inserimento del
plasmide nella
cellula batterica
Copie del gene
isolato
Cellula
contenente il
gene da
isolare
DNA purificato
Batterio
ricombinante
duplicazione
Copie delle proteine come
ad esempio insulina, ormone
della crescita
Enzimi di restrizione
Gli enzimi di restrizione sono endonucleasi capaci di tagliare il DNA producendo
rotture a doppia elica in corrispondenza di sequenze specifiche di nucleotidi (siti di
restrizione); di essi si servono i batteri per distruggere i cromosomi virali
endonucleasi
Sito di restrizione
EcoRI
5’GAATTC
3’CTTAAG
HaeIII
5’CC GG
3’GGCC
Formazione di code
5’CCAA
3’GG
GG
TT CC
Tipo di taglio
Sfalsato, adesivo
Tronco, non adesivo
Un taglio è detto adesivo
quando:
1) È sfalsato, o sono
aggiunti nucleotidi
al 3’;
2) I nucleotidi a singolo
filamento, sfalsati o
aggiunti, sono fra
loro complementari
Le estremità di un taglio adesivo si possono unire
mediante i legami idrogeno tra le basi complementari
presenti a singolo filamento sui 2 segmenti di DNA;
successivamente, mediante l’azione di ligasi e, se serve,
di DNA polimerasi, si saldano i filamenti
polinucleotidici (vedere diapositiva 8)
Si possono aggiungere al 3’ di uno dei 2 filamenti di un capo di un taglio tronco, non
“adesivo”, una breve sequenza, ripetizione di un solo nucleotide e al 3’ dell’altro capo la
sequenza complementare, rendendo adesivo il taglio
I siti di restrizione sono brevi sequenze palindromiche: la sequenza posta a un lato
rispetto al centro del sito è complementare alla sequenza inversa rispetto a quella posta al
lato opposto
Mappe di restrizione
Una mappa di restrizione consiste nell’identificazione della successione dei
siti di restrizione per diverse endonucleasi in un segmento di DNA,
misurando la distanza tra i siti come numero di nucleotidi
1) Si esamina un
frammento di DNA,
marcandolo con 32P al 5’
2) Si digerisce il frammento con
diversi enzimi di restrizione
Marcatura con con 32P
3) Si stima la lunghezza dei
frammenti dallo spazio
percorso su un gel
mediante elettroforesi
Siti di restrizione
dell’enzima
Siti di restrizione
dell’enzima
L’elettroforesi consiste nel collocare le macromolecole in un
pozzetto all’estremità di un gel (gelatina) che viene poi sottoposto
un campo elettrico; le molecole si spostano con una velocità
proporzionale alla propria carica elettrica e inversamente
proporzionale lla propria lunghezza; poiché nel DNA la carica è
distribuita uniformemente, la velocità dipende solo dalla lunghezza
del frammento
Il DNA ricombinante
DNA da clonare
vettore
VETTORI
Sono cromosomi
capaci di replicare e
selezionabili, in cui si
possono inserire
segmenti di DNA di
interesse
Digestione con
endonucleasi
DNA polimerasi I +
ligasi
I vettori possono essere :
-plasmidi,
-cromosomi di fagil,
ibridi fago-plasmide (cosmidi),
tutti capaci di replicarsi in E. coli,
-YAC, cioè cromosomi artificiali di
lievito, capaci di replicarsi in lievito
In ogni tipo di vettore si può
inserire un frammento di DNA di
una particolare lunghezza massima
Estremità adesive spontanee o
costruite, fra loro uguali
Estremità adesive
complementari alle precedenti
La clonazione del DNA
Clonazione non
selettiva
Genoma da saggiare
frammentato con
enzimi di restrizione
P = 1-(1-f)N
Individuazione dei
geni clonati:
Southern blotting
Clonazione
selettiva
Cromosomi di l
frammentati
ligasi
L’ibridazione cn la
sonda radioattiva si
effettua
direttamente sui
frammenti del
genoma,
identificando così il
frammento da
saggiare
Genoteca di DNA
ricombinante
1) Si effettua
l’elettroforesi su gel
dei frammenti a
doppia elica
2) Si denatura il DNA e lo si fa
aderire a singolo filamento,
nelle stesse posizioni, a un filtro
Clonazione
Ibridazione su filtro
con sonda radioattiva
complementare al
DNA da studiare
3) Si ibridizza con sonda
radioattiva complementare al
frammento cercato e si effettua
l’autoradiografia, localizzando
così il gene studiato
C-DNA, walking cromosomico e sequenziamento
cDNA
WALKING CROMOSOMICO
RNA
Mediante piccole sonde, in comune a più
frammenti è possibile mettere questi
ultimi in sequenza
Trascrittasi
DNA
polimerasi
inversa
AAAAA
TTT
primer
IL cDNA CONSENTE DI
STUDIARE GLI mRNA E
CONFRONTARLI CON I
GENI O I TRASCRITTI
PRIMARI
SEQUENZIAMENTO DEI NUCLEOTIDI
1) Si marca con 32P l’estremità 5’ del
frammento da analizzare, quindi si
denatura il DNA
2) Si distruggono selettivamente basi o
loro combinazioni (G, G+A, C, C+T), che
rendono più fragile il filamento saggiato,
che quindi si rompe in quei punti
3) Si stima la lunghezza dei frammenti
dallo spazio percorsoin una corsa
elettroforetica
G
G+A
C
C+T
G
C
A
T
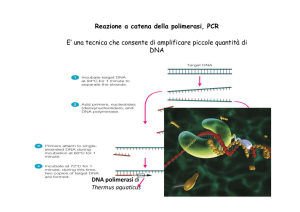
Reazione a catena della polimerasi (PCR)
PCR
Taq polimerasi, resistente al caldo
Primer 1
Regione complementare a
Primer 2
Regione complementare a
Si alternano cicli di denaturazione (a temperatura alta) e di
replicazione ( a temperatura bassa), ciascuno dei quali
porta un raddoppiamento del numero delle molecole
Costruire nuovi geni
SINTESI DI SEQUENZE NON SU STAMPO:
è possibile allungare filamenti di DNA al 5’ aggiungendo nucleotide
dopo nucleotide costruendo oligonucleotidi di sequenza voluta
INDUZIONE DI MUTAZIONI SPECIFICHE
1) Si rompe il DNA
con endonucleasi
2) Si digeriscono singoli
filamenti in direzione 5’
con esonucleasi
3) Si modificano specifici nucleotidi sul filamento
rimasto; dopo la replicazione, anche il nuovo
filamento sarà mutato nello stesso sito
Progressi nel sequenziamento
Sequenziamento del DNA di singoli cromosomi
Isolamento di singoli
cromosomi con citometria
a flusso
3) Singoli cromosomi contenuti
in microgocce sono riconosciuti
da sensori laser…
2) Colorazione con 2 fluorocromi,
rispettivamente specifici per A-T e G-C,
la cui emissione risultante è specifica per
ciascun cromosoma
1) Raccolta di
cromosomi in
metafase con
shock osmotico
4) Si effettua
l’elettroforesi dei
nuovi filamenti…
Sequenziamento automatico del DNA
primer
1) Si dispone di singoli
filamenti di DNA con
primer e DNA polimerasi
AGGTTAC
3’
G
TCCAAT
T
TCCAA
2) Si forniscono miscele di
nucleotidi contenenti di-desossinucleotidi, privi di –OH al C3’,
che bloccano l’allungamento del
nuovo filamento di DNA
4) …ricevono una carica
elettrica specifica…
5) …e sono deviati da un campo
elettrico nella “propria” provetta
ddA
ddG
ddT
ddC
3) Ciascun di-desossinucleotide
è marcato con un fluorocromo
specifico
A
TCCA
TCC
A
TC
C
T
C
T
5’
TCCAATG
5’
3’
5) …e mediante spettroscopia
automatica, si ottiene il
profilo della sequenza
Dalla mappatura al sequenziamento del genoma
Marcatori molecolari
Sequenziamento “clone dopo clone”
Alleli dei siti di
restrizione (RFLP)
1) Mappatura di restrizione (“fingerprinting”)
di una raccolta di grandi cloni da una
genoteca, identificando le sovrapposizioni
Sito di restrizione
Trattamento con
l’enzima di restrizione
Allele non “tagliabile”
Alleli per singole
sostituzioni
nucleotidiche (SNP)
2) Selezione del numero minimo di cloni sovrapposti
3) Frammentazione in sub-cloni dei grandi cloni
superstiti e loro sequenziamento
Alleli per numero di
ripetizioni di mini- e
microsatelliti (SSLP)
Identificati per
sequenziamento
Sequenziamento “shotgun”
1) Frammentazione di genomi in
genoteche di cloni di diversa grandezza
Mappature di marcatori
molecolari fra loro/con geni
Mappe genetiche (ricombinazione)
Mappe cromosomiche: ibridazione in
situ con sonde fluorescenti (FISH)
Mappe fisiche: sequenziamento del genoma
2) Sequenziamento di successivi campioni casuali
di cloni di diversa grandezza e analisi
automatizzata delle sequenze sovrapposte (“prova
ed errore”)
Genomica strutturale
Il sequenziamento del genoma utilizza le metodiche già descritte seguendo diverse fasi
1: compilazione – consiste nella ripetizione per
numerose volte dell’intero processo per riempire
le lacune e correggere gli errori dovuti alla
natura empirica e casuale del processo
Le lacune, oltre agli errori di metodo, sono
dovute a regioni difficilmente sequenziabili:
DNA satellite e geni ripetuti in tandem
2: annotazione – consiste nell’identificazione di
presunti geni, riconoscibili come “open reading
frames” (ORF), nel caso di codificazione di
polipeptidi
Le ORF iniziano con codoni di inizio traduzione
(sul filamento trascritto 3’TAC5’, su quello
complementare 5’ATG3’) e , dopo numerose
triplette, terminano con codoni di stop.
In un tratto di DNA sequenziato per la ricerca di
ORF sono possibilli 3 tentativi di lettura, sfalsati
di un nucleotide, per ciascuno dei due filamenti
complementari.
3: verifica delle ORF – consiste nella ricerca in
prossimità delle ORF di altre sequenze
funzionali (promotori; negli eucarioti siti di
splicing, di poliadenilazione etc.).
Oggi sono disponibili software sofisticati che
minimizzano gli errori, tenendo conto di
caratteristiche note del genoma (p. es.
abbondanza di “isole CpG”- 5’CG3’ in
prossimità di geni)
Solo il 5% del genoma umano consiste nei suoi
circa 26.000 geni; il 10% consiste in DNA
satellite, il 50% in elementi trasponibili
interspersi.
Genomica funzionale e proteomica
Data l’estrema maggiore ricchezza del repertorio proteico
(proteoma) di un organismo rispetto al genoma, è utile
indagare il proteoma nel suo complesso.
Genomica funzionale
1) Una volta identificata una ORF come
possibile gene, per verificare la funzione
del polipeptide codificato si ricorre a
banche dati di sequenze geniche note.
Si effettua un’elettroforesi bidimensionale del proteoma espresso
in determinate condizioni ambientali di una popolazione cellulare.
2) Si effettua un confronto dettagliato
con i geni simili trovati.
3) Si analizzano i ”motivi” funzionali dei
polipeptidi codificati, con l’ausilio di
banche dati di catene polipeptidiche.
Ancora oggi per moltissime ORF dei genomi finora
sequenziati non è stata definita una funzione.
Microarray a DNA
Su una lastra sono
collocati 104 -105
pozzetti, ciascuno con un
oligonucleotide a singolo
filamento di DNA.
Si possono ottenere 102 -104
diverse macchie.
Una volta isolata una
proteina nel proteoma,
prelevandola dal gel, la si
tenta di identificare con
una digestione parziale e il
confronto con banche dati
di catene polipeptidiche e
loro frammenti.
Gli oligonucleotidi di ciascuna riga riguardano lo stesso gene,
con l’allele normale e diverse mutazioni.
Il/i gene/i da saggiare è amplificato per PCR, coniugato con
fluorocromi e collocati sul microarray.
Solo sequenze complementari si legheranno ai pozzetti,
rivelando eventuali mutazioni.
Genetica diretta e genetica inversa
Genetica diretta
Genetica inversa
1) Si inducono mutazioni casuali (con radiazioni,
sostanze chimiche, elementi trasponibili).
1) Si isola e si sequenzia una proteina la cui funzione
deve essere indagata).
2) Si effettua uno screening genetico, attrverso
test specifici, per riconoscere i mutanti.
2) Si determinano le possibili sequenze di cDNA,
costruendo un corredo di oligonucleotidi.
3) Si assegnano i nuovi mutanti ai propri gruppi di
complementazione, identificando il gene mutato.
3) Si saggiano gli oligonucleotidi con i cloni di una
ganoteca, identificando il gene putativo.
4) Si analizza il ruolo dell’eventuale nuovo
gene nella rete di interazioni fra geni
(epistasi, soppressione etc.).
4) Si confronta la sequenza polipeptidica codificata dal
gene putativo con sequenze presenti in banche dati,
cercandone i motivi funzionali.
5) Si mappa il gene, prima costruendo una
mappa genetica, quindi una mappa fisica.
5) Si costriscono mutanti “knock out” (con perdita di
funzione, spesso delezioni intrageniche) o con
sostituzione genica, mediante mutagenesi sito-specifica
e se ne studiano gli effetti fenotipici.
6) Si clona e si sequenzia il gene.
6) Si possono usare gli siRNA per inattivare i geni
bersaglio, al posto dei mutanti knock out
(silenziamento epigenetico