Fisiologia del muscolo
1. Il movimento
2. Il tessuto muscolare
3. Struttura del muscolo scheletrico
1
1. Il movimento
La capacità di muoversi è un
vantaggio evolutivo che diventa
sempre più importante
all’aumentare delle dimensioni
dell’organismo. Essa permette
infatti di spostarsi fra diversi
ambienti alla ricerca del cibo, di
partner sessuali e per colonizzare
nuove aree.
Comporta la capacità di trasformare
l’energia chimica in energia meccanica
a. Il movimento ameboide
b. Il movimento ciliare
c. I movimenti muscolari
2
Il movimento ameboide
Chiamato così perché tipico
delle amebe, ma lo
ritroviamo anche in cellule
di organismi superiori,
come leucociti e macrofagi.
• L’ectoplasma contratto e
rigido, si rilassa nella zona
dove originerà lo
pseudopodio.
• L’endoplasma più fluido
scorrerà verso la zona più
rilassata originando lo
pseudopodio.
ENDOPLASMA
ECTOPLASMA
• L’endoplasma dello
pseudopodio ricostituisce il
citoscheletro, mentre il
citoplasma del corpo cellulare
si sposta verso lo
pseudopodio per la
contrazione di filamenti actomiosinici.
• Così si ottiene lo spostamento
3
Il movimento ciliare
assonema
Ciglia e flagelli hanno
struttura molto simile
anche se il movimento
dei flagelli è più
complesso.
4-5 mm
0.2 mm
Il movimento ciliare può produrre due effetti: se la
cellula è sessile provoca il movimento del liquido
circostante, se è mobile, ne provoca lo
4
spostamento.
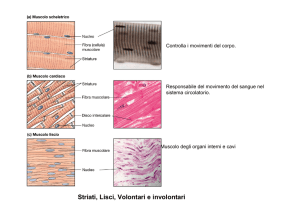
2. Tessuto muscolare
Tessuto muscolare:
il movimento è dovuto ad
un apparato contrattile
costituito da
miofilamenti
Tessuto muscolare liscio:
Evolutivamente più antico.
Nei mammiferi costituisce
la muscolatura degli organi
della vita vegetativa
6
Tessuto muscolare cardiaco:
Tipo particolare di muscolo striato. Le
fibre sono morfologicamente diverse da
quelle della muscolatura scheletrica, ma
presentano striature
7
Tessuto muscolare striato o scheletrico:
Deputato ai movimenti rapidi. Presenta
una caratteristica striatura dovuta alla
disposizione ordinata dei miofilamenti
3. Struttura del muscolo scheletrico
Testo di riferimento:
Silverthorn
9
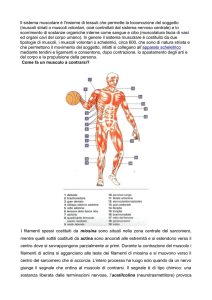
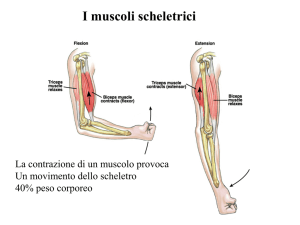
Le fibre muscolari scheletriche
Il muscolo scheletrico è un
insieme di cellule dette fibre
muscolari che lavorano in
maniera integrata.
L’insieme di fibre muscolari
innervate da un motoneurone
costituisce l’unità motoria
Il muscolo intero è contenuto
in una guaina connettivale
che è in continuità con i
tendini
Le singole cellule sono
parallele fra loro secondo il
loro asse longitudinale e
tenute insieme da tessuto
connettivo in cui si trovano
nervi e vasi sanguigni
10
• Le fibre muscolari sono
elementi cellulari giganti (Ø
centinaia di mm; lunghezza
cm) polinucleati, delimitati
dal sarcolemma.
• Il citoplasma prende il nome
di sarcoplasma ed è
premuto verso la periferia
dall’apparato contrattile che
riempie la cellula: le
miofibrille (alcune centinaia
per fibra muscolare).
• Il reticolo sarcoplasmatico
costituisce una rete di tubuli
comunicanti con l’esterno,
fondamentali per il
meccanismo della
contrazione.
11
Ogni fibra muscolare contiene migliaia di miofibrille
Ogni miofibrilla è una struttura contenente diversi tipi di proteine che
si organizzano in un modulo che si ripete uguale lungo tutta la
miofibrilla: il sarcomero.
Il sarcomero è l’unità contrattile del muscolo
12
Il sarcomero è
composto da zone più
scure alternate ad altre
più chiare che vengono
indicate con le lettere
dell’alfabeto:
Dischi Z;
Bande I;
Banda A;
Zona H;
Linea M.
Esse hanno un preciso
significato fisiologico.
13
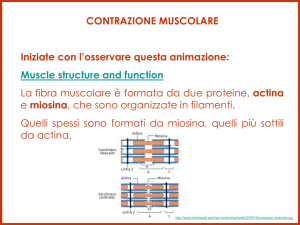
Le proteine che costituiscono il sarcomero sono di diverso tipo:
• Actina e miosina: proteine filamentose contrattili
• Tropomiosina e troponina: proteine regolatorie
• Titina e nebulina: proteine che mantengono in posizione le altre
L’ Actina va a costituire i filamenti sottili
La Miosina va a costituire i filamenti spessi
14
I filamenti sottili
I filamenti sottili sono costituiti da: actina
(p.m.=43000), tropomiosina
(p.m.=70.000) e troponina (p.m.=80000)
Monomeri di G-actina si uniscono a
formare molecole filamentose di Factina.
Ogni filamento sottile comprende 2
molecole di F-actina avvolte a formare
una doppia elica con passo 700Å.
15
L’actina
Ogni molecola di G-actina possiede
un dominio in grado di legarsi con
il filamento spesso a formare ponti
molecolari.
Le molecole di F-actina sono
disposte a lisca di pesce con
orientamento opposto ai due lati
del sarcomero, per potersi legare ai
filamenti spessi che pure sono
orientati in modo opposto ai due
estremi.
16
I filamenti spessi
Sono costituiti da una sola proteina: la miosina (p.m. = 460000)
costituita da una «coda» filamentosa e due «teste»
300-400 molecole di
miosina sono organizzate
in una configurazione
coda-coda in modo da
ottenere due porzioni
simmetriche con la parte
centrale nuda, cioè priva di
teste, centrata sulla linea M
del sarcomero.
In sezione trasversale le
teste, sfasate di 60°,
formano una struttura
elicoidale che si sviluppa
longitudinalmente .
Le teste della miosina
presentano un sito ATPasico e i
siti che le legano l’ actina.
17
Dischi Z: Strutture proteiche che servono per l’attacco dei filamenti sottili
Bande I: parte in cui sono presenti solo i filamenti sottili
Banda A comprende tutta la lunghezza del filamento spesso. Alle estremità
esterne i filamenti spessi si alternano a quelli sottili, al centro ci sono solo
quelli spessi.
Zona H: E’ la zona centrale della banda A occupata solo dai filamenti spessi
Linea M: E’ la zona di attacco dei filamenti spessi
18
Eccitabilità
Il potenziale d’azione del muscolo
scheletrico possiede tutte le
caratteristiche di quello studiato
nella fibra nervosa: per questo
motivo le fibrocellule sono comprese
nei tessuti eccitabili.
La contrazione muscolare
avviene in seguito ad un
evento elettrico. Il neurone
motore, tramite la placca
motrice (giunzione
neuromuscolare), eccita il
muscolo che genera un
potenziale d’azione.
19
Il meccanismo di
contrazione del
sarcomero
Misurazioni a raggi X dimostrarono che
la lunghezza dei miofilamenti resta
costante durante la contrazione, quello
che cambia è il loro grado di
sovrapposizione (teoria dello scorrimento
dei filamenti).
A
I
H
Z
Durante la contrazione, le bande I e la H diminuiscono notevolmente, mentre la A
rimane costante, come ci si aspetta da uno scorrimento dei miofilamenti sottili su
quelli spessi.
20
Lo scivolamento si ha quando le teste
della miosina si attaccano alle
molecole dell’actina adiacente
trascinando il filamento sottile verso il
centro del sarcomero.
Il meccanismo di attacco e distacco
della miosina dall’actina è chiamato
ciclo dei ponti
(1)La testa della
miosina idrolizza
l’ATP ed è pronta al
legame;
1
2
(2) Arriva il Ca2+ e
libera i siti di
aggancio per la
miosina
(3) Movimento del
ponte con idrolisi
dell’ADP;
4
3
(4) L’ATP si lega alla
miosina e ne provoca
il distacco dall’actina
(1)Ricomincia il ciclo.
21
22
A. Sarcomero rilassato:
miosina carica con ADP;
molta affinità per l’actina,
ma siti di legame
mascherati dalla
troponina.
B. Impulso nervoso: liberazione di
Ca2+. Il Ca2+ si lega alla troponina
liberando i siti per il legame con
l’actina.
23
C. La miosina-ADP si lega all’actina
e libera ADP, contraendosi.
D. L’ATP si lega alla miosina-actina
provocando il distacco della
miosina e l’idrolisi dell’ATP e si
torna a miosina-ADP per un nuovo
ciclo.
5 cicli/s
50 nm/s
E. Se termina lo stimolo nervoso,
non c’è più Ca2+ e la troponina
maschera di nuovo i siti di legame
per l’actina e il muscolo torna
rilassato
24
Accoppiamento
eccitazione-contrazione
• Il comando centrale (a) attiva i motoneuroni e la
conduzione del potenziale d’azione (b) fino alla
giunzione neuromuscolare (c).
• Qui il p.d’a. induce il rilascio di acetilcolina dalla
terminazione presinaptica causando un potenziale
del recettore che attiva i canali V-dipendenti del
Na+ e del K+ generando un potenziale d’azione (d).
ACh
25
• Il p.d’a. si propaga lungo la superficie della membrana anche all’interno della cellula
attraverso il sistema dei tubuli trasversi (e), causando il rilascio di Ca2+,
immagazzinato all’interno del reticolo sarcoplasmatico, nel citoplasma (f).
• Il Ca2+ si lega alla troponina C, liberando i siti attivi dell’actina (g) e attivando il ciclo
dei ponti (h).
• Una pompa ATP-dipendente (i) riporta il Ca2+ all’interno del reticolo sarcoplasmatico.
Quando la stimolazione nervosa
termina e con essa il rilascio di
Ca2+ dal reticolo sarcoplasmatico,
il riassorbimento attivo del Ca2+
intracellulare determina il
distacco del calcio dalla troponina
Il ciclo dei ponti si arresta e ha
inizio il fenomeno del
rilasciamento muscolare.
26
27
Relazione tra la lunghezza
del sarcomero e la forza
sviluppata dalla fibra
muscolare
a) Sovrapposizione totale
dei filamenti sottili: non
è possibile l’interazione
fra il filamento sottile ed
il suo corrispondente
spesso
b) Diminuisce la
sovrapposizione fra
filamenti sottili: inizia lo
sviluppo di forza
c) Tutto il filamento spesso
può legarsi al sottile
quindi ancora massima
forza
d) Diminuisce la
sovrapposizione della
parte contrattile quindi
minore forza
e) Filamenti completamente
sfilati non possono
sviluppare forza
28
LA FORZA CHE LA FIBRA MUSCOLARE PUÒ
GENERARE È DIRETTAMENTE PROPORZIONALE AL
NUMERO DI PONTI TRASVERSI CHE SI FORMANO
TRA FILAMENTI SPESSI E FILAMENTI SOTTILI
Se è il sarcomero è troppo lungo,
filamenti spessi e sottili saranno
parzialmente o totalmente sfilati e
formeranno pochi o nessun ponte
trasverso
Se è il sarcomero è troppo corto, i
filamenti sottili dell’emisarcomero
opposto andranno ad interferire
con i ponti che la miosina può
creare con i filamenti del suo
emisarcomero, con minor
sviluppo di forza
29
La scossa muscolare semplice
• Periodo di latenza: tempo che intercorre tra il termine dello
stimolo elettrico e l’inizio della risposta meccanica (circa
10ms)
• Tempo di contrazione tempo necessario a raggiungere il
picco di contrazione
• Fase di rilasciamento: è più lenta nel muscolo affaticato
30
La singola contrazione NON
rappresenta il massimo della forza
che la fibra muscolare può produrre
Aumentando la frequenza dei
potenziali d’azione che dal
neurone arrivano alla fibrocellula,
si può aumentare la sua forza di
contrazione
Il p.d.a. dura 1-3 ms, mentre la
scossa muscolare semplice può
superare i 100 ms. Se i potenziali
sono abbastanza frequenti, la
fibrocellula non avrà tempo di
rilassarsi completamente
In questo caso le contrazioni
potranno sommarsi
31
Sommazione, tetano incompleto,
tetano completo
Il periodo di refrattarietà elettrica della fibra muscolare termina molto prima che essa
si sia rilassata, perciò un secondo p.d.a. induce una contrazione che si somma alla
precedente.
Se la frequenza dei p.d.a è abbastanza alta, le contrazioni successive si fondono
sempre di più fino a dare una contrazione sostenuta nel tempo definita tetano
completo
32
Perché la forza sviluppata durante uno stimolo
tetanizzante è fino a 4 volte più grande di quella
sviluppata dalla scossa muscolare semplice?
1
L’apparato elastico è già in tensione
2
La concentrazione di Ca2+ intracellulare
aumenta ad ogni stimolo perché la sua velocità
di rilascio è maggiore di quella di sequestro
Ca2+
33
Proprietà meccaniche del muscolo
Contrazione isotonica
Quando il muscolo si
muove spostando un
carico, la contrazione
viene definita isotonica
Se il muscolo si flette, parleremo
di contrazione concentrica; se si
estende parleremo di
contrazione eccentrica
34
Contrazione isometrica
Se il muscolo si contrae senza
possibilità di accorciarsi, la
contrazione viene definita isometrica
Entrambe le condizioni
sono fisiologiche
35
Nella contrazione isotonica la forza cresce fin quando non raggiunge il livello del
peso da spostare, poi resta costante per tutta la durata dello spostamento
Nella contrazione isometrica la forza cresce, ma non raggiunge il livello del
peso da spostare
36
Affaticamento muscolare
Se il rifornimento di ATP è più
lento rispetto alla richiesta
del muscolo: la glicolisi si
disaccoppia dal ciclo di Krebs
ed il piruvato è traformato in
lattato che si accumula ed è
tossico per i muscoli.
• Dipende dall’allenamento del muscolo e dall’intensità dello
sforzo.
• E’ un fenomeno reversibile
• Ha funzione protettiva perché informa l’organismo che sta
superando le sue possibiltà
37
In ogni muscolo ci sono tipi diversi di fibre:
1. Fibre di tipo I ossidative a contrazione lenta
Sottili, meno ricche di miofibrille e con r. sarcoplasmatico poco sviluppato. Ricche
di mioglobina (rosse), miosina di tipo «lento». Consumano ATP ad una velocità tale
da poterlo sempre recuperare con la respirazione
Hanno un metabolismo aerobico e
sono resistenti alla fatica
Sono reclutate negli sforzi muscolari
di intensità moderata ma di lunga
durata
Tipo I
Tipo II-B
38
2. Fibre di tipo II-B glicolitiche a contrazione rapida
Maggior diametro, ricche di miofibrille e con r. sarcoplasmatico molto
sviluppato. Poca di mioglobina (bianche), miosina «veloce». L’ATP viene
fornito da PCr e dalla glicolisi.
Intervengono negli sforzi rapidi e
intensi, ma si affaticano rapidamente
3. Fibre ossidative di tipo
II-A a contrazione rapida
Hanno caratteristiche intermedie
rispetto ai primi due tipi di fibre
Piccolo diametro, ricche di mioglobina (rosse). Metabolicamente orientate
sia in senso ossidativo che glicolitico.
39
• I due tipi di fibre a contrazione rapida sono
convertibili con un opportuno allenamento
• Le fibre lente e quelle rapide, invece, non sono
convertibili
• La maggior parte dei muscoli contiene una
miscela dei tre tipi di fibre
• La percentuale di ciascun tipo dipende in gran
parte dal tipo di attività nel quale il muscolo è
specializzato
La percentuale di ciascun
tipo di fibre varia anche
da individuo a individuo,
in quanto ha una base
genetica
maggiore percentuale
di fibre glicolitiche rapide
maggiore percentuale
di fibre ossidative lente
40