Come funziona la pianta (nutrizione idrica)
In una coltura senza suolo in sistema chiuso di pomodoro ciliegino (allevato in vaso in 114 giorni
ciclo primaverile-estivo, 2,6 piante/m2 cimate a 6 palchi) sono stati forniti 130 litri di soluzione
nutritiva. Circa il 10% ha alimentato l’evaporazione dalla superficie del vaso, il 3% era nella pianta
a fine coltura e i rimanenti 113 litri sono stati traspirati dalla pianta.
Per produrre 1 kg di biomassa secca la pianta di pomodoro ha dovuto traspirare 300 litri di acqua
(700 litri per kg di frutto).
Perché la pianta deve farsi attraversare da questo fiume d’acqua che entra dalle radici ed esce dalle
foglie?
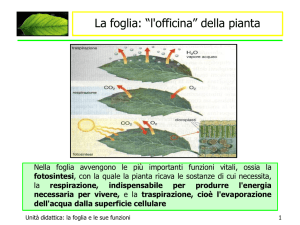
Il compromesso fotosintesi-traspirazione
La fotosintesi è il più importante processo di riduzione biologica che si compie sulla superficie
terrestre, grazie al quale le piante assorbono energia luminosa, la convertono in energia chimica
potenziale e, partendo da composti inorganici semplici (anidride carbonica e acqua), formano
sostanze organiche (glucidi ecc.) le quali costituiscono i composti della materia vivente, sia vegetale
sia animale.
Reazione globale: CO2 + 2 H2O + hν = CH2O + H2O + O2
dove CH2O rappresenta uno zucchero e hν un fotone. Tale reazione è fortemente endotermica.
L’ossigeno che si sviluppa nella fotosintesi proviene dalla scissione fotolitica di molecole di acqua e
non, come si supponeva un tempo, dalla CO2, la quale viene incorporata come tale in un composto
fosforilato a 5 atomi di carbonio, ribulosio-1,5-difosfato (Ru-d-P), che è presente nel cloroplasto ed
è continuamente rigenerato.
La CO2 è presente nell’atmosfera a bassa concentrazione ed entra nelle foglie attraverso gli stomi e
nelle cellule attraverso l’acqua. L’apertura degli stomi consente l’entrata della CO2 ma comporta la
perdita di acqua.
La pianta deve riuscire ad assorbire quanta CO2 è possibile da un’atmosfera dove è presente a bassa
concentrazione (0,03 % v/v) senza perdere tanta acqua prelevata con le radici che serve a tenere
turgide le cellule e permettere l’organicazione del carbonio atmosferico.
Il paradosso dei pori
L’evaporazione da una superficie libera può essere ridotta in proporzione alla copertura della stessa
con un film impermeabile. Ma cosa succede se si copre la superficie per il 99%?
Se i pori sono distribuiti come gli stomi e sono delle stesse dimensioni l’evaporazione viene ridotta
solo circa della metà rispetto alla superficie libera (mentre quando sono chiusi …. il blocco è
totale).
L’evaporazione è un processo diffusione e pertanto fortemente influenzato dal gradiente di
potenziale o di pressione di vapore tra l’aria presente dentro la foglia e quella atmosferica.
Le molecole di acqua evaporanti da una superficie libera salgono in colonna una dietro l’altra e
quindi in parte si ostacolano tra loro mentre quelle che escono da un poro si possono diffondere in
tutte le direzioni all’interno di una semisfera.
Come funzionano gli stomi
Gli stomi regolano gli scambi gassosi della pianta attraverso cambiamenti di potenziale osmotico
(flussi di K+) tra le cellule di guardia e le cellule accessorie. Ma qual è la causa di tali cambiamenti
di potenziale osmotico?
1. Basse concentrazioni di CO2 causano l’apertura degli stomi. Pertanto si aprono al sorgere
del sole quando la fotosintesi consuma la CO2 delle cellule parenchimatiche (palizzata) e del
mesofillo fogliare e si chiudono al buio. In questo modo di notte le piante non perdono
acqua.
2. Basso potenziale idrico della foglia chiude gli stomi anche se la CO2 è bassa e c’è tanta luce.
3. Alte temperature (30-35°C) chiudono gli stomi per stress idrico e/o per l’aumento di
respirazione e quindi di CO2.
4. Ventosità bassa può aumentare la concentrazione di CO2 e farli aprire ma se è elevata può
aumentare la traspirazione e abbassare il potenziale idrico.
5. Gli stomi non si chiudono per effetto diretto dello stress idrico ma in risposta all’ABA,
ormone messaggero. Questo meccanismo spiega il ritardo di 10-15 minuti della risposta
della pianta.
6. La luce blù è più efficace della rossa per l’apertura degli stomi soprattutto a bassa intensità
luminosa ed indipendentemente dall’effetto di CO2 e di ABA.
Il compromesso fotosintesi-traspirazione è stato risolto dalle piante attraverso due controlli a
feedback. Uno fornisce ‘cibo’ alla pianta ed è guidato dalla concentrazione di CO2. L’altro protegge
la pianta dalla ‘sete’ ed è guidato dalla concentrazione di ABA. Inoltre questi due controlli
interagiscono per cui il grado di risposta degli stomi all’ABA dipende dalla concentrazione di CO2 e
la risposta alla CO2 dipende dalla concentrazione di ABA.
Qual è il vantaggio selettivo della traspirazione? Mostra foto DSCN5499.JPG
Le piante possono vivere senza traspirazione? Pensate alle piante chiuse in un terrario.
Quando le piante sono in un ambiente al 100% di umidità relativa la traspirazione è fortemente
ridotta. Come arriva l’acqua alle foglie?
Pressione radicale e guttazione (gocce di acqua liquida sui bordi delle foglie nelle mattinate con
cielo nuvoloso).
La pressione radicale consente una corretta alimentazione idrica delle foglie anche in assenza di
traspirazione. Ma la nutrizione minerale può avvenire senza traspirazione?
Normalmente gli elementi minerali assorbiti dalle radici arrivano alle foglie attraverso il flusso
generato dalla traspirazione. In assenza di questa, la pianta riesce comunque ad avere una
circolazione tra tessuti floematici e xilematici ma si potrebbe avere carenza di Ca e B, elementi
‘poco mobili’ il cui trasporto richiede un attivo flusso traspirativo.
Per le piante allora possiamo parlare che c’è una turgidità ottimale che si ha quando è consentito
almeno un piccolo stress idrico che aziona la traspirazione.
Normalmente in natura e in agricoltura le piante affrontano il problema della carenza di acqua e la
traspirazione è un efficientissimo sistema di raffrescamento (a 30°C l’evaporazione di un grammo
di acqua consuma 580 calorie).
Una foglia esposta alla radiazione solare riflette, trasmette e assorbe parte dell’energia. Assorbe
circa il 50% dell’energia radiante (visibile e infrarosso corto) e di questa energia ne emette circa la
metà come infrarosso lungo secondo la legge di Stefan-Boltzmann.
La foglia può dissipare calore anche per conduzione-convezione ma è soprattutto la traspirazione
che consente di mantenere una temperatura anche più bassa di quella esterna in una giornata
soleggiata. Dal punto di vista fisico convenzione e traspirazione rispondono alla legge generale del
trasporto (analoga alla legge di Ohm), il flusso evaporativo è direttamente proporzionale alla forza
trainante (gradiente di pressione di vapore tra foglia e atmosfera) e inversamente proporzionale alla
resistenza (nel caso della traspirazione entrano gli stomi).
F12= Ψ2-Ψ1/Resistenza
Il gradiente di pressione di vapore dipende soprattutto dalla temperatura e dall’umidità relativa
(UR). L’acqua nella foglia ha un potenziale di -5 ~ -30 bar in equilibrio con un’atmosfera al 98% di
umidità relativa. L’aumento di 10°C della temperatura raddoppia la pressione di vapore. Per es. aria
a 10°C con 100% di UR contiene la stessa quantità di acqua dell’aria a 21°C e 50% di UR e
dell’aria a 33°C e 25% di UR.
Per stimare il bilancio idrico delle piante è quindi fondamentale conoscere T e UR dell’aria.
L’acqua nel continuum suolo-pianta-atmosfera
aria = -400 ~ -500 bar
foglia = -10 ~ -20 bar
suolo = -0,3 ~ -15 bar
Sistema suolo:
tot = m + g + e
Sistema suolo-pianta:
tot = m + g + e + o
tot
potenziale totale è la quantità di lavoro che deve essere svolta per unità di acqua pura per trasferire
reversibilmente e isotermicamente una quantità infinitesima di acqua pura da un deposito di
riferimento (altezza caratteristica, a pressione atmosferica e a temperatura ambiente) all’acqua del
suolo. L’acqua si muove da un punto con Ψ più alto (meno negativo: es. –20) ad uno con Ψ più
basso (più negativo: es. –100).
m
potenziale matriciale è il lavoro da compiere per vincere la forza con cui l’acqua è trattenuta dalla
ritenzione interstiziale dei capillari e all’adsorbimento igroscopico dei colloidi del suolo.
g
potenziale gravitazionale è il lavoro da compiere per vincere la forza peso dell’acqua nel suolo. Se
il terreno è sommerso di acqua è una pressione (valori positivi) se il terreno è drenato è una tensione
(valori negativi).
e
potenziale di envelope (inviluppo) è il lavoro da compiere per vincere la forza peso della matrice
(componente solida del suolo). Aumenta con la profondità.
o
potenziale osmotico, pressione negativa (suzione) cui è sottoposta l’acqua separata da una soluzione
più concentrata da una membrana semipermeabile. È il lavoro che compie l’acqua pura in
movimento verso la soluzione (potenziale negativo).
Dal ‘punto di vista della pianta’ l’acqua nel suolo dovrebbe essere poco salata (alto o), in
superficie (alto g e e) e poco trattenuta (alto m).
Nella gestione dell’irrigazione è molto utilizzata la scala pF.
pF=log10 Free Energy = log10 m
pF
0,0
1,0
2,0
1,7-2,5
2,7
3,0
4,2
cm acqua, hPa, mbar
0
10
100
50-350
700
1000
16000
stato idrico
saturazione
capacità di campo substrati
fine acqua utile substrati
capacità di campo suoli
limite utilizzo tensiometri
vuoto completo
punto di appassimento
La curva tensione/umidità permette di conoscere la quantità di acqua presente nel suolo ad una certa
tensione. Inoltre la tensione dell’acqua del suolo è informativa anche della porosità del suolo
h(mbar)=3/d(mm)
(per es. una tensione di 104 mbar si ha in pori del diametro di 0,3 µm) e quindi anche della sua
granulometria (tessitura).
Adattamento della pianta alla maggiore domanda evapotraspirativa
F12= Ψ2-Ψ1/Resistenza
Aumento del flusso in ingresso (assorbimento radicale)
aumento del potere assorbente delle radici (abbassamento potenziale radicale,
adattamento veloce)
aumento della superficie assorbente (sviluppo dell’apparato radicale, adattamento
lento)
Riduzione del flusso in uscita (traspirazione)
riduzione della differenza Ψaria – Ψfoglia, abbassa il Ψfoglia attraverso l’aumento della
concentrazione osmotica (adattamento veloce)
aumento della resistenza
con chiusura stomatica (adattamento veloce)
con variazioni morfologiche (peli, cuticola, … adattamento lento)
Come funziona la pianta (nutrizione minerale)
Elementi nutritivi e rapporti
Gli elementi essenziali nelle piante sono 17:
C, O e H sono ottenuti dall’acqua assorbita dalle radici e dall’assimilazione della CO2 atmosferica e
costituiscono circa il 95% della biomassa di una pianta.
N, P, K, Ca, Mg e S, sono considerati elementi macronutritivi perché necessitano in maggiore
quantità degli elementi micronutritivi Cl, Fe, B, Cu, Mn, Zn, Mo e Ni (scoperto nel 1987) e
vengono tutti assorbiti essenzialmente attraverso l’apparato radicale.
Un elemento è considerato essenziale se soddisfa queste condizioni (Arnon e Stout, 1939)?
1. la pianta in sua assenza non può completare il ciclo vitale (da seme a seme);
2. si conosce la sua funzione nella pianta e questo ruolo è essenziale;
3. la funzione dell’elemento non può essere sostituita da un altro elemento minerale.
Altri elementi sono quelli benefici, capaci di stimolare la crescita pur non essendo essenziali o sono
essenziali solo per alcune specie. Per es. Na e specie alofite, Si e graminacee, Co e leguminose, Se e
crucifere, Al e barbabietole, mais e legumi tropicali.
Gli elementi essenziali sono presenti nelle piante secondo rapporti variabili tra le specie e frutto
dell’evoluzione su suoli e climi diversi (per es. terreni calcarei e acidi).
Gli elementi nutritivi essenziali (macro e micro) vanno forniti alle piante secondo determinati
rapporti e in quantità sufficienti a soddisfare il tasso di crescita.
Cosa è la crescita?
Un organismo partendo dalla cellula zigote cresce in peso, numero di cellule, quantità di
protoplasma e in complessità.
Una curva sigmoide idealizzata è spesso utilizzata per descrivere la crescita cumulata (per es.
altezza della pianta di pomodoro in cm durante la coltivazione) e una curva a campana (ottenuta
dalla derivata prima di questa) per descrivere la crescita relativa (crescita giornaliera in cm al
giorno).
Si distinguono tre fasi di crescita:
1. fase logaritmica, tasso di crescita crescente (come l’interesse composto);
2. fase lineare, tasso di crescita costante;
3. fase di senescenza, tasso di crescita decrescente.
alt - cum ( cm)
al t -cum (cm)
16 0
400
14 0
350
12 0
300
10 0
250
80
200
60
150
40
100
20
50
0
0
1
6
11
16
21
26
31
36
41
46
51
56
61
66
71
76
81
86
91
0
96
20
40
60
80
100
120
alt * gg
alt * gg
5
4
4,5
3,5
4
3
3,5
2,5
3
2
2,5
2
1, 5
1, 5
1
1
0,5
0,5
0
0
1
6
11
16
21
26
31 36
41 46
51
56
61
66
71
76
81 86
91 96
Pomodoro a ciclo breve (pianta cimata)
0
20
40
60
80
10 0
12 0
Pomodoro a ciclo lungo (pianta non cimata)
Nelle successive lezioni vedremo come fornire alle piante in modo ottimale acqua ed elementi
minerali tenendo conto dell’evapotraspirazione (nutrizione idrica) e del tasso di crescita (nutrizione
minerale).
Gli elementi essenziali vanno forniti in rapporto equilibrato tra loro e con un flusso di fornitura che
assecondi dinamicamente il tasso di crescita.
In genere per decidere se un terreno deve essere concimato con un certo elemento si misura la
concentrazione presente. Ma questo dato non è sufficientemente informativo riguardo alla domanda
della pianta perché questa si estrinseca sempre in termini di flusso (tasso di assorbimento
giornaliero). È quindi necessario conoscere oltre alla concentrazione anche il volume di soluzione
presente o fornita periodicamente attraverso la fertirrigazione.
Ingestad 1982
Con il suo lavoro ha chiarito definitivamente che l’assorbimento minerale è determinato dalla
dinamica della crescita e non dalla concentrazione esterna di un certo elemento nutritivo presente
nel suolo.
In esperimenti con concentrazioni diverse tenute costanti mediante l’aggiunta continua di soluzione
nutritiva fresca il tasso di crescita relativo era indipendente dalla concentrazione esterna e
linearmente correlato col tasso di aggiunta relativo e con la concentrazione interna alla pianta.
A=FCest
L’aggiunta A di nutriente nell’unità di tempo (µmol/min) è assicurata da un flusso F di soluzione
(dm3/min) a una certa concentrazione Cest (mmol/m3).
La variabile guida è quindi il tasso di crescita che deve essere soddisfatto attraverso un flusso di
soluzione a una certa concentrazione.
Negli esperimenti classici è stato usato un volume di soluzione fisso ad elevata concentrazione
esterna ed è stata ottenuta la legge degli incrementi decrescenti (Mitscherlich, 1954) perché con
l’impoverimento della soluzione esterna, a causa dell’assorbimento da parte della pianta, diminuiva
la concentrazione interna alla pianta e quindi diminuiva la crescita e si manifestavano sintomi di
carenza.
Ma nelle piante in natura avete mai visto carenze? foto DSCN2163.JPG
Rg/ gg e Ra/ gg
Rg/ gg e Cin t
0,3
0,3
0,25
0,25
0,2
0,2
0 , 15
0 , 15
0,1
0,1
0,05
0,05
0
0
0
0,05
0,1
0 , 15
0,2
0,25
0,3
Relazione tra Rgr/gg e Rar/gg
0,35
0
0,2
0,4
0,6
0,8
1
1, 2
Relazione tra Rgr/gg e Cint
In natura la crescita delle piante è commisurata al tasso di aggiunta relativo, cioè cresce per quanto
l’ambiente le permette. In agricoltura spesso ad inizio coltura si promette alla pianta di nutrirla per
ottenere la crescita massima ma quando la promessa non viene mantenuta diminuisce la
concentrazione interna e si manifestano sintomi di carenza. Ovviamente tali sintomi spariscono
appena la pianta adegua il tasso di crescita alle risorse nutrizionali disponibili localmente.
Qual è il tasso di aggiunta ottimale?
È quello che consente di ottenere il massimo tasso di crescita. Un ulteriore aumento del tasso di
aggiunta non si traduce in crescita ma in un aumento della concentrazione esterna (accumulo di sali)
e successivamente in una diminuzione della crescita o può anche avere effetti letali.
Quanto detto è vero per la fase di crescita esponenziale quando nessun fattore è limitante e la
crescita dipende solo dalla nutrizione. Poi arriva l’auto-ombreggiamento e altri fattori diventano
limitanti e interferiscono e complicano l’analisi della crescita.
Tornando al pomodoro a ciclo lungo in sistema chiuso, nella lunga fase lineare il tasso di aggiunta
deve soddisfare la crescita vegetativa di circa 30 cm a settimana e la crescita riproduttiva di circa un
palco a settimana.
Il flusso di soluzione da assicurare ogni giorno sarà pari a?
alla domanda evapotraspirativa.
La concentrazione della soluzione nutritiva dovrà essere?
quella necessaria a fornire gli elementi nutritivi richiesti dalla crescita vegetativa e riproduttiva
all’interno di un volume di soluzione definito dalla domanda evapotraspirativa.
In un bosco la crescita dipenderà dalla densità del flusso nutritivo cioè dalla quantità di nutrienti
disponibili per unità di suolo e di tempo. Questa densità è funzione del tasso di mineralizzazione
della sostanza organica del suolo.
In una foresta di conifere in Svezia è stata aumentata la densità del flusso nutritivo secondo una
curva a S. Mediante fertirrigazione sono stati assecondati i cambiamenti stagionali nell’attività
dell’ecosistema causate dal clima e sono riusciti ad aumentare il tasso di assorbimento della
vegetazione. Circa il 40% della azoto fornito per fertirrigazione è stato ritrovato nella lettiera e
nell’humus incrementando notevolmente la densità del flusso nutritivo naturale (mineralizzazione).
Modalità di movimento degli elementi nutritivi verso le radici:
Sviluppo dell’apparato radicale;
Flusso di massa;
Diffusione.
Elementi mobili e poco mobili nel suolo e nelle piante.