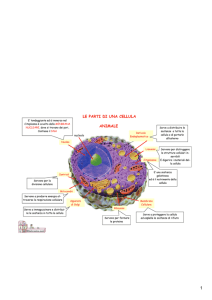
Componenti citoplasmatici
Guardando il citoplasma al microscopio sono visibili numerose membrane e vescicole delimitate da
membrana. Una sorta di organelli distinti o compartimenti con funzioni specializzate.
Nel citoplasma, i materiali sono trasportati da un organello all'altro racchiusi in vescicole di trasporto
che si formano per gemmazione dalla membrana di un compartimento e si fondono per fusione con la
membrana del compartimento di destinazione. Queste vescicole viaggiano da un organello a un altro
ancorate ai microtubuli del citoscheletro.
Quindi dentro alla cellula ci sono
vari flussi di trasporto di sostanze.
Il flusso principale avviene dal reticolo endoplasmatico (nel quale sono sintetizzati i materiali) al
complesso del Golgi (nel quale sono modificati) per essere destinati alla membrana plasmatica, ai
lisosomi, agli endosomi, alle vescicole escretorie, ai vacuoli. Questa via biosintetica viene definita
secretoria perchè molte sostanze sono riversate all'esterno della cellula.
Le vescicole di
trasporto si muovono
verso direzioni precise
quindi durante la loro
formazione viene
impresso loro anche
un indirizzamento, una
sorta di bollo con
l'indirizzo del
destinatario da
raggiungere.
Anche le proteine
devono essere
caricate nella vescicola
adatta a poter
raggiungere il
compartimento
corretto.
Proteine di trasporto che
viaggiano lungo un
microtubulo
Il reticolo endoplasmatico è costituito da
un sistema di membrane che delimita
uno spazio interno chiamato spazio
luminale. Esistono due tipi di reticoli
endoplasmatici: liscio e rugoso. Il reticolo
endoplasmatico rugoso si distingue per la
presenza di ribosomi legati alla superficie
esterna. Entrambi i tipi di reticoli sono
presenti nelle cellule ma in proporzioni
diverse a seconda del tipo di cellule. Ad
es., cellule che devono secernere grandi
quantità di proteine hanno
una maggior proporzione
di reticolo endoplasmatico
rugoso.
Il reticolo endoplasmatico rugoso è il compartimento
dal quale vengono sintetizzati proteine, carboidrati e
fosfolipidi. Le proteine che si originano sono destinate
ad essere escrete o a finire nella membrana come
proteine integrali di membrana o ad andare
all'interno di compartimenti del sistema delle
endomembrane.
Invece i ribosomi liberi sintetizzano proteine che
rimangono nella cellula.
Il reticolo endoplasmatico liscio è coinvolto nella sintesi di lipidi, nella detossificazione di sostanze
dannose per l’organismo, nell’accumulo di ioni Ca. E’ presente in gran quantità nelle cellule endocrine
dove è deputato a produrre ormoni steroidei; nelle cellule del fegato dove è implicato nella
detossificazione di numerosi composti.
Ricordiamo che il citocromo P450 è uno degli enzimi di detossificazione che ossida composti in modo
non specifico i composti idrofobici e li rende più idrofilici e quindi più facilmente smaltibili
dall’organismo. Questo enzima è implicato anche nel metabolizzare i farmaci e polimorfismi di questo
gene producono varianti proteiche che possono avere diversi livelli di attività enzimatica. Tali differenti
livelli sono la causa della diversa efficacia ed effetti collaterali dei farmaci da individuo a individuo.
Alcuni farmaci possono inibire o indurre il citocromo P450. Questo meccanismo è uno dei responsabili
delle interazioni tra farmaci.
Ad es. alcuni antidepressivi inibiscono forme del citocromo P450 causando alte concentrazioni sieriche
(tossicità) di farmaci somministrati in contemporanea.
Es. Il ritonavir (usato contro l’HIV) inibisce alcuni enzimi del citocromo P-450 epatico e può aumentare le
concentrazioni sieriche dei farmaci metabolizzati da questi enzimi (es. gli antiaritmici, l'astemizolo (un
antistaminico), la maggior parte delle benzodiazepine, la cisapride (promuove motilità intestinale)).
Anche il succo di pompelmo, cannella, pepe bianco e nero, zenzero, curcumina, noce moscata, chiodi di
garofano inibiscono forme del citocromo P450.
Altri farmaci possono indurre forme
del citocromo P450 rendendo
inefficace l’altro farmaco
somministrato in contemporanea.
Hanno questo effetto alcuni
sedativi barbiturici e anche
il fumo di sigaretta.
Come si fa a riconoscere la destinazione di una proteina?
La parte iniziale della sequenza amminoacidica, cioè la prima che viene sintetizzata da un ribosoma
libero, contiene una sequenza di amminoacidi detta peptide segnale che viene riconosciuta e indirizza
dal ribosoma verso il reticolo endoplasmatico o altri compartimenti.
L'estremità del peptide nascente si infila in un canale in modo da entrare nel reticolo endoplasmatico e
finirvi nel lume man mano che viene la sintesi proteica. Alla fine della sintesi il ribosoma si distacca dal
reticolo endoplasmatico. Alla nuova proteina viene rimosso il peptide segnale e le vengono aggiunti
carboidrati.
Proteine chaperon aiutano la nuova proteina ad
assumere la struttura tridimensionale corretta.
Un enzima controlla che il ripiegamento proteico
sia avvenuto correttamente (non ci devono essere
amminoacidi idrofobici esposti all'esterno della
proteina ripiegata), in caso contrario lascia ulteriore
tempo alla proteina di ripiegarsi e, se dopo alcuni
tentativi il ripiegamento non è ancora corretto,
la proteina viene marcata (ubiquitinata), trasferita
nel citosol e distrutta.
Qui, gli oligosaccaridi sono rimossi e
la proteina viene distrutta nei proteasomi.
Il complesso di Golgi appare come un insieme di cisterne appiattite e con bordi dilatati. Queste cisterne
sono impilate in ammassi con al massimo 8 cisterne. Nel citoplasma ci possono essere molte di queste
pile interconnesse da tubuli membranosi.
Nella parte del Golgi che
è più vicina al reticolo
endoplasmatico avviene
la distinzione tra
proteine da rimandare al
reticolo endoplasmatico
e quelle che possono
continuare verso la
successiva cisterna del
Golgi. La parte del Golgi
più lontana al reticolo
endoplasmatico
racchiude le proteine in
diversi tipi di vescicole
dirette verso la
membrana plasmatica o
verso altre destinazioni.
Quindi il Golgi è una
sorta di catena per il
completamento delle
proteine.
Il trasporto delle proteine dal reticolo endoplasmatico al Golgi avviene all'interno di membrane
formatesi per gemmazione dal reticolo endoplasmatico e viaggianti lungo i microtubuli.
Le vescicole che portano le proteine da e verso il Golgi sono rivestite esternamente da proteine, in
particolare da tre tipi di proteine diverse
1) COPII, che indirizza le
vescicole verso il Golgi
2) COPI, che indirizza le
vescicole all'indietro verso il
reticolo endoplasmatico o
dalle cisterne esterne del
Golgi verso quelle più interne.
Questo meccanismo serve a
recuperare le proteine
sfuggite dal reticolo
endoplasmatico.
3) clatrina, che indirizza le
vescicole verso endosomi e
lisosomi. Da qui saranno
secrete in maniera regolata.
I lisosomi sono organelli digestivi, contengono enzimi che lavorano bene a pH acido. Possono anche
digerire organelli della cellula (autofagia) per assicurare un turnover. Un lisosoma che ha digerito un
organello può essere espulso dalla cellula (esocitosi) o trattenuto a formare granuli di lipofuscina.
L'accumulo di granuli di lipofuscina nei neuroni (cellule a lunga vita) è correlato all'invecchiamento.
Le proteine destinate ai lisosomi sono glicosilate e alcuni di questi residui sono fosforilati a
mannosio-6-fosfato.
Lipofuscina: accumulo di molecole non più degradabili. L’accumulo è legato all’invecchiamento
Difetti negli enzimi lisosomiali o nei mecanismi che indirizzano gli enzimi lisosomiali ai lisosomi danno
luogo a malattie da accumulo lisosomiale, in cui nella cellula si accumulano lisosomi contenenti
materiale non digerito. La tossicità di queste sostanze porta alla morte cellulare.
Per diagnosticare precocemente queste
malattie è disponibile la diagnosi
prenatale (amniocentesi).
A proposito di eliminazione della spazzatura…
la fagocitosi è realizzata da macrofagi e neutrofili che circolano nel sangue e fagocitano organismi
estranei, cellule danneggiate, globuli rossi invecchiati e detriti. La fagocitosi è quindi un meccanismo di
protezione e il materiale estraneo è digerito dagli enzimi lisosomiali.
Non tutti i batteri inglobati dai macrofagi riescono ad essere digeriti, ad es., il Mycobacterium
tuberculosis (responsabile della tubercolosi), il Coxiella burnetii (responsabile della febbre Q) e la
Listeria monocytogenes (responsabile della meningite)
non si fondono con i lisosomi o non vengono digeriti e
riescono a sfuggire o a riprodursi all'interno del macrofago.
Batterio della tubercolosi
Il citoscheletro
La cellula possiede una struttura semirigida capace di sostenerne la forma ma anche permetterle di
modificarla. Questa impalcatura fornisce il supporto strutturale alla cellula, ne conferisce la resistenza alle
forze che tendono a deformala, determina la posizione dei vari organelli all'interno della cellula, crea una
rete di percorsi per guidare i movimenti di materiali e di organelli, permette lo spostamento delle cellule
(es. spermatozoi, globuli bianchi, fibroblasti, assoni in accrescimento), permette la divisione cellulare
separando i cromosomi e poi la cellula madre in due cellule figlie.
Questa impalcatura, detta citoscheletro, è composta da tre strutture:
1) i microfilamenti o filamenti sottili, strutture piene e sottili composte da actina, implicati nella motilità
e nella contrattilità
2) i microtubuli, lunghi
e rigidi tubi composti
da tubulina, implicati
nel supporto, trasporto
intracellulare e
nell'organizzazione
cellulare. In particolare,
sono formati da
subunità di tubulina
alpha e beta.
I microtubuli sono
connessi e mantenuti
paralleli da ponti
trasversali formati da
proteine.
3) i filamenti
intermedi, robuste
fibre implicate nel
supporto strutturale
della cellula
Disposizione dei componenti del citoscheletro
La tubulina
citoscheletrica di
alcune cellule
endoteliali
marcata in verde,
l'actina in rosso,
osservata in
microscopia
confocale.
Microfilamenti
Microtubuli
Foto di fibroblasti, in verde i microtubuli, in rosso i bordi di contatto tra cellule, in blu il DNA.
Filamenti intermedi
Devono il nome al fatto che hanno spessore intermedio tra microtubuli e microfilamenti, formano
strutture di sostegno per sopportare stress meccanici, ancorano strutture quali il nucleo e i desmosomi
I filamenti intermedi sono costituiti da proteine specifiche per alcuni tipi cellulari
(utili per la tipizzazione)
I filamenti intermedi connettono i desmosomi
Epidermide
Lungo i microtubuli e i microfilamenti scorrono proteine che fungono da trasportatori di carichi
convertendo l'energia chimica in energia meccanica. Il moto è unidirezionale. Queste proteine
trasportatrici che si muovono lungo il citoscheletro sono:
1) chinesine, si muovono lungo i microtubuli, negli assoni spostano vescicole e organelli dal centro della
cellula alla membrana plasmatica
2) dineine, si muovono lungo i
microtubuli, implicata nella disposizione
del fuso mitotico e nel movimento dei
cromosomi nella mitosi, implicata nel
posizionare il complesso di Golgi e nel
movimento di vescicole e organelli nel
citoplasma, trasporta il virus HIV fino al
nucleo. Probabilmente si muove in
direzione opposta rispetto alla chinesina
3) miosine, si muovono lungo i
microfilamenti, sono implicate nella
contrazione muscolare, nella divisione
cellulare, per generare tensione alle
adesioni focali
Miosine
Il taxolo, un principio antitumorale, inibisce il
disassemblaggio dei microtubuli impedendo alla
cellula di assemblare nuove strutture.
In presenza di questo principio chimico le cellule
normali bloccano la loro divisione mentre le cellule
tumorali continuano a dividersi pur non avendo un
fuso mitotico funzionante e muoiono.