Che cosa è il ciclo cellulare?
E’ il processo con il quale le cellule si dividono e si
moltiplicano, duplicando le informazioni genetiche
racchiuse nel loro nucleo. E’ così che dall’uovo fecondato
hanno origine circa i 100mila miliardi di cellule presenti in
un organismo adulto.
Dalle amebe all’uomo, il ciclo cellulare avviene nello stesso
modo in tutte le cellule eucariotiche. Le uniche differenze
riguardano i tempi.
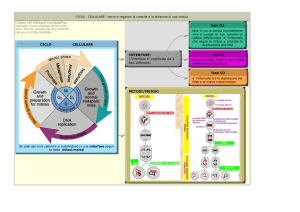
CICLO CELLULARE
Tutte le cellule di un individuo pluricellulare, ad
eccezione dei gameti, cioè le cellule somatiche e le cellule
germinali immature, contengono lo stesso numero di
cromosomi (diploide). Il processo di divisione per
MITOSI assicura che il numero diploide di cromosomi,
caratteristico della specie, sia mantenuto costante
durante tutte le successive divisioni delle cellule, dallo
zigote all’individuo adulto.
I gameti (cellula uovo e spermatozoo) hanno un numero
aploide di cromosomi. Il processo mediante il quale il
numero diploide di cromosomi delle cellule somatiche si
riduce alla metà nelle cellule germinali è un tipo di
divisione speciale, detta MEIOSI.
La modalità di divisione cellulare che prevede la
replicazione del DNA seguita dalla mitosi è
responsabile:
dell’accrescimento e del mantenimento della massa
corporea degli organismi pluricellulari
della sopravvivenza di popolazioni di organismi
unicellulari
Una seconda modalità di divisione cellulare, la
replicazione seguita da meiosi, si riscontra
esclusivamente negli eucarioti a riproduzione sessuata.
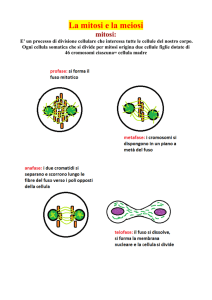
La divisione cellulare per mitosi ha luogo mediante una
successione continua di stadi che nel complesso vengono
chiamati ciclo cellulare.
LABILI: cellule epiteliali, cellule ciclanti (cellule
ematopoietiche, spermatogoni). La durata di G1 è di
poche ore. Si dividono continuamente passando da un
ciclo al successivo.
STABILI: osteoblasti, cellule epatiche, linfociti. Il
periodo G1 si prolunga per giorni, anche anni. Dopo un
certo numero di mitosi entrano in una fase quiescente,
di transitoria fuoriuscita dal ciclo (G0).
PERENNI: fibre muscolari, neuroni, granulociti. Cellule
che hanno un elevato grado di differenziazione, non si
dividono più ed escono irreversibilmente dal ciclo,
arrestandosi permanentemente in fase pre-sintetica
(G2?).
Il periodo intercinetico può essere suddiviso in:
G1 (Gap, intervallo) o periodo pre-sintetico.
Trascrizione dell’RNA, sintesi proteica.
S (Sintesi). Duplicazione del DNA; sintesi di
istoni e di una gran parte delle proteine acide.
G2, periodo post-sintetico. Sintesi di gran
parte dei componenti dell’apparato mitotico, dei
componenti di membrana che verranno utilizzati
per le due cellule figlie, degli induttori della
divisione.
L’interfase occupa oltre il 90% dell’intero ciclo
cellulare. La sua durata e quindi la frequenza
della mitosi è piuttosto costante in cellule dello
stesso tipo ma diversa nei vari tipi cellulari.
Nelle cellule dei mammiferi la durata del periodo
intercinetico dipende dalla durata di G1 in
quanto la durata dei periodi S e G2
è
relativamente costante.
G1: 12-24h
S:
6-7h
G2: 4-6h
M: 1-2h
FASI DELLA MITOSI
Durante il primo stadio della mitosi:
graduale riduzione della trascrizione degli RNA fino
al completo arresto al termine della profase,
scomparsa del nucleolo;
sintesi proteica a livelli minimi;
rallentamento o arresto del traffico vescicolare;
blocco dell’esocitosi e dell’endocitosi;
disgregazione della m. nucleare;
assemblamento del fuso.
APPARATO MITOTICO
Centrioli
Astrosfera
Fuso mitotico
Il FUSO MITOTICO è responsabile dei movimenti
cromosomici durante la mitosi
I cinetocori nel lievito sono piccoli e ciascuno è legato a un solo MT, mentre nelle
cellule di mammifero i cinetocori sono molto più grandi e ognuno è attaccato a 3040 MT del fuso.
LE COESINE
Gruppo di proteine, localizzate al centro del
centromero, funzionano come colla che tiene insieme i
cromatidi prima dell’inizio dell’anafase e sono
degradate all’inizio dell’anafase, consentendo così la
separazione dei cromatidi fratelli.
I MT polari prendono contatto con i MT provenienti dal
centrosoma opposto. Quando le due estremità positive di
due MT di polarità opposta iniziano a sovrapporsi,
intervengono delle proteine che le legano insieme
formando dei legami crociati. Come il legame tra
cinetocore e i MT del cinetocore, questi legami crociati
stabilizzano i MT polari.
Possiamo immaginare che i MT durante la profase e la
prometafase escono rapidamente a raggiera dai due
centrosomi, quelli che riescono a incontrare un
cinetocore o un MT di polarità opposta si stabilizzano;
gli altri si disassemblano e si ritirano.
allineamento e separazione dei cromosomi
La combinazione di forze che tirano e spingono i
cromosomi li fa disporre sulla piastra metafasica, la
loro localizzazione più stabile, dove si allineano in
ordine casuale.
All’inizio dell’anafase, la regione centromerica di
ciascun cromosoma si divide in due, consentendo ai due
cromatidi fratelli appaiati di separarsi e di muoversi
verso i poli opposti del fuso. La topoisomerasi II è uno
degli enzimi coinvolti.
proteine motrici e movimenti cromosomici
Le proteine motrici hanno almeno tre distinti ruoli nel movimento
dei cromosomi anafasici
Anafase A
Le proteine motrici del cinetocore generano la forza
motrice, l’accorciamento dei MT è un evento secondario.
Anafase B
Le proteine motrici associate ai MT polari rappresentano
la forza primaria che fa allungare il fuso, mentre
l’allungamento dei MT polari è secondario.
CITOCINESI o CITODIERESI: divisione della cellula
CARIOCINESI: divisione del nucleo
La posizione del fuso
determina il punto in
cui si dividerà il
citoplasma.
Anello contrattile:
fibre di actina
La polimerizzazione dei monomeri di actina in
microfilamenti
avviene
subito
prima
dell’invaginamento che dà inizio alla formazione
del solco di clivaggio e lo smantellamento
dell’intera struttura avviene poco dopo il
completamento della citocinesi.
La CITOCINESI nelle nelle cellule vegetali avviene
per assemblaggio di una membrana plasmatica e di
una parete cellulare tra i due nuclei figli.
Appena i nuclei figli riprendono la loro struttura
caratteristica:
riparte la trascrizione dell’RNA, riappare il
nucleolo;
la sintesi proteica riprende a velocità normale, così
come il traffico vescicolare, l’endocitosi e l’esocitosi.
G1: 12-24h
S:
6-7h
G2: 4-6h
M: 1-2h
La variabilità del ciclo cellulare (lunghezza
relativa delle varie fasi e la vicinanza temporale
tra mitosi e citocinesi) indica l’esistenza di
meccanismi di regolazione che si adattano alla
necessità di un particolare tipo cellulare o
specie.
Il sistema di controllo che regola la progressione del
ciclo cellulare deve:
1) Garantire che tutti i processi associati con le diverse
fasi siano portati a termine al tempo dovuto e nella
sequenza corretta;
2) Assicurare che ogni fase del ciclo sia stata
completata correttamente prima che inizi la
successiva;
3) Essere in grado di rispondere alle condizioni esterne
che segnalano alla cellula di crescere e di dividersi.
L’importanza relativa dei punti di controllo di
G1 e G2 nella regolazione del tasso di divisione
cellulare varia da cellula a cellula e nei diversi
organismi.
.
Esperimenti di fusione cellulare
forniscono
la prova dell’esistenza di
• .
molecole che controllano il ciclo
cellulare.
Fondendo cellule in fase S con cellule in
fase G2 le cellule in G2 non duplicano il
DNA.
Questi esperimenti suggerivano che nel
citoplasma dovessero esistere “molecole segnale”
responsabili del passaggio oltre i punti di
controllo G1 e G2, cioè gli induttori della sintesi
del DNA (fase S) e della mitosi fossero delle
molecole specifiche presenti nel citoplasma
(MPF).
Sebbene i fattori in grado di modificare la progressione
delle cellule attraverso il ciclo di divisione cellulare siano
numerosi, i meccanismi di base che controllano la
divisione cellulare sono genetici.
H. Hartwell (USA) Saccaromyces cerevisiae
P.M. Nurse (UK) Saccaromyces pombe
cdc28
cdc2 cdk1
R.T. Hunt (UK) stelle marine e ricci di mare
cicline
Nobel per la medicina, 2001
cdc2 è un gene la cui attività è essenziale per l’entrata
in mitosi (cioè oltrepassa il punto di controllo G2)
I risultati delle ricerche sui lieviti si unirono a quelli
sulla rana e si arrivò a stabilire che:
cdc2 codifica una delle due proteine che fanno
parte di MPF
La proteina codificata dal gene cdc2 del lievito
funziona come una proteina chinasi, cioè un enzima che
catalizza il trasferimento di un gruppo fosfato dall’ATP
a determinate proteine bersaglio.
Questa proteina funziona solo quando è legata a un
membro di un’altra famiglia di proteine, le cicline, e
dunque è una chinasi ciclina-dipendente (cdk).
Le cicline sono proteine che, attraverso oscillazioni
della loro concentrazione intracellulare, controllano
l’attività delle varie molecole cdk ai diversi stadi del
ciclo. Un secondo tipo di regolazione per le cdk è basato
sulla fosforilazione delle stesse molecole cdk.
I complessi cdk-cicline hanno in comune:
la loro attività è regolata da reazioni di fosforilazione
e defosforilazione catalizzate da proteine chinasi e
proteine fosfatasi;
le cicline sono sintetizzate e degradate durante le
diverse fasi del ciclo.
PUNTO DI CONTROLLO G2
MPF attivo:
fosforila le lamìne della lamina nucleare interna. Ciò
determina la depolarizzazione delle lamine che porta alla
disgregazione
della
lamina
nucleare
(destabilizzazione
dell’involucro nucleare).
fosforila alcune proteine cromosomiche, tra cui l’istone H1 ed il
complesso multiproteico (complesso della condensina) favorendo
la condensazione cromosomica.
fosforila proteine associate ai microtubuli per la formazione del
fuso
controlla i meccanismi che assicurano che il DNA sia stato
completamente replicato prima di consentire alla cellula di
oltrepassare il punto di controllo G2 ed entrare in mitosi.
Anche gli altri punti di controllo del ciclo cellulare sono
regolati da complessi Cdk-cicline. Anche se nei diversi
organismi le componenti sono un po’ diverse, tutti gli
eucarioti possiedono una gamma di molecole Cdk e di
cicline che interagiscono in combinazioni diverse ai vari
stadi del ciclo cellulare.
I complessi cdk-cicline hanno in comune:
la loro attività è regolata da reazioni di fosforilazione
e defosforilazione catalizzate da proteine chinasi e
proteine fosfatasi;
le cicline sono sintetizzate e degradate durante le
diverse fasi del ciclo.
PUNTO DI CONTROLLO G1
I diversi segnali (dimensione cellulare etc……)
segnalano alla cellula che è arrivato il momento
di dividersi. Questi segnali funzionano
attivando complessi Cdk-ciclina che inducono
l’entrata in S fosforilando diverse proteine
bersaglio.
Tra metafase e anafase
START
PUNTO DI RESTRIZIONE
PUNTO DI CONTROLLO DELL’ASSEMBLAGGIO DEL FUSO
MPF consente il superamento del punto di controllo
dell’assemblaggio del fuso catalizzando una o più reazioni
di fosforilazione di proteine che portano all’attivazione di
un grosso complesso proteico, il complesso che promuove
l’anafase (APC). Questo complesso degrada, legandoli
all’ubiquitina, gli inibitori dell’anafase e la ciclina
mitotica.
Il volume citoplasmatico è il fattore più critico per la
regolazione del ciclo cellulare negli organismi
unicellulari e svolge anche un ruolo fondamentale nel
controllo della divisione cellulare nelle forme
pluricellulari.
Le componenti fondamentali del meccanismo
di regolazione che promuove o inibisce la
crescita cellulare sono delle molecole segnale
presenti all’esterno della cellula.
Fattori che promuovono la moltiplicazione:
fattori di crescita (ormoni, NGF….)
sostanze nutrienti (aa, vitamine…..)
Fattori che inibiscono la moltiplicazione:
differenziamento
inibizione da contatto
aumento della temperatura
veleni mitotici
agenti chimici e fisici
Schema 1
Il cancro è associato a alterazioni dei meccanismi di
controllo del ciclo cellulare.
mutazioni del DNA a carico di proto-oncogeni, geni
oncosoppressori, geni del riparo del DNA (agenti chimici,
radiazioni,
mutazioni
spontanee,
errori
della
replicazione; alcune mutazioni sono ereditate, alcune
sono introdotte nelle cellule da infezioni virali)
mutazioni di geni che regolano l’apoptosi
Il cancro è associato ad alterazioni dei meccanismi di
controllo del ciclo cellulare
Tumori benigni
Tumori maligni
Mutazioni di uno o più geni deputati al controllo del
ciclo cellulare rappresentano la maggiore causa dei
tumori.
Le mutazioni che inducono il cancro sono a carico di tre
classi di geni: protoncogeni, geni oncosoppressori, geni
del riparo del DNA.
Caratteristiche peculiari delle cellule
neoplastiche:
riprodursi indefinitamente nel tempo
perdita di adesività al substrato
(metatasi)
perdita di inibizione da contatto
Proto-oncogeni
geni che codificano prodotti
coinvolti nell’induzione della
proliferazione cellulare
Oncosoppressori
geni che si oppongono alla
proliferazione indefinita e
promuovono l’arresto della crescita
cellulare e possono favorire
l’apoptosi (es. p53)
Un oncogene è un gene la cui presenza può indurre lo
sviluppo di un tumore. Alcuni oncogeni derivano da
mutazioni di normali geni cellulari (protoncogeni) mentre
altri sono introdotti nelle cellule da virus oncogeni (che
inducono i tumori). In entrambi i casi gli oncogeni
codificano per proteine che stimolano in modo eccessivo la
proliferazione cellulare.
Alcuni oncogeni codificano per forme alterate e
iperfunzionanti della proteina, mentre altri
inducono la iperproduzione della forma normale
della proteina. In entrambi i casi il risultato
finale è una iperstimolazione della via del
segnale mediato dai fattori di crescita e quindi
una eccessiva proliferazione cellulare.
Quali tipi di mutazioni possono trasformare un
gene normale (proto-oncogene) in un gene che
induce lo sviluppo di un tumore, e cioè un
oncogene?
Alcune mutazioni che inducono il cancro sono
provocate da agenti chimici che danneggiano il DNA e
dalle radiazioni, altre derivano da mutazioni
spontanee del DNA e da errori della replicazione.
Alcune mutazioni che contribuiscono allo sviluppo dei
tumori possono anche essere ereditate o essere
introdotte nelle cellule da infezioni virali.
Quali tipi di mutazioni possono trasformare un gene
normale (proto-oncogene) in un gene che induce lo
sviluppo di un tumore, e cioè un oncogene?
4 meccanismi diversi:
mutazione puntiforme (es. oncogeni ras)
riarrangiamenti del DNA:delezioni o scambi di
sequenza tra un proto-oncogene e i geni vicini (es.
oncogene trk)
amplificazione genica (es. alcune forme di tumore del
polmone: amplificazione c-myc)
traslocazione cromosomica (es. linfoma di Burkitt. Un
tratto crom.8 trasloca sul crom. 2,14 o 22)
Tab.2
La maggior parte dei tumori umani non è
associata a infezioni virali.
Virus del papilloma umano (HPV)
Virus di Epstein Barr (EBV) (linfoma di Burkitt)
Un tumore può essere causato anche dalla
assenza o inattivazione di geni oncosoppressori.
Il gene p53 è uno dei geni più frequentemente mutato nei
tumori umani.
p53 è la proteina “guardiano del genoma” protegge la
cellula dagli effetti dei danni al DNA.
Con la sua capacità di indurre l’arresto del ciclo cellulare
e/o la morte cellulare, la proteina p53 funziona come un
semaforo rosso che, in presenza di danni al DNA, blocca
la proliferazione cellulare impedendo alla cellula di
trasmettere il danno alle cellule figlie.
La perdita della funzione di p53 può contribuire
allo sviluppo di un tumore in quanto consente la
sopravvivenza e la riproduzione di cellule che
hanno danni al DNA.
Fasi dell’apoptosi:
•Disgregazione del nucleolo
•Taglio lamina
•Condensazione e taglio della cromatina in frammenti di 180-200
paia di basi
•Migrazione dei granuli compatti di cromatina degradata verso la
periferia del nucleo e verso la membrana plasmatica dove vengono
circondati da evaginazioni della membrana che conferiscono alla
cellula un aspetto a bolla (blebbing)
•Le blebs si staccano dal corpo cellulare trascinando con sé
citoplama e materiale cellulare-corpi apoptotici-fagocitosi
Durata del processo:
1-2 hr
IL CICLO CELLULARE MEIOTICO
Nella specie umana:
le cellule somatiche (tutte le cellule dell’organismo ad
eccezione dei gameti e dei loro immediati progenitori)
sono diploidi, cioè contengono due esemplari di ciascun
cromosoma (uno di origine materna e uno di origine
paterna). In totale contengono 46 cromosomi (22 paia di
autosomi e 1 paio di cromosomi sessuali). I due
esemplari di ciascun cromosoma costituiscono una
coppia di “omologhi”.
i gameti sono aploidi (contengono un solo esemplare
di ciascun cromosoma) e hanno perciò 23 cromosomi.
Corredo cromosomico: l’insieme dei cromosomi in una
cellula.
Cariotipo: l’insieme delle caratteristiche (numero e
morfologia) che identificano un particolare corredo
cromosomico.
Corredo e cariotipo sono caratteristici della specie.
I cromosomi omologhi sono simili per
dimensioni, forma e posizione dei loro
centromeri.
Portano l’informazione per il controllo degli
stessi tipi di caratteristiche genetiche, sebbene
non necessariamente la stessa informazione
genetica. (da ricordare che: i cromatidi fratelli
sono perfettamente identici)
Diploide (2n) : è una cellula che contiene 2 serie
di cromosomi (2 per ogni tipo).
Aploide (n): è una cellula che possiede solo un
cromosoma di ogni coppia di omologhi.
Poliploide: è una cellula che contiene 3 o più
serie di cromosomi (rara negli animali, comune
nelle piante)
La MEIOSI si verifica solo nelle cellule
germinali (o gameti) degli animali e delle piante
a riproduzione sessuata. Negli organismi
unicellulari (es. protozoi) la meiosi si verifica
solo nei casi di riproduzione sessuale.
La meiosi consiste in due divisioni consecutive
precedute da una sola duplicazione del DNA.
div. riduzionale
div. equazionale
Ciascuna delle cellule aploidi prodotta dalla
meiosi II contiene un assortimento casuale di
cromosomi paterni e materni. Ciascun
cromosoma è inoltre il risultato della
combinazione di sequenze di DNA materne e
paterne creata dal crossing over durante la
profase I.
FASI DELLA MEIOSI
I div. Meiotica
interfase o preleptotene 20-30 hr (duplicazione DNA)
profase I leptotene
1-2
zigotene
2-3
pachitene
7-9
diplotene
0.5-1
diacinesi
0.5
metafase I
anafase I
telofase I
II div. Meiotica
interfase II (no duplicazione DNA)
profase II
metafase II
anafase II
telofase II
In meiosi I:
Ricombinazione genetica
COMPLESSO SINAPTINEMALE
Leptotene-zigotene
All’inizio dello zigotene i telomeri dei singoli
cromosomi si aggregano in una regione del nucleo e si
attaccano alla membrana nucleare mentre la parte
centrale del cromosoma sporge nel nucleo. Questa
disposizione dei cromosomi, detta bouquet, facilita
l’allineamento dei cromosomi. L’allineamento dei
cromosomi di lunghezza simile facilita la formazione
dei complessi sinaptonemali durante il pachitene.
Negli eucarioti superiori la formazione dei complessi
sinaptonemali è strettamente associata con il processo
del crossing over, ma in tale processo sono coinvolti
anche altri complessi proteici, detti noduli di
ricombinazione.
CROSSING OVER= interscambio di segmenti tra i due
cromosomi omologhi
Come avviene:
rottura trasversale allo stesso livello su due cromatidi
non fratelli;
trasposizione, rispetto al centromero, dei segmenti distali;
saldatura dei segmenti.
Tutto ciò richiede l’intervento di:
endonucleasi, esonucleasi, DNA polimerasi, ligasi.
Ciascuna delle cellule aploidi prodotta dalla
meiosi II contiene un assortimento casuale di
cromosomi
paterni
e
materni
(dovuto
all’orientamento casuale dei bivalenti alla
metafase I). Ciascun cromosoma è inoltre il
risultato della combinazione di sequenze di DNA
materne e paterne creata dal crossing over
durante la profase I.
Uniti dai chiasmi
Risultato della mitosi: due cellule ciascuna con lo
stesso numero di cromosomi della cellula parentale.
Risultato della meiosi: quattro cellule aploidi
ciascuna con la metà dei cromosomi rispetto alla
cellula parentale. Ciascuna cellula aploide contiene
un assortimento casuale di cromosomi materni e
paterni.