LA
BIOCHIMICA
DEL
MUSCOLO
Le attuali conoscenze sulla contrazione muscolare si
basano
sul
modello
a
scorrimento
dei
filamenti
.
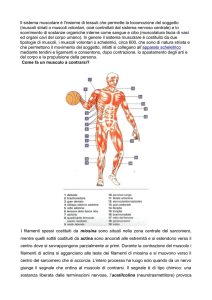
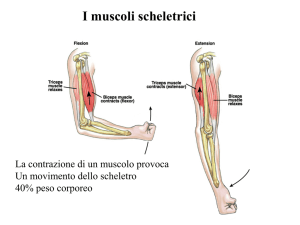
Il
muscolo
scheletrico
comprende
circa
il
40%
della
massa
corporea
ed
è
formato
da
cellule
cilindriche
multinucleate
chiamate
fibre
muscolari.
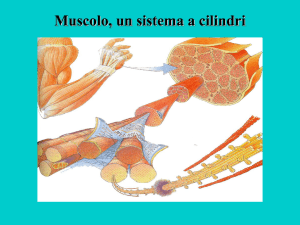
La membrana plasmatica delle fibre muscolari è
conosciuta come sarcolemma. Dentro il sarcolemma è
contenuto il sarcoplasma che contiene tutti i comuni
elementi
subcellulari
più
delle
lunghe
miofibrille.
• Ciascuna miofibrilla è composta da fasci di proteine
filamentose contrattili ed è costituita da corte unità
strutturali
note
come
sarcomeri.
• Le
proteine
alle
giunzioni
dei
sarcomeri
formano
le
linee
Z, per cui un sarcomero si estende lungo una
miofibrilla
da
una
linea
Z
ad
un’altra
linea
Z.
• I sarcomeri sono composti principalmente da sottili
filamenti
di
actina
e
da
filamenti
spessi
di
miosina.
I
sarcomeri
rappresentano
la
minima
unità
contrattile
del
muscolo. La contrazione coordinata di milioni di
sarcomeri in un muscolo è la base dell’attività
meccanica
del
muscolo.
Il
tessuto
connettivale
che
avvolge
la
miofibra
viene
denominato
sarcolemma.
La
diramazione
di
questo
tra
i
fasci
prende
il
nome
di
perimisio
che
a
sua
volta
diventa
epimisio
quando
avvolge
l'intero
muscolo.
Il
liquido,
che
circonda
le
miofibre
e
che
contiene
anche
i
substrati
energetici
del
muscolo,
viene
definito
sarcoplasma.
Fibra
Muscolare
All’interno del sarcomero i filamenti spessi e sottili sono
intervallati in modo tale che facendo una sezione trasversale
del sarcomero appare un motivo a reticolo esagonale, in cui 6
filamenti sottili circondano ciascun filamento spesso.
Durante la contrazione ed il rilassamento la distanza tra le
linee Z varia, diminuendo durante la contrazione. I filamenti
spessi e sottili riprendono poi le loro posizioni a seguito del
rilassamento, salvo il verificarsi di situazioni estreme.
Il cambiamento nella lunghezza del sarcomero è determinata
dallo scorrimento dei filamenti sottili su quelli spessi; i filamenti
sottili si spostano verso la zona centrale del sarcomero
(linea M).
Ciascuna
miofibrilla
è
formata
da
diverse
proteine:
a) Miosina
e
Actina
che
sono
proteine
contrattili
b) Tropomiosina
e
Troponina
che
sono
proteine
regolatorie
c) Titina
e
Nebulina
che
sono
proteine
accessorie
giganti
PROTEINE DEI MIOFILAMENTI
La base biochimica dell’attività muscolare è collegata alle proprietà
enzimatiche e fisiche di actina, miosina e proteine accessorie.
MIOSINA: lunga proteina fibrosa con peso molecolare di
circa 500.000 daltons.
Ciascuna molecola di miosina è costituita da 6 subunità:
2 catene pesanti (HC) molto grandi e
4 catene leggere (LC) più piccole.
La HC contiene un lungo dominio terminale in alfa-elica (circa 1300
amminoacidi) e un dominio globulare (testa globulare) all’estremità
ammino-terminale (circa 800 amminoacidi). Le due catene pesanti (HC)
hanno i tratti in alfa-elica superavvolti a dare un lungo tratto rigido con due
teste globulari.
Una molecola di miosina completa contiene anche 4 catene leggere (LC)
associate alle teste globulari delle catene pesanti. Queste piccole proteine
(LC) hanno peso molecolare di 16.000-24.000 daltons e si suddividono in
LC1, LC2 ed LC3.
Ciascun dominio globulare presenta 1LC2 e 1LC1 o 1LC3 a seconda del
tipo di muscolo considerato (cardiaco, scheletrico, embrionale, liscio).
Tutte le catene leggere legano il Ca2+ con una elevata affinità e servono
per regolare l’attività ATPasica della miosina, nonché l’assemblaggio della
miosina per formare i filamenti spessi.
L’attività ATPasica della miosina è localizzata a livello delle teste
globulari.
ACTINA
I filamenti sottili sono composti da molte subunità di
una proteina globulare nota come actina-G (42 kD) e
diverse proteine accessorie.
In un filamento sottile, l’actina-G è polimerizzata in lunghi
segmenti fibrosi noti come actina-F.
Un paio di filamenti di actina-F sono avvolti elicoidalmente
per formare la struttura portante di un filamento completo di
actina. Ciascun monomero di actina-G contiene un sito di
legame per la testa della miosina.
Le principali proteine accessorie presenti nei filamenti
sottili sono:
Tropomiosina
e
Troponina.
Tn-T: lega la troponina alla tropomiosina
Tn-I: lega la troponina all’actina
Tn-C: sito di attacco per il calcio
La tropomiosina è un eterodimero costituto da due filamenti superavvolti.
Nel muscolo rilassato ciascuna molecola di tropomiosina copre i siti di
legame con la miosina di 7 residui di actina-G, prevenendo il legame tra
actina e miosina e mantenendo il muscolo rilassato.
L’inizio di un evento di contrazione richiede l’intervento della seconda
proteina accessoria, la troponina.
La troponina è un eterotrimero che fisicamente lega la tropomiosina
all’actina.
Cambiamenti conformazionali nella troponina sono responsabili del
movimento della tropomiosina che alternativamente copre e scopre i siti di
legame tra actina e miosina.
Una delle subunità della troponina, la troponina-C (Tn-C), è una proteina
analoga alla calmodulina e quindi in grado di legare il calcio.
Quando Tn-C lega il calcio, l’intera molecola di troponina va incontro a
variazioni conformazionali che muovono la tropomiosina ad essa solidale e
liberando il sito di attacco della miosina sull’actina.
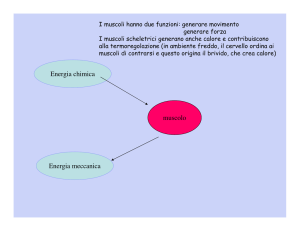
La miosina e l’ATP:
Il muscolo scheletrico converte l’energia chimica in energia meccanica con
altissimo rendimento:non più del 30-50% si dissipa in energia termica.
L’energia necessaria ai fini della contrazione muscolare proviene dall’idrolisi
dell’ATP. L’energia di idrolisi dell’ATP è utilizzata per guidare la miosina da
uno stato conformazionale a più bassa energia a uno ad energia più elevata.
Quando i livelli citosolici di calcio aumentano, e il legame per la miosina
sull’actina diventa accessibile, si forma il complesso ACTOMIOSINA a cui
segue la dissociazione sequenziale di Pi e ADP con ritorno della miosina allo
stato a bassa energia.
Questi eventi sono accompagnati da una simultanea traslocazione dei
filamenti sottili verso il centro del sarcomero (linea M).
Alla fine dell’impulso di forza (power stroke) il complesso actomiosinico
rimane intatto fino a quando non si ha ATP disponibile.
Il legame dell’ATP alla miosina è una reazione fortemente esoergonica con
conseguente distacco dell’actina dalla miosina.
La sequenza di eventi che conducono ad
un evento di contrazione possono essere
così riassunti:
1. una testa miosinica libera lega l’ATP
(stato 1) e lo idrolizza;
2. la miosina si trova a questo punto nella
forma a più alta energia con ADP e Pi legati
(stato 2);
3. la miosina a questo punto si lega
all’actina liberando il Pi (stato 3);
4. appena legatasi in questo modo, la testa
subisce un mutamento di conformazione
che genera una fase utile esercitando una
trazione sul resto del filamento spesso. Al
termine della fase utile (stato 4) si libera
l’ADP e la testa si associa con una nuova
molecola di ATP, dissociandosi dal filamento
di actina e riacquistando lo stato 1.
Gli eventi che riguardano i
filamenti
sottili
durante
la
contrazione possono così essere
riassunti:
1. Prima della comparsa di calcio
libero
nel
sarcoplasma,
la
tropomiosina copre i siti di legame
per la miosina presenti sull’actina;
2. la comparsa del calcio nel
sarcoplasma porta al legame tra il
calcio e Tn-C;
3. le variazioni conformazionali
nella troponina spostano la
tropomiosina e scoprono i siti di
legame sull’actina;
4. i siti esposti sono ora disponibili
per interagire con le teste
globulari della miosina;
5. la rimozione del calcio dal
sarcoplasma ripristina lo stato
conformazionale
originario
di
troponina
e
tropomiosina,
mascherando nuovamente i siti di
attacco della miosina.
Un’istantanea di un filamento spesso mostrerebbe nel muscolo che si contrae
alcune teste agganciate al filamento actinico ed altre disimpegnate.
Ciascun filamento spesso contiene approssimativamente 500 teste miosiniche, e nel
corso di una contrazione rapida ciascuna testa percorre circa 5 cicli al secondo,
facendo scorrere l’uno sull’altro il filamento spesso e quello sottile al ritmo di 15 µm
al secondo.
REGOLAZIONE DEL CALCIO SARCOPLASMATICO:
L’interazione molecolare generatrice di forza che è stata appena descritta ha luogo
solamente a patto che il muscolo scheletrico riceva un segnale dal suo nervo motore.
Tale segnale scatena un potenziale di azione nella membrana cellulare della cellula
muscolare, e l’eccitazione elettrica si diffonde velocemente in una serie di pliche
membranose, i tubuli trasversali o tubuli T, e il segnale viene poi trasmesso al reticolo
sarcoplasmatico.
L’eccitazione induce una depolarizzazione locale del sarcolemma. Si ha
conseguentemente l’apertura dei canali del calcio nel reticolo sarcoplasmatico, seguita
da un rapido e massivo movimento delle cisterne del calcio nel reticolo
sarcoplasmatico vicino alle miofibrille. La comparsa del calcio in prossimità della
subunità Tn-C della troponina produce i power stroke multipli della miosina per il tempo
determinato dal permanere del calcio a livelli superiori a 1-5 µM.
Alla cessazione dell’attività contrattile, le membrane sarcoplasmatiche ritornano al
potenziale elettrico di riposo (circa 60mV), come pure il reticolo sarcoplasmatico.
Il calcio sarcoplasmatico viene ripompato nelle cisterne del reticolo sarcoplasmatico da
una pompa del calcio ATP dipendente. Per ogni ATP idrolizzato, due ioni calcio
vengono estratti dal sarcoplasma e il livello in tale comparto scende alla fine sotto 0.1
µM (in circa 30 millisecondi).
MECCANISMI ENERGETICI
del MUSCOLO
ANAEROBICO ALATTACIDO
ANAEROBICO LATTACIDO
AEROBICO
46
MECCANISMI
ENERGETICI
del
MUSCOLO
L'energia per la contrazione muscolare viene fornita dall’ ATP (Adenosin
trifosfato)
che
si
scinde
in
ADP
(Adenosin‐difosfato)
e
P
(fosfato
inorganico) ATP
=
ADP
+
P
+
energia
La
quantità
di
ATP
presente
nei
muscoli
è
molto
limitata
per
cui
è
necessario
ricostituirla
in
continuazione.
La
resintesi
dell'ATP
avviene
attraverso
tre
diversi
meccanismi,
ognuno
legato
alla
durata
e
all'intensità
dell'impegno
muscolare.
Il
muscolo può utilizzare tutti e tre i sistemi contemporaneamente oppure
privilegiarne
maggiormente
uno
rispetto
altri
due:
1. Sistema
anaerobico
alattacido
2. Sistema
anaerobico‐lattacido
3. Sistema
aerobico
47
1)
Sistema
ATP‐CP
(anaerobico
alattacido)
Questo meccanismo si innesca in assenza di O2 e senza
formazione
di
Acido
lattico
nei
muscoli,
utilizzando
una
molecola
altamente
energetica
immagazzinata
nel
muscolo
la
creatinfosfato
o
fosfocreatina
‐
CP,
la
CP
in
seguito
allo
stimolo
nervoso
libera
una grande quantità di energia scindendosi in creatina (C) e
fosforo
(P),
quest'ultimo
con
l'ADP
va
a
riformare
l'ATP.
C
CP
P
+ ADP
energia liberata
ATP
Sforzi di breve durata
10” – 20”
Contrazioni rapide
intensità massimale
Molta Potenza
48
Questo processo di ricostruzione di ATP è molto rapido, quasi simultaneo,
purtroppo
la
quantità
di
CP
presente
nel
muscolo
è
relativamente
limitata
e
si
esaurisce
in
brevissimo
tempo
(8‐10
secondi).
Questo sistema consente al muscolo di eseguire contrazioni molto rapide,
anche
d’intensità
massimale,
ma
per
periodi
di
tempo
assai
limitati
(corse
di
velocità
fino
a
100
mt.,
salti,
lanci
etc.)
che
richiedono
un
impiego
d’energia
massimale.
L'energia
spesa
viene
ripristinata
dopo
circa
3
minuti.
L'utilizzazione
di
questo
sistema
può
andare
ben
oltre
gli
8‐10
secondi
qualora
l'impiego muscolare sia tale da non richiedere la massima potenza del
processo,
ma
percentuali
più
basse
(durata
massima
40
–
45
secondi).
49
2)
Sistema
dell’ac.
lattico
(anaerobico
lattacido)
Quando
lo
sforzo
si
protrae
nel
tempo
e
l’atleta
ha
esaurito,
tutte
le scorte di CP presenti nel muscolo e quindi non può più
ricostituire
l'ATP
con
le
proprie
riserve
chimiche,
non
cessa
la
sua
attività, ma riesce a continuarla perché subentra il sistema
dell'acido lattico o glicolisi (in assenza di O2), che produce una
sostanza detta Acido Lattico (sostanza tossica, il cui accumulo nei muscoli
provoca fenomeni di affaticamento che costringono l’atleta a ridurre l’intensità dello
sforzo,
fino
al
blocco
totale
dell’attività
muscolare).
Questo meccanismo, che utilizza l’energia liberata dalIa
demolizione delle molecole di GLUCOSIO (presenti nei muscoli) e di
GLICOGENO (accumulato nel fegato) tramite reazioni chimiche
accelerate
da
particolari
enzimi,
permette
la
ricostituzione
di
ATP
ma
produce
anche
acido
lattico
50
Il
processo
anaerobico
lattacido
è
di
fondamentale
importanza
per
compiere prestazioni fisiche nelle seguenti specialità sportive:
quelle
individuali
continuative
sub‐massimali
di
durata
compresa
tra i 40 ‐ 45 secondi e i 4 minuti circa; quelle di squadra con
riferimento agli atleti che forniscono un impegno intenso e
continuo.
Fegato sotto forma di glicogeno
ACIDO LATTICO
Accumulato nei muscoli
ZUCCHERI
Sforzi di
media durata
GLICOGENO
+
ENZIMI
15” – 45”
Intensità elevata
+ ADP
energia liberata
ATP
51
CONOSCERE IL CORPO UMANO: AEROBICO
3)
Sistema
dell’ossigeno
(aerobico)
Se
la
quantità
di
ATP
richiesta
dal
muscolo
per
svolgere
la
sua
attività non è molto elevata, l'ossigeno (O2) che viene
immesso
nel
nostro
organismo
per
mezzo
della
respirazione
ha
la
possibilità
di
ossidare
(combinare)
le
sostanze
presenti
(zuccheri,
proteine
e
grassi)
e
di
riformare
ATP
producendo
sostanze
di
rifiuto
quali
l’anidride
carbonica
(CO2)
e
l’acqua
(H2O)
che
sono
espulsi
mediante
la
respirazione
(polmoni)
e
la
sudorazione.
In
tale
situazione
il
lavoro
muscolare
può
essere
protratto più a lungo, teoricamente senza alcun limite.
Utilizzando
questo
sistema,
la
quantità
d’ossigeno
trasportata
ai muscoli non è mai inferiore a quella necessaria per
riformare
l'ATP
e
quindi
l'organismo
può
lavorare
in
"steady‐
state"
cioè
in
stato
d’equilibrio.
52
Il
limite
di
questo
processo
energetico
è
la
lentezza
(ci
vuole
molto
tempo
affinchè
l’
CO2 venga
espulsa
e
l’O2
arrivi
ai
muscoli.
Se
lo
sforzo
si
intensifica
(maggiore
consumo
di
O2),
si
creerà
un
accumulo
di
CO2
e
contemporaneamente
attraverso
il
processo
di
scissione
del
glicogeno,
un
accumulo
di
ac.
lattico
che
costringe
il
corpo
ad
una
richiesta
superiore
di
O2
definita:
“DEBITO
di
OSSIGENO”
E’
necessario
quindi
rallentare
il
lavoro
in
modo
da
dare
tempo
al
sangue
e
ai
polmoni
di
espellere
l’CO2,
di
trasportare
al
fegato
l’ac.
Lattico
e
di
immettere
O2
con
l’inspirazione,
questo
tempo
di
attesa
si
definisce:
“PAGARE
IL
DEBITO
DI
OSSIGENO”
O
periodo
di
tempo
necessario
per
ripristinare
l’equilibrio
tra
consumo
di
O2
e
immissione
di
O2
e
poter
riprendere
il
lavoro
muscolare.
53
CO2
O2
OSSIDA
Sforzi di
lunga durata
Superiori a 180”
Intensità moderata
ZUCCHERI
GRASSI
PROTEINE
H 2O
ESPULSA
con L’ESPIRAZIONE
Anidride Carbonica
ESPULSA
con LA SUDORAZIONE
Acqua
+ ADP
energia liberata
ATP
54
Scambi
metabolici
tra
fegato
e
muscoli
Particolare
importanza
hanno
gli
scambi
che
avvengono
tra
il
fegato
e
i
muscoli
e
che
riguardano
il
metabolismo
degli
zuccheri.
Il
fegato
può
rilasciare
nel
sangue
glucosio
che
ricava
scindendo
le
sue
scorte
di
glicogeno
(glicogenolisi)
oppure
sintetizzandolo
a
partire
da
vari
precursori
(gluconeogenesi);
tra
questi,
il
lattato
e
l'alanina,
forniti
principalmente
dai
muscoli.
Il
glucosio
immesso
nel
torrente
circolatorio
può
raggiungere
le
fibre
muscolari
ed
essere
impiegato
come
fonte
di
energia
per
la
contrazione.
Durante
un'intensa
attività,
i
muscoli scheletrici ottengono rapidamente energia attraverso il processo anaerobico di
glicolisi;
l'acido
lattico
che
ne
deriva
ritorna
al
fegato,
dove
viene
riconvertito
in
glucosio.
La
transaminazione
permette
la
conversione
dell'amminoacido
alanina
in
acido
piruvico
e
viceversa.
55
F
O
N
T
E
d
I
Sistema
ATP‐CP
(anaerobico
alattacido)
Sistema anaerobico lattacido
Glicolisi
Glicogeno
muscolare
E
N
E
R
G
I
A
Sistema aerobico
TEMPO
MECCANISMI ENERGETICI del MUSCOLO 56
IN RAPPORTO ALLA DURATA NEL TEMPO
CLASSIFICAZIONE BIOENERGETICA DELLE ATTIVITA’ SPORTIVE
PROCESSO
COMBUSTIBILE
POTENZA
DURATA
TIPO di SPORT
REAZIONE di BASE
ATP
ALTISSIMA
FINO A 3”
GESTI SINGOLI 8salti, lanci, tuffi)
ATLETICA LEGERA
ANAEROBICO ALATTACIDO
DISGREGAZIONE
della Fosfocreatina
(CP)
ALTA
10” – 15”
100 e 110 hs
Lanci (disco, giavellotto, martello, peso)
Salti (alto, lungo, triplo, asta)
SOLLEVAMENTO PESI -PATTINAGGIO (velocità)
ATLETICA LEGERA
800 – 1500 – 400 hs.
ANAEROBICO LATTACIDO
SCISSIONE
del GLICOGENO
GLICOLISI
ELEVATA
15” – 45”
PATTINAGGIO
Ghiaccio 3000 mt. - Rotelle 1500 mt.
NUOTO
400 mt.
ATLETICA LEGERA
200 e 400 piani
ANAEROBICI
AEROBICI MASSIVI
SCISSIONE
del GLICOGENO
GLICOLISI
ELEVATA
45” – 180”
PATTINAGGIO
Ghiaccio 5 - 10 Km. - Rotelle 3 – 20 Km.
NUOTO
50 e 100 mt. stile libero
ATLETICA LEGERA
3.000 siepi, 5.000 mt., 10.000 mt., maratona, marcia
AEROBICO
OSSIDAZIONE degli
ZUCCHERI - GRASSI
MODERATA
SUPERIORI
a 180”
PATTINAGGIO
Ghiaccio 500 mt. - Rotelle 300 mt.
NUOTO
800 mt., 1.500 mt.
CICLISMO SU STRADA, CANOA
AEROBICO
ANAEROBICO ALTERNATO
57
SPORT di SQUADRA – TENNIS - SQUASH
SISTEMI ENERGETICI
58
Riepilogo
Aerobico
Anaerobico Lattacido
Anaerobico Alattacido
Minima
Quasi Max
Max
Illimitata
15” – 45” * 45” –
180”
> 160
10” – 15”
Norm - > 180
da 2-3’ a giorni
da 0” a 120’
120 - 150
da 0” a giorni
59
UNITA’
MOTORIE
del
MUSCOLO
All'interno di ogni muscolo si riconoscono
diversi tipi di fibre, classificate in base alla
velocità
di
contrazione
e
alla
resistenza
alla
fatica.
60
Le UNITA' MOTORIE DEL MUSCOLO
sono fondamentalmente di due tipi (Figura 1 – Figura 2):
- Unità motorie lente: dette anche fibre rosse (prevalentemente aerobie e ricche
di mitocondri e mioglobina ) o ST o di tipo I.
- Unità motorie rapide: o fibre bianche (aerobie e anaerobie, povere di
mioglobina) o FT o di tipo II.
Le fibre a contrazione rapida si suddividono a loro volta in:
- fibre del tipo II A (o FTa) aerobie e anaerobie;
- fibre del tipo II B (o FTb) anaerobie.
In risposta ad uno sforzo
fisico intenso si attivano per
prime le unità motorie più
lente (FI) e, mano a mano che
l'intensità aumenta, si ha un
progressivo
Figura
1
61
reclutamento
delle fibre rapide F IIa – F IIb)
Entità della contrazione muscolare e tipo di fibre attivate
Figura 2
62
63
Sezione trasversale di biopsia muscolare, colorata con l'enzima ATP-asi acida (duecento
ingrandimenti). Si osservano fibre muscolari che reagiscono positivamente e vengono
identificate come fibre di tipo 1 (fibre scure) e fibre di tipo 2 che non si colorano
(fibre chiare). Le fibre muscolari che rigenerano presentano con questa reazione una
colorazione di tipo intermedio.
64
Quesiti
1) Quale e’ la minima unita’ contrattile del muscolo scheletrico?
2) Quali sono le proteine contrattili presenti nelle miofibrille?
3) Quali sono le proteine accessorie presenti nei miofilamenti?
4) Indicare le caratteristiche strutturali della Miosina. In quali filamenti e’ presente la Miosina?
5) Indicare le caratteristiche strutturali dell’Actina. In quali filamenti e’ presente l’Actina?
6) Quale e’ il ruolo svolto dalla Tropomiosina?
7) Quale e’ la sequenza di eventi che conduce alla contrazione muscolare?
8) Quale forma di energia usa il muscolo per produrre lavoro meccanico?
9) Quale e’ l’evento che determina il rilascio di ioni calcio dal reticolo sarcoplasmatico permettendo
l’inizio della contrazione muscolare? Quale e’ il mediatore che innesca l’accoppiamento
eccitazione-contrazione?
10) Quale e’ la funzione della fosfocreatina?
11) Quali sono le caratteristiche delle fibre muscolari rosse (lente)
12) Quali sono le caratteristiche delle fibre muscolari bianche (rapide)
13) Indicare I meccanismi energetici del muscolo in condizioni aerobiche ed in condizioni
anaerobiche.