PROF. GALDIERO - MICROBIOLOGIA - 26.03.2012 - SIMONA MARTONE Oggi parleremo un po’ velocemente di queste spore batteriche, non pare che le abbiate fatte
ancora. Fondamentalmente quando parliamo di spore parliamo di una forma di resistenza
all’ambiente. La cosa importante è che questo signore (slide?) dimostrò tanti anni fa che l’acqua
bollente uccideva le cellule vegetative di questo batterio ma non le endospore e in questo modo
confutò la teoria secondo la quale le cose potevano nascere dal niente etc etc perché anche se
l’acqua veniva bollita la possibilità che ci fosse dentro qualcosa che portasse poi alla crescita di
qualche batterio era possibile. Queste spore sono molto resistenti tant’è vero che sono state
descritte nelle mummie trovate tanti anni fa, monumenti antichi.. sono state trovate spore che in
opportune condizioni potevano dare crescita batterica. Dove si originano queste spore? Si trovano
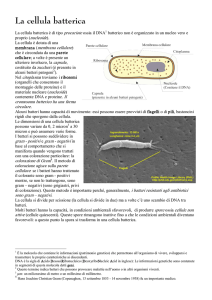
all’interno della cellula batterica. Questo qui è un batterio le spore si trovano direttamente
all’interno di questo batterio per questo sono denominate endospore. Si liberano nel momento in
cui questa cellula si disgrega e rimane questa pallina che rappresenta la nostra endospora e si
trova più o meno dappertutto. Chi può dare queste spore? Quali tipi di batteri possono dare queste
spore? Non tutti i batteri possono darle, fondamentalmente abbiamo (stiamo chiaramente parlando
di infestazioni di interesse medico) batteri gram+ che appartengono ai bacillus ed ai clostridium. Di
alcuni sicuramente avrete sentito parlare: il bacillus anthracis che causa il carbonchio. Allo stesso
gruppo appartengono altri che danno infezioni alimentari, respiratorie. E poi ci sono i clostridi di cui
sicuramente avrete sentito parlare, il clostridium botolinum, il clostridium tetani, gli altri sono meno
conosciuti ma ce ne sono vari altri. Quali sono le funzioni della spora? E’ molto resistente alla
penetrazione di qualsiasi sostanza estranea, resiste all’essiccamento e alle radiazioni, sopravvive al
calore anche oltre i 100°. Resiste per centinaia di anni nell’ambiente. Si possono eliminare? Sì. Ma
non attraverso una semplice sterilizzazione in autoclave (a 121° per 11 minuti a pressione
atmosferica le spore resistono). Allora c’è un’altra tecnica della tindalizzazione, da Tindal, che
prevede tre intervalli di 30 minuti seguiti da periodi di raffreddamento. Quindi si va da 100° poi si
ha un periodo di raffreddamento. Cosa succede? Tra i vari intervalli di 100°, tra il secondo ed il
terzo ciclo di bollitura il materiale viene tenuto a temperatura ambiente per almeno 8 ore. Significa
che noi prendiamo questo materiale, lo mettiamo a 100°, lo facciamo stare per un 30 minuti
dopodiché lo mettiamo a temperatura ambiente per un certo numero di ore. Cosa succede in
questo periodo? Precedentemente avevamo eliminato tutte le cellule vegetative, tutte le spore
rimangono lì. Però quando le mettiamo a temperatura ambiente queste spore cominciano a
germinare perché trovano delle condizioni favorevoli. Quindi se ci sono delle spore che
germineranno, nel secondo ciclo a 100° noi le andiamo uccidere e, per evitare che ne rimangano
altre, facciamo un terzo ciclo. Al termine noi abbiamo statisticamente un’altissima probabilità di
aver eliminato le spore. La cosa importante è che la spora è una forma dormiente, non si ha sintesi molecolare, non si ha
produzione di proteine, non fa niente. Le possiamo riconoscere anche all’osservazione. Nel genere
bacillus il diametro della spora non eccede quello della cellula batterica (viene definito sporangio,
che sarebbe la cellula batterica) che non si deforma e quindi abbiamo il bactrillio (?) ad esempio
qua: la spora è la pallina al centro. La forma del batterio è la stessa con o senza la spora. Nel caso
invece dei clostridi il diametro della spora è maggiore di quella della cellula che appare ingrossata
in corrispondenza della spora. Il clostridio vedete è allargato. Oppure abbiamo il vettrilio (?) se si
trova ad un’estremità. E’ facilmente evidenziabile all’interno della cellula con il microscopio ottico
perché colorata la spora appare come un corpicciolo rifrangente. Perché? Perché è molto resistente
anche alla penetrazione di coloranti e quindi non si colora e rimane traslucida. (illustra altre
immagini, anche al microscopio elettronico). Questa è una spora, con tutte le strutture fondamentali, all’interno c’è il citoplasma. Una ristretta
porzione di citoplasma, con materiale nucleare addensato vicino la membrana plasmatica, ovvero il
genoma batterico. Dopodiché c’è una parete rudimentale, perde una serie di strutture che noi
abbiamo visto nello studio dei batteri. Ha una serie di altre strutture che vengono definite come
cortex, piccoli residui della membrana plasmatica della cellula, un rivestimento interno, un
rivestimento esterno ed infine un ultimo rivestimento definito esosporio. Vedete che questo è
fondamentalmente costitiuto da una serie di rivestimenti che conservano il materiale genetico. Il
Scaricato da www.sunhope.it
core di questa struttura abbiamo visto cos’era. Ci sono alcuni ribosomi, alcuni RNAtransfer, non ci
sono RNAmessaggeri perché non c’è sintesi di macromolecole, ci sono piccole proteine solubili
SASP, non ci sono molti enzimi, c’è pochissima acqua (perde acqua, la prima cosa che fa) e
contiene un peptigoglicano modificato nella cortex, dove molti residui di acido N-acetilmuramico
sono sostituiti da anelli di acido lattamico (lattami muramici). E’ una struttura in grado di
mantenere la disidratazione di questa spora. Ancora più all’esterno abbiamo i due rivestimenti,
interno ed esterno, che corrispondono alla tunica sporale. Quindi un rivestimento interno che è
elettronlucido ed un rivestimento esterno che è elettrondenso. Che significa? Significa che diventa
scuro o chiaro a secondo delle colorazioni. Contiene diverse proteine similcheratiniche, molto stabili
e ricche di ponti disolfuro. E’ una struttura di natura fosfolipidica simile alla membrana plasmatica,
contiene basse concentrazioni acidi tecoici che ricordate si trovano nella parete dei gram+,
contiene acido diamminourmetico (? è sicuramente sbagliato, ma non riesco a capire) e
glucosammina. Cos’è che rende quest’endospora così resistente? Il calcio complessato con l’acido
dipicolinico. Questo è un acido particolare che si trova solo nelle spore. Ci sono delle proteine SASP
che si legano al core deidratato e rendono ancora più stabile questa struttura e poi vari
rivestimenti sporali. Come si chiama il processo? Sporulazione. Perché comincia? Normalmente
comincia se la crescita viene bloccata perché manca la cosa che serve a tutti i batteri, il
nutrimento. Vengono a mancare i nutrienti (frutti (?) di carbonio, azoto, fosforo) e quindi questa
cellula non è più in grado di replicarsi in maniera adeguata e se ne ha la possibilità, comincia la
sporulazione, che è un complesso processo formato da una serie di eventi. Quindi è una forma di
adattabilità di questa cellula batterica all’ambiente esterno: si adatta perché non è più in grado di
replicarsi. Infatti per esempio se volessimo farlo in laboratorio potremmo conservare queste spore
per molto tempo, continuando a mantenere questa mancanza di nutrienti, oppure porle in
condizioni favorevoli, causare una germinazione e queste cellule vegetative potrebbero essere
mantenute con l’ambiente adatto, le condizioni adatte, i nutrienti adatti a replicarsi continuamente.
L’intero processo dura circa 8 ore e può essere diviso in un certo numero di stadi. Qual è il primo
stadio: abbiamo una cellula, il DNA comincia ad addensarsi e, quello che vi ho spiegato un po’ di
tempo fa quando si cominciano e separare le molecole di DNA ecc ecc, si spostano in poli diversi
della cellula batterica. Si comincia a formare questo setto, sempre di più, ad un certo punto
comincia ad avere una diversa conformazione perché, mentre la cellula normalmente si
dividerebbe, in questo caso non ha più nutrienti, non può più aumentare a dismisura il numero
delle proteine ecc. quindi comincia a nascere un setto molto più spesso intorno a questa parte
(slide). Mano a mano il setto porterà alla formazione di questa corteccia, poi si farà la sintesi della
tunica, dell’esosporio, si ha la disidratazione di tutta la cellula batterica e quindi la cellula si
disgrega e libera la spora nell’ambiente. Al contrario, nel momento in cui ci siano condizioni
favorevoli, queste spore dalla loro condizione di spora in cui si possono ritrovare per periodi di
tempo più o meno lunghi, fa il percorso al contrario e quindi germina, liberando una cellula
batterica che può replicarsi. Quindi c’è un’attivazione, una germinazione ed un esocrescita. Quindi
l’attivazione prepara le spore per la loro germinazione (illustra la slide: questa è la spora, questo è
il batterio in forma vegetativa che sta uscendo dai suoi involucri, dopodiché sarà in grado di
replicarsi). Si ha un rigonfiamento in seguito a trattamento con il calore, si ha la rottura dei vari
rivestimenti della spora. Diminuisce la resistenza dei rivestimenti, aumenta l’attività metabolica e
una volta uscita comincia a replicarsi. La germinazione o trasformazione delle spore in cellule allo
stato germinativo richiede circa 90 minuti, è un processo abbastanza veloce, nel momento in cui
tutte le condizioni sono adeguate. L’induttore primario è uno stimolo traumatico tipo lo shock
termico che permette l’attivarsi di questi meccanismi. Fattori di germinazione sono la presenza di
questa serie di fattori, la L-alanina, l’aspargina, il fruttosio, il glucosio, ioni di calcio e magnesio ecc
ecc. che permettono la germinazione. Allora la perdita di frammenti degradabili di peptidoglicano,
di dipicolinato di calcio che era proprio una causa della resistenza di questa spora, viene ad essere
perso e si ha l’assunzione di acqua, quindi aumenta il volume, perde la resistenza al calore,
all’essiccamento, agli agenti chimici e quindi dopodiché può cominciare a crescere separata
completamente dai vari involucri che la circondavano. In vitro si può realizzare esponendo la spora
alla temperatura di 70° gradi per un’ora in presenza di zuccheri, amminoacidi, lisozima ecc. Quindi
Scaricato da www.sunhope.it
vuol dire che in laboratorio si può fare molto semplicemente. Quali sono i principali microrganismi in grado di dare spore? Abbiamo visto che appartengono ai
bacilli o ai clostridi. Quelli di interesse dal punto di vista medico sono il bacillus anthracis in grado
di dare l’antrace o il carbonchio, il clostridium tetani il tetano, il clostridim botulinum il botulismo e
clostridium perfringens la gangrena gassosa, quindi sono tutte patologie abbastanza serie,
importanti. Il clostridium tetani è un bacillo di piccole dimensioni, mobile, sporigino, acapsulato, gram+. Le
spore sono in posizione terminale, rotonde, quindi danno questa caratteristica forma a mazza di
tamburo. Cresce difficilmente in laboratorio perché essendo anaerobio non tollera la presenza di
ossigeno e quindi bisogna avere certe accortezze particolari. Si trova dappertutto, colonizza il tratto
gastroenterico degli erbivori e dell’uomo ed è presente nel terreno. La forma vegetativa è sensibile
alla presenza di ossigeno, la forma di spora può sopravvivere. È una patologia che ancora esiste,
contiene una tossina (e oggi vedremo proprio questa parte dei batteri, delle tossine che non
abbiamo ancora fatto. Subito dopo che abbiamo finito le spore, lo vedremo velocemente) si lega ai
recettori nervosi periferici, fa una serie di cose che riguardano le sinapsi inibitorie dei motoneuroni.
Blocca il rilascio di un mediatore chimico e questi motoneuroni che sono disinibiti perché non arriva
più il GABA, inviano impulsi continui per cui provocano il tetano che si manifesta con
manifestazione visiva: il paziente ha una paralisi spastica. I bacilli contengono un sacco di specie, poche sono associate a malattie dell’uomo, una di queste è
quella che viene chiamata bacillus anthracis. La maggior parte sono innocui, non sono neanche
studiati. Ce ne sono alcune di particolare importanza, ma fondamentalmente dovete ricordare il bacillus
anthracis. Sono gram+, sporigini, aerobi, rimangono anche questi nel terreno dove possono
resistere per moltissimi anni. Quelli associati a malattie sono il bacillus anthracis che causa il
carbonchio (il carbonchio può essere definito come cutaneo, da inalazione o gastrointestinale), poi
c’è bacillus cereus che causa infezioni intestinali, gastroenteriti, infezioni oculari, sepsi legate al
catetere, infezioni opportunistiche. Tutti gli altri danno infezioni opportunistiche che come dice
stesso il nome, in determinate condizioni, possono dare la patologia. Una caratteristica di questo
bacillo è che ha le estremità spigolose, sembra un rettangolino. Tende a crescere in filamenti, non
si separano i vari bacilli, restano legati per un certo periodo di tempo, vengono definiti a canna di
bambù. Che cosa fa esattamente? Il batterio produce queste endospore, qualche volta capita che
un erbivoro che mangi e queste si sviluppano all’interno dell’animale e possono portarlo a morte.
Nella carcassa dell’animale morto il batterio continua a riprodursi fino a quando non ha esaurito le
sostanze nutritive. A quello stadio il batterio produce nuove endospore che restano nel terreno e
vengono portate da una parte all’altra. Dico una fesseria qualsiasi, noi ci camminiamo sopra e le
spostiamo da una parte all’altra. La tossina del bacillo dell’antrace è formata da tre componenti che
sono antigenicamente distinte. La prima frazione è il fattore dell’edema, così definito, c’è un
adenilciclasi extracellulare, la terza frazione è una proteina tossica che corrisponde al fattore letale
e infine un fattore protettivo (se manca questa subunità le altre frazioni non riescono a
funzionare). La combinazione del fattore protettivo con una delle altre due componenti ha capacità
tossiche, quindi si capisce che il fattore protettivo necessita che ci siano le altre due e a seconda di
come si accoppiano portano un danno piuttosto che un altro. Esiste di questo batterio più di un
ceppo, almeno 89 ceppi. Differiscono per la presenza di diversi geni e a seconda di cosa
contengono hanno una diversa virulenza e produzione di antigeni e tossine. Alcuni di questi sono
noti, nel senso che dovremmo ricordare come si chiamano, ad esempio questo ceppo Ames che
era quello degli attacchi terroristici dell’inizio del 2000-2002; vollo novellum (? 31.55) anche questo
può essere utilizzato come arma chimica e pensate è stato isolato nel 1935 da una mucca, in
Inghilterra; altri ceppi come questo qui, lo Sterne, innocui, vengono utilizzati a scopo di ricerca. Le
spore possono sopravvivere per un sacco di tempo, il fattore di virulenza principale è la tossina del
carbonchio. Come si può trasmettere? Si può trasmettere per inoculazione o per inalazione.
Inoculazione vuol dire anche sulla pelle. Chi è a rischio? Chi la può prendere? Chi si trova a
contatto con animali infetti, potrebbe prenderla un veterinario che va a lavorare con la carcassa di
un animale morto o anche persone che lavorano con materiali di origine animale. Esiste una forma
Scaricato da www.sunhope.it
cutanea, una forma gastroenterica ed una forma inalatoria. Penso che non ci sia bisogno che ve lo
dico io, la più pericolosa è quella inalatoria, che porta a morte, le altre sono meno pericolose. Le
spore passano attraverso la cute, attraverso il tratto gastrointestinale o quello respiratorio. Si ha
germinazione dei macrofagi, trasporto ai linfonodi regionali, produzione locale di tossine, necrosi
od edema, batteriemia o tossinemia. L’antrace cutaneo è quello meno pericoloso, infatti ha una
mortalità abbastanza alta in pazienti non trattati, ma rara in caso di terapia antibiotica (esistono
antibiotici per trattare questo microrganismo); il batterio trattato adeguatamente viene eliminato
entro 24 ore e le lesioni scompaiono nel giro di una settimana. E’ chiaro che nel caso attacco
terroristico, se ci fosse il contagio di tutta la popolazione, come si farebbe a curarli tutti? Ecco la
mortalità. Però se è un caso, due o tre, uno ha sulla mano una schifezza immonda (...) si va a
curare. I sintomi: si forma una papula simile alla puntura di un insetto, pruriginosa. Dopo uno o
due giorni compare una vescicola.. (disquisizione sul puntatore che non funziona e che costa 50
euro). La vescicola si rompe, si trasforma in un’ulcera di 2-3 cm con un caratteristico centro di
colore nero (da qui anche il nome, nero come il carbone), quindi si nota molto che non è una
puntura di zanzara. La pustola non fa male, si accompagna ad edema e tumefazione dei linfonodi
regionali. Questo è un esempio (slide). Nel caso del contagio gastrointestinale, avviene per ingestione di carni infette poco cotte. e
caratterizzata da infiammazione acuta dell’intestino. Inizialmente si ha nausea, febbre, dolori
addominali, vomito, vomito ematico, diarrea ematica. La mortalità è più alta, anche con
trattamento antibiotico. E’ necessario un intervento tempestivo. Quella ancora più pericolosa è quella da inalazione che è rara. Il periodo di incubazione è vario a
seconda del numero delle spore inalate. Chiaramente è definito in base a studi su scimmie, quindi
non si sa esattamente quali sia, o tramite dati di incidenti. In questo posto, credo sia in Russia,
c’era un laboratorio dove si studiavano questi microrganismi, si ruppe un filtro nel 1979 e ci fu una
fuga di spore (il prof ridacchia), 77 casi da antrace di inalazione. Il tempo di incubazione fu di circa
10 giorni (i casi sono stati studiati molti anni dopo), anche se i primi casi ci furono dopo 2 giorni e
l’ultimo dopo 40 giorni. Allora, inalazione di particelle che trasportano migliaia di spore negli
alveoli, anche in questo caso intervengono i macrofagi; le spore possono essere portati fino ai
linfonodi; si trasformano, germinano in forme vegetative e possono causare malattia, si ha
formazione locale di tossine, edema o necrosi, batteriemia e tossinemia. I sintomi iniziali sono
febbre, astenia, tosse, l’inizio non è grave, non ce ne si accorge facilmente. Poi dispnea, meningite
nel 50% dei casi, shock e morte. Radiografia del torace rivela ingrandimento del mediastino con
pleurite senza infiltrati polmonari. Mortalità fino al 100% se la terapia antibiotica non è iniziata
immediatamente. Questa è la forma più grave. E’ la malattia del tosatore, sciocchezze varie, che
ha a che fare con gli animali. I primi sintomi sono simili ad un’influenza, sono sintomi piuttosto
comuni, quindi uno che non si immagina di aver preso l’antrace non si cura e le conseguenze
possono essere molto gravi. Nel periodo in cui c’era molta paura per le lettere minatorie che
arrivavano in America con le spore, se qualcuno le inalava, non mi chiedete come, ci pensava
molto di più. Se succedesse qui in Italia, nessuno ci penserebbe mai. Questa è una radiografia
(slide), va be’. A seconda della forma clinica uno va a prendere un campione diverso. Più o meno dovete solo
sapere che esistono vari campioni (lo farete meglio in diagnostica di laboratorio) e dove si trova il
microrganismo, perché io posso chiedervi cosa fa il microrganismo e dove si va a localizzare.
(Illustra una tabella) Nel caso di infezione cutanea si fa un prelievo sul sito della lesione e si manda in laboratorio dove
vengono fatte delle analisi. In questo caso la diagnosi è piuttosto facile. Nel caso gastrointestinale
si possono analizzare le feci, fare un tampone rettale (più o meno la stessa cosa). Si deve inviare
al laboratorio abbastanza presto. Oppure attraverso il sangue e in questo caso è in una fase più
avanzate perché si trova solo successivamente come batteriemia, sangue. Nel caso respiratorio la
prima cosa a cui pensiamo è l’espettorato, come per qualsiasi altra malattia respiratoria, ma non è
di grande utilità. Ancora una volta è importante il sangue, ma anche in questo caso la diagnosi si
può fare solo se la malattia è già avanzata. un’altra cosa che dovete sapere è quale farmaco agisce
su cosa, non specificatamente quanti grammi servono, ma almeno quale sia il farmaco. Il farmaco
Scaricato da www.sunhope.it
principale in questo caso è la penicillina, farmaco di elezione, l’eritromicina e il cloramfenicolo
anche sono utilizzati nel caso in cui non si possa utilizzare la penicillina. Per quanto riguarda gli
animali infetti, vengono abbattuti. Ogni volta che si utilizzano dei farmaci ci sono vantaggi e svantaggi. Non esiste nessun farmaco
che non abbia nessun effetto negativo. In questo caso conviene usare il medicinale? E’ certo.
Anche se in questo caso rischiamo di creare dei ceppi resistenti perché ogni volta che usiamo un
farmaco contro un microrganismo rischiamo di selezionare dei ceppi resistenti. Questo non
significa che non lo dobbiamo usare o usarlo assolutamente, lo dobbiamo usare correttamente e
quindi limitare la possibilità di creare resistenza. Esiste anche un vaccino preparato con delle
colture: si prendono queste colture di batteri, si fa una centrifugata e si prende il liquido che sta
sopra la provetta; in realtà questo è il terreno in cui sono cresciuti i microrganismi che contiene
degli antigeni protettivi di un ceppo (? Aggettivo che non ho capito) di bacillus anthracis. Viene
fatto il vaccino che funziona anche abbastanza bene, il 91% hanno anticorpi dopo la seconda dose
(dopo il richiamo). Ci sono altri batteri sporigeni di interesse medico: il bacillus cereus, anche in
questo caso contiene una serie di tossine, le spore possono sopravvivere nel terreno, la
trasmissione principale è mediante intossicazione alimentale, porta infezione oculare e quindi è
abbastanza importante. Può dare vomito nausea, crampi addominali. E poi ce ne sono altri
opportunisti elencati in questa tabella (slide). Mancano pochi minuti, ma cominciamo ad affrontare il discorso delle tossine. Cosa fanno questi
batteri quando incontrano una cellula ospite? Sono patogeni, hanno una certa patogenicità x.
Questa patogenicità è dovuta ad una serie di cose che sono intrinseci al microrganismo e possiamo
definire fattori di virulenza. Così come la carica batterica, se prendo un batterio e me lo mangio
non succede niente, se mangio una scatoletta di batteri, succede. L’altra cosa importante, e questa
è un fattore estraneo alla capacità del microrganismo, alla carica, e dipende dal nostro set up
generico e dal nostro stato di salute, principalmente a seconda del nostro stato immunitario, è un
fattore individuale: la stessa quantità dello stesso tipo di batterio può portare differenti
conseguenze tra due individui diversi perché siamo fatti diversamente. Non c’è una regola precisa. I batteri di per sé hanno una serie di proprietà che li rendono patogeni. Le cose fondamentali sono
tre: capacità di invadere, capacità di sviluppare un’azione tossica e riduzione di ipersensibilità o
fattori immunologici. Tutte queste capacità sono codificate a livello del genoma batterico, a volte
anche i fagi intervengono nella trasmissione di questi fattori. Una cosa molto interessante è
l’organizzazione genica in isole di patogenicità: a seconda della presenza o assenza di queste isole
di patogenicità un batterio può essere patogeno o non patogeno. Sono dei clusters particolarmente
importanti per la virulenza di questo organismo, se mancano non è patogeno. E’ importante perché
significa che questi geni patogeni sono stati trasferiti, sono derivati da altri microrganismi per una
serie di meccanismi.. avete fatto genetica batterica? No, va be’. Comunque con il passare del
tempo si sono accumulati in queste zone che sono state definite isole di patogenicità. Per esempio
nella tabella (slide) io la leggo, voi? Come non la leggete?? Va be’. Ad esempio E. Coli: c’è un
numero che corrisponde alla zona del genoma ove si trova, che chiaramente non è importante, e
contiene una serie di geni che codificano per una serie di cose importanti per la patogenicità dei
batteri. Ad esempio l’alpha-emolisina, le fimbrie, vari fattori di aderenza importanti nelle
infestazioni delle vie urinarie. Un altro esempio: stafilococco aureo ha SAP1 che contiene la
proteina dello shock tossico, è una enterotossina quindi contiene geni che fanno più patogeno il
microrganismo. Fondamentalmente dobbiamo essere esposti al patogeno, poi questo deve attaccarsi da qualche
parte per esplicare la sua azione patogena, deve aderire all’organismo, dopodiché si ha
colonizzazione e crescita e quindi aumento del numero di batteri, dopodiché ci sarà la
colonizzazione di altre cellule o il trasporto ad altri siti, tipo come dicevamo prima nel caso
dell’antrace: uno se le mangia e poi le può ritrovare nel sangue. La tossicità che può essere sia
locale che sistemica ed infine il danneggiamento dei tessuti e quindi il danno sui tessuti che
rappresenta quello che noi subiamo. Scaricato da www.sunhope.it
SCONOSCIUTA ( :/ ) Tratteremo tre argomenti generali: il metabolismo microbico la genetica batterica le colorazioni METABOLISMO MICROBICO Per quanto riguarda il metabolismo microbico voi siete sicuramente a conoscenza che tutte le
cellule sia eucariotiche, sia procariotiche, hanno bisogno di produrre energia. Questa viene estratta
da delle reazioni chimiche. Il metabolismo è l’insieme delle reazioni chimiche che avvengono negli
organismi. Si divide in anabolismo (sintesi di micro o macromolecole) e catabolismo (degradazione
di grandi molecole ad alto contenuto energetico). Normalmente i batteri si dividono in tre grandi gruppi che sono i: chemiorganotrofi,
chemioautotrofi e fotosintetici. La differenza fondamentale sta nel fatto che questi organismi possono utilizzare diverse fonti per
produrre energia. I chemiorganotrofi utilizzano composti organici, i chemioautotrofi utilizzano
sostanze che producono essi stessi e i fotosintetici la luce. In questo schema (slide) voi riuscite già a vedere la differenza fra i chemioautotrofi e i
chemioeterotrofi. Come composto organico i chemioautotrofi utilizzano la CO2, la autoproducono,
mentre i chemiorganotrofi utilizzano comporti organici introdotti nella cellula attraverso meccanismi
di endocitosi o trasporto passivo. I fotoautotrofi li raggruppiamo insieme con i fotosintetici, la
forma di energia è la luce. Qual è la molecola che utilizzano come moneta di scambio di energia?
Ovviamente è l’ATP. La forza di questa molecola sta nel fatto che nei legami in cui sono racchiusi i
gruppi fosfato, è racchiusa una forza molto elevata. Tutto quello che non può essere utilizzato dalla
cellula viene dissipato sotto forma di calore e quindi viene utilizzato per mantenere la temperatura.
Ci sono due meccanismi nei batteri ma non prescindono l’uno dall’altro, non è detto che il batterio
che sia in grado di effettuare la respirazione non sia anche in grado di fermentare. Tutto dipende
dallo status e l’habitat in cui si trova. La respirazione è il meccanismo che prevede la scissione del glucosio, la formazione di 2 molecole
di piruvato ed infine il trasporto degli elettroni e il ricavo di energia netta che di solito avviene a
carico di una membrana. Come sapete negli organismi superiori avviene nei mitocondri, nei batteri
a carico della membrana cellulare. La fermentazione invece prevede la formazione di molecole di
piruvato però non esiste la catena di trasporto degli elettroni, e quindi in assenza di ossigeno,
queste molecole danno origine ad altre molecole con ricavo di energia minore. La respirazione è
più conveniente. Oltre all’ATP ci sono altre molecole con legami ad alta energia, abbiamo già introdotto il glucosio
che ha 6 atomi di carbonio, viene scisso attraverso la glicolisi in due molecole di piruvato da 3
molecole di carbonio l’uno, poi c’è ancora l’acetilCoA e l’acetilfosfato. Sono tutti intermedi di queste
reazioni di scissione del glucosio. Parliamo della glicolisi che è appunto questo meccanismo attraverso cui il glucosio viene
trasformato in piruvato. Il glucosio viene introdotto attraverso trasportatori, endocitosi o trasporto
passivo (semplicemente perché nell’ambiente esterno c’è una maggiore concentrazione che
all’interno) e subito viene trasformato in glucosio-6-fosfato. E’ una reazione importantissima,
mediata da un enzima chiamato esochinasi. In realtà è una reazione limitante perché nel momento
in cui il glucosio si trasforma subito il G6P, automaticamente abbassa la concentrazione
intracellulare di glucosio. Ciò fa si che nella diffusione passiva di glucosio, in tutti i casi, verrà
mantenuta una concentrazione interna bassissima, quindi il glucosio entra nella cellula ma la sua
concentrazione rimane bassissima perché viene subito trasformato. A carico poi di altre due
trasformazioni a carico di altri due enzimi che sono l’isomerasi e la fosfofruttochinasi, c’è la finale
trasformazione in fruttosio-1,6-difosfato. Questa prima fase è detta fase di investimento perché
come potete vedere, la prima e la terza reazione avvengono con consumo di due molecole di ATP
(che diventa ADP). Lo stesso ADP passa nella fase due che è la fase di ossidazione. Inizia quindi la
Scaricato da www.sunhope.it
trasformazione delle molecole a 6 atomi di carbonio in molecole a 3 atomi di carbonio: avremo due
molecole di gliceraldeide-3-fosfato, successivamente 2 molecole di 1,2-difosfoglicerato ed infine
due molecole di 3-fosfoglicerato. Tutto questo attraverso il consumo di elettroni. Queste molecole
perdono atomi di H che vanno a loro volta a ridurre i coenzimi che sono il NAD+ che diventa NADH
ed infine con produzione di due molecole di ATP nell’ultima reazione. Il bilancio finale di questa
prima fare della respirazione batterica è la produzione di 2 molecole di NADH, poiché le 2 molecole
di ATP spese all’inizio vengono recuperate. Le reazioni controllo sono quelle che permettono in
questa via metabolica di inibire la stessa reazione, abbiamo la esochinasi, la fosfofruttochinasi e la
piruvatochinasi che, onde evitare l’accumulo eccessivo di piruvato, che può essere dannoso per la
cellula, sono autoindotte da elevate concentrazioni del prodotto della reazione. Quindi la esochinasi
è inibita da alte concentrazioni di glucosio-6-fosfato, la fosfofruttochinasi è inibita da ATP e citrato
e la piruvatochinasi da elevate concentrazioni di ATP. Nel caso in cui non avessimo a che fare con un organismo in grado di ossidare il piruvato, che non
è in grado di respirare, che non ha una catena di trasporto di elettroni e che quindi effettua la
fermentazione, le molecole di piruvato vengono trasformate in due molecole di lattato attraverso,
appunto, la fermentazione. Nella fattispecie se il batterio attraverso una lattato deidrogenasi, con
ossidazione del NADH che diventa NAD+ e conseguente riduzione del substrato, trasforma il
piruvato in lattato. Allo stesso modo se il batterio è dotato di una alcol deidrogenasi, il piruvato
darà origine ad etanolo. Ancora, se è in grado di fermentare il piruvato attraverso un altro processo
avremo la formazione di acetato e infine H2 + CO2. Questi processi li trovate normalmente in natura e negli alimenti, nella fattispecie la lattato
deidrogenasi la trovate nella produzione di yogurt con formazione di lattato. L’etanolo invece si
forma nella produzione di alcolici e vini in particolare. Se ci troviamo di fronte ad un batterio fermentante, il substrato viene parzialmente ossidato, il
NAD+ si riduce a NADH grazie agli idrogeni sottratti al piruvato (questo eccesso di NADH alla fine
sarà un fattore limitante della reazione). L’ATP invece sarà prodotto grazie a reazioni di
fosforilazione a livello del substrato. La fermentazione ha una resa energetica estremamente bassa
perché il piruvato ed i successivi prodotti comunque sono molecole a basso contenuto energetico:
producono pochissima ATP e non contengono legami con il fosfato. Ricapitolando i prodotti della fermentazione: abbiamo quella omolattica, che dà come prodotto
finale acido lattico, a carico del microrganismo lactobacillus species (tutte le specie del
lactobacillo); la fermentazione alcolica produrrà etanolo, a carico dei lieviti; la fermentazione acidomista, in cui vengono prodotti una serie di acidi (etanolo, acetato, acetoino)
e questi sono a carico klebsiella o enterobatteri. Fra l’altro la fermentazione acidomista e la
quantizzazione di questi acidi vengono utilizzati nelle analisi microbiologiche delle acque, per
vedere effettivamente la carica dei coliformi fecali che sono presenti all’interno dei reflui. Negli organismi che non sono fermentanti o che possono fermentare ma sono anche aerobi, dopo
la formazione di piruvato e quindi la prima scissione del glucosio, generalmente sussegue il ciclo di
krebs. Nel ciclo il piruvato entra a partire dalla glicolisi e viene subito trasformato in AcetilCoa. Da
qui c’è questo ciclo in cui ci sono tantissimi intermedi (citrato, isocitrato, alpha-chetoglutarato ecc)
che attraverso una serie di reazioni di ossidazione del substrato, porteranno alla fine alla
formazione di acido ossalacetico. Così di nuovo può ricominciare il ciclo con condensazione di acido
ossalacetico e acetilCoA. La funzione principale del ciclo è di produrre ATP, di ridurre il NAD+ a NADH ed infine di ossidare il
substrato. La maggior parte degli intermedi sono estremamente importanti per altre reazioni di
biosintesi. Come vedete, infatti, il citrato entra nella via di biosintesi degli acidi grassi o del
colesterolo; l’alpha-chetoglutarato fa parte della biosintesi e della ossidazione degli amminoacidi.
Quindi entrano nelle vie di biosintesi di molecole fondamentalmente importanti per l’attività della
cellula stessa. Il succinilCoA fa parte delle sintesi delle porfirine; il fumarato fa parte della via di
biosintesi di aspartato, tirosina e fenilanalina che conoscete benissimo come amminoacidi; il
malato entra nella via di biosintesi del glucosio ovvero della gluconeogenesi. Questo significa che
attraverso la reazione della glicolisi ed il ciclo di krebs, la cellula istaura un equilibrio metabolico
Scaricato da www.sunhope.it
attraverso cui riesce a terminare il ciclo di krebs se ha bisogno di energia, oppure a produrre questi
intermedi per le vie di biosintesi. Questa è una scelta che la cellula fa ogni volta che ha bisogno di
intermedi o energia. Il bilancio finale del ciclo di krebs abbiamo 1 Acetil-CoA, 3 molecole di NADH, 2 molecola di FADH2
e ATP e CO2. La produzione massima di energia sarà a carico dell’ossidazione del substrato e della
catena respiratoria. Vediamo questa catena respiratoria a carico di una membrana. Come già ho detto prima negli
organismi superiori avviene nel mitocondrio, nei procarioti sulla membrana citoplalsmatica. A
partire dall’alto abbiamo 4 complessi proteici che hanno il ruolo di trasportare gli elettroni
dall’ambiente esterno all’ambiente interno. L’ambiente esterno è già acidificato, c’è una grande
concentrazione di elettroni a carico della membra esterna, mentre sul versante interno c’è già un
ambiente scarico di elettroni. Qual è il ruolo di questa catena respiratoria? Attraverso questi
complessi gli elettroni vengono portati fuori e questo avviene con un minimo spreco di energia e
soprattutto avviene a carico di una pompa (slide) che è l’ATP sintetasi. In pratica l’energia che
viene sfruttata dagli ioni H+, che per diffusione sono portati all’interno del citoplasma, viene
sfruttata dalla ATP sintetasi per produrre energia. L’energia del trasporto, quindi, viene sfruttata
per legate un gruppo fosfato all’ADP che diventa ATP. Questo enzima è sempre legato alla
membrana è costituito da 3 subunità alpha, 2 subunità beta ed infine 1 gamma, una epsilon ed
una delta. E’ molto simile nei procarioti ed eucarioti ed ha la semplicissima funzione di sfruttare
l’energia di questi H+, che normalmente sono spinti ad entrare nella cellula, per formare ATP. I
complessi sono tutti proteici e prendono il nome della proteina stessa. Il primo complesso è
formato da una flavoproteina, il secondo è formato dal chinone ed infine ci sono questi altri due
che sono costituiti da proteine ferro-zolfo e quindi semplici trasportatori di elettroni. Per quanto
riguarda le esigenze gassose di questi microrganismi, semplicemente, gli aerobi sono capaci di
effettuare la respirazione e non sono generalmente fermentanti a meno che non vi sia una carenza
di ossigeno; gli anaerobi crescono in assenza di ossigeno e sono fermentanti; gli anaerobi
facoltativi possono vivere anche in assenza di ossigeno. Infine i microaerobi capaci di vivere in
presenza di bassissime concetrazioni di ossigeno (anche 2%). Qual è il bilancio finale della respirazione aerobia? 38 molecole di ATP per ogni molecola di
glucosio. Nella respirazione anaerobia abbiamo la fermentazione, la formazione di altri intermedi,
dove l’accettore finale può essere il nitrato (nitrobatteri), lo zolfo, il solfato (solfobatteri), il
fumarato, il NAD o la CO2 (metanobatteri). C’è una bassissima produzione di energia, circa 10 ATP. Adesso parliamo di fotosintesi. Ci sono alcuni organismi come le alghe ed i cianobatteri che non
hanno né una respirazione aerobia, né la fermentazione. Questi si avvalgono di una catena di
trasporto degli elettroni che è molto simile a quella che abbiamo visto prima, sempre a carico di
una membrana: la membrana dei cloroplasti. Nella fattispecie dei tilacoidi che sono quei serbatoi
impilati che si trovano all’interno del cloroplasto stesso. In questo caso vengono assorbite le
radiazioni luminose sottoforma di fotoni. Il fotone risulta come il nostro H+ precedente e come
potete vedere si ha formazione di ATP. I complessi sono praticamente identici e alla fine esiste
sempre la nostra ATP sintetasi che sfrutta l’energia dell’H+ che entra all’interno del tilacoide per
produrre ATP e quindi fosforilare il substrato. Questi 3 complessi Bs2 B6s e Bs1 (1.31.55), sono costituiti da altre proteine a loro volta. LA GENETICA BATTERICA Tenete presente che i batteri sono gli organismi che hanno una ricombinazione estremamente
elevata del proprio genoma, molto più degli eucarioti. Questo perché i batteri, esistendo come
cellule singole, sono più esposti a determinati eventi. In particolar modo: all’ingresso di DNA
eterologo attraverso semplicemente degli accettori che si trovano sulla membrana; attraverso
l’infezione da parte di virus e quindi l’integrazione del DNA o RNA virale; infine tramite un processo
tipico dei batteri chiamato coniugazione. Coniugazione: due cellule batteriche entrano in contatto attraverso una struttura chiamata pilus
che permette il trasferimento di materiale genetico, in particolar modo di plasmidi (cromosomi
circolarizzati che non sono integrati all’interno del DNA batterico, ma esistono liberi all’interno del
Scaricato da www.sunhope.it
citoplasma o del nucleo). Il pilus è un ponte fibroso che permette il passaggio fisico del DNA di un
batterio al DNA di un altro batterio. La trasformazione: passaggio di frammenti di DNA libero, originati dalla lisi batterica, ad un
batterio ricevente (DNA eterologo) attraverso la membrana plasmatica. Trasduzione o conversione fagica: il trasferimento genetico è mediato dai batteriofagi (virus in
grado di infettare i batteri). Il virus inietta il suo patrimonio genetico che se è DNA può integrarsi
nel genoma batterico. (illustra slide) Parliamo della coniugazione. Questi pili sono presenti in genere da uno a dieci per cellula, lo
spessore è di 9-10 nm ed ogni singolo batterio ha il gene per la produzione di questo pilus e se
non ne è provvisto può essere solo un batterio ricevente. Come funziona? Vedete che il
cromosoma batterico si trova all’interno della cellula donatrice, chiamata cellula F+, insieme ad un
plasmide che è il plasmide F, il gene che codifica per la produzione del pilus. Quindi, quando il
nostro donatore sarà in grado di produrre questo pilus, semplicemente aggancerà la cellula
ricevente (F-). A questo punto le due cellule si avvicinano, il pilus si accorcia, e quando la coppia si
stabilizza inizia il trasferimento. Il trasferimento non è a carico di tutto il plasmide (costituito da
due eliche) ma di una singola elica. A carico del donatore infatti esiste un’elicasi che provvede alla
rottura di questa doppia elica; dal lato del ricevente esiste una DNA polimerasi che si occupa della
duplicazione della singola elica in modo che alla fine del trasferimento la cellula F+ avrà dato alla
cellula F- un plasmide già sotto forma di doppia elica. Alla fine avremo due cellule F+ entrambe in
grado di produrre il pilus. Questo trasferimento può essere interrotto per qualsiasi meccanismo,
anche un semplice movimento (perché le due cellule sono passive nel substrato) e così non tutte le
coniugazioni hanno buon termine. Questo è solo un sistema modello. Esiste un altro gene HFR
(high frequence of ricombination), tutti i batteri che hanno un HFR hanno un’elevata capacità di
produrre questi pilus, sono capaci di trasferirlo ai riceventi. Idealmente, al termine della
coniugazione avremo un F- che è diventato F+ ed ha anche il gene HFR. La coniugazione ha
bisogno di tre particolari eventi per poter avvenire: uno stretto contatto cellula-cellula, un plasmide
coniugativo (F+) e geni per il trasferimento e la sintesi del pilus stesso (elicasi). I plasmidi sono
materiali extracromosomiali, circolari, con dimensioni variabili ma molto più piccoli del DNA
batterico, presenti in centinaia di copie, non hanno informazioni genetiche fondamentali
(semplicemente conferiscono un vantaggio selettivo alle singole cellule) e sono importanti per la
coniugazione. Possono avere anche un’altra funzione, ovvero possono essere portatori, possono
essere portatori di virulenza e malattie. Es. ceppi enterotossici di E. Coli che presentano delle
tossine, le quali sono plasmidi che fuoriescono dal batterio una volta che si trova nell’intestino. Le
tossine possono essere termolabili o termostabili a seconda della loro stabilità al calore e sono
tutte portati dai plasmidi. Quindi ceppi enterotossici di E. Coli diventano tali solo se sono portatori
di plasmidi. Ci sono anche altri batteri che portano plasmidi virulenti: la tossina tetanica del
clostridium tetani e la tossina del bacillus anthracis. Per quanto riguarda invece la trasformazione che normalmente viene anche fatta anche in
laboratorio ed è un evento abbastanza frequente, abbiamo già spiegato che succede che i batteri
vadano incontro a lisi dopo un infezione con un virus oppure poiché vanno in apoptosi perché non
sono presenti nutrienti a sufficienza. In una colonia batterica quindi diversi batteri possono andare
incontro alla rottura della cellula stessa. Nell’ambiente extracellulare si riversa tutto il contenuto
della cellula appena lisata, materiale genetico. Se questo materiale genetico riesce ad entrare
all’interno di un’altra cellula batterica, esso può ricombinarsi con il DNA della cellula ricevente.
Questo DNA viene chiamato eterologo. Il processo viene anche chiamato trasformazione perché la
cellula finale è detta trasformata. Ovviamente sulla membrana del batterio devono essere presenti
determinati enzimi, come le nucleasi, le quali sono capaci di permettere l’ingresso del DNA
eterologo. A dispetto della coniugazione, non richiede uno stretto contatto tra le cellule, utilizza
DNA libero e non DNA che si trova già all’interno dei batteri, è tipica dei batteri del suolo (perché
sapete che nel suolo ci sono moltissimi batteri ed è molto più facile che avvenga la lisi dei batteri).
Le cellule che sono in grado di captare il DNA libero sono dette competenti. Peraltro le cellule
possono essere rese competenti anche a livello di laboratorio. Queste cellule possono essere rese
Scaricato da www.sunhope.it
tali attraverso l’elettroporazione: la cellula batterica viene stimolata con basse scariche elettriche e
sulla membrana si formano dei pori che permetteranno l’ingresso del DNA eterologo. La cellula può
essere stimolata anche attraverso CaCO2, carbonato di calcio, che comporta ugualmente la
formazione di pori alla membrana batterica. Questi meccanismi porteranno all’entrata di DNA
eterologo anche in batteri sprovvisti di nucleasi sulla membrana. Come si fa a capire se questo
DNA trasforma o meno la cellula? Semplicemente vedendo se si riscontra l’espressione di questo
gene, normalmente si utilizzano dei geni che la cellula non porta (geni che esprimono degli
enzimi). Questo processo di trasformazione viene utilizzato per acquisire nuove caratteristiche
genetica, per riparare dei geni danneggiati, addirittura per nutrimento della cellula, il DNA viene
degradato ed utilizzato come nutrimento. Infine c’è la traduzione. E’ un processo che avviene frequentemente in natura, attraverso il quale
dei batteriofagi (virus capaci di infettare un batterio), iniettano il proprio DNA all’interno del
batterio e questo riesce a ricombinarsi a livello del DNA batterico. Questo è un tipico fago (slide),
all’interno della testa esiste il DNA, poi c’è un corpo che ha forma di spirale ed infine esistono
queste fimbrie che hanno la capacità di interagire con la membrana plasmatica ed agganciarsi al
batterio. Dopo questo primo momento di attacco del virus, il virus modifica la proprio
conformazione ed inietta (come una siringa) il proprio DNA all’interno del batterio. Nel caso in cui
abbiamo a che fare con batteriofagi virulenti, il primo ed unico ciclo che riescono ad attuare è il
ciclo litico. Una volta che è stato iniettato il DNA virale, questo viene utilizzato dalla cellula
batterica per produrre progenie virale (identica al virus che la ha infettata). Il virus non è altro che
un parassita che utilizza le strutture del batterio, anche gli enzimi (DNA polimerasi), i ribosomi, per
poter produrre le proteine strutturali e non che gli servono per poter dare origine a questa
progenie virale. Oltre al ciclo litico esiste anche il ciclo lisogenico. Questo è molto più semplice.
Risulta nei batteriofagi temperati, dove questo DNA fagico è capace di integrarsi all’interno della
cellula batterica e rimanere silente per molto tempo (non quantificabile). A seguito di un evento,
chiamato induzione, automaticamente questo DNA che si era prima integrato all’interno del
cromosoma batterico, subisce un attivazione. Vengono tradotti i geni, si trasformano in mRNa e poi
in proteine strutturali e non strutturali. In seguito ad un periodo, dunque, chiamato silente
finalmente si ha la produzione di progenie virale che sarà talmente numerosa che uscirà dalla
cellula solo quando essa sarà veramente piena andando, in tal modo, a lisare la cellula batterica.
Le cellule a quel punto diventano veramente gonfie e vanno incontro a rottura. Distinguiamo la traduzione generalizzata o specializzata, sempre dipendentemente dal virus. È
generalizzata quando il batteriofago indipendentemente dal batterio che si trova davanti può
andare incontro al un ciclo litico o lisogenico; la specializzata avviene quando un batteriofago nella
fattispecie può infettare soltanto alcuni tipi di batteri (E. Coli o bacillus subtilis) e in questo caso
avremo un ciclo litico o lisogenico specificatamente a carico di quel batteriofago e di quella cellula
batterica. COLORAZIONI Per quanto riguarda l’identificazione dei microrganismi, noi faremo l’identificazione non biochimica
(perché vi è già stata spiegata). Noi faremo una parte abbastanza divertente che è l’identificazione
dei microrganismi attraverso le colorazioni. Allora le colorazioni in microbiologia vengono effettuate perché i batteri sono trasparenti e si vanno
ad uniformare con il piano del vetrino. Mentre le andiamo sicuramente ad identificare perché i
contorni si riescono a vedere, in grigiastro o comunque in una colorazione molto più scura,
comunque i coloranti servono ad evidenziare le cellule batteriche, per creare un forte contrasto tra
cellula e vetrino, ma anche per evidenziare i diversi caratteri morfologici perché altrimenti non
saremmo in grado di farlo. Tra l’altro esiste una colorazione che addirittura è in grado di
differenziare i gram+ e i gram- (colorazione di Gram). Un altro motivo per cui andiamo a colorare è
per evidenziare una serie di strutture: flagelli, capsule, endospore. Esistono infatti colorazioni che
ci permettono di colorare in maniera diversa le differenti strutture, sia intracellulari che
extracellulari. Questo ovviamente ci consente di riuscire a vedere nel migliore dei modi il
microrganismo. Scaricato da www.sunhope.it
Le colorazioni vengono fatte attraverso coloranti divisi in acidi e basici. I basici sono dotati di carica
+ e si legano a strutture acide (superfici cellulari, proteine, acidi nucleici). I coloranti basici sono:
blu di metilene, fuxina basica, violetto di genziana, cristal violetto o tionina. Gli acidi, viceversa
hanno carica – e si legano a strutture basiche (attorno al microrganismo). Sono: eosina,
negrosina? e rosso fuoco. I più utilizzati sono sicuramente l’eosina per i coloranti acidi ed il blu di
metilene, il cristal violetto o la fuxina basica per i coloranti basici. Le colorazioni sono normalmente semplici o differenziali. Le semplici quando utilizziamo un solo
tipo di colorante, normalmente basico. E’ detta colorazione diretta ed è effettuata normalmente
con il blu di metilene o la fuxina. Il colorante viene applicato al campione, precedentemente fissato
con la formaldeide (formalina), per un tempo variabile. L’eccesso di colorante è poi eliminato con
acqua ed il vetrino è osservato al microscopio. Questo permette di osservare la morfologia e
l’organizzazione cellulare, poiché tutti i microrganelli presenti all’interno della cellula verranno
colorati in maniera indifferenziata. Quando invece facciamo una colorazione differenziale, andiamo ad utilizzare due o più coloranti ed
altri reagenti. Oltre alla formaldeide, inoltre, si utilizzano dei mordenzanti per il secondo coloranti o
dei decoloranti per poter permettere al secondo colorante di penetrare in quelle strutture dove il
primo non è riuscito a legarsi. Questa permettere di distinguere le diverse tipologie di
microrganismo (gram+ o gram-) e le diverse strutture dello stesso microrganismo (distinguiamo la
membrana, il nucleo, il citoplasma, il Golgi). Andremo a vedere: la colorazione di Gram, la colorazione per i bacilli acido-resistenti (ziehl-neelsen
, la colorazione della capsula, dei flagelli, delle spore, dei villi, dei funghi e delle spirochete. Per quanto riguarda la colorazione semplice, prima di passare alle differenziali, è quella che
abbiamo già anticipato con la fuxina, il blu di metilene, il cristal violetto. Come avviene con il blu di
metilene (ma anche con tanti altri)? Il vetrino viene poggiato sulla vaschetta con la colorazione o
semplicemente su dei sostegni che si trovano all’interno del lavello. Viene aggiunto il blu di
metilene e si attendono 3 minuti circa. Il campione è già stato fissato prima con la formaldeide.
Dopo 3 minuti con acqua viene lavato il campione, viene fatto asciugare e poi viene visto al
microscopio. Si vedono in risalto, sul vetrino, tutti i batteri colorati, in maniera indifferenziata. Se volessimo avere, invece, una colorazione differenziale, utilizzeremo la colorazione di Gram. Si
applica il cristal violetto sul campione già fissato. Dopo il fissaggio si applica il mordenzante. Dopo
il lavaggio con alcol avverrà una decolorazione. Nella prima fase il colorante è penetrato all’interno
di tutte le strutture batteriche, in maniera indifferenziate. Applicando il mordenzante e poi l’alcol, io
riesco a decolorare alcune strutture ove il cristal violetto non è riuscito a penetrare. In realtà il
cristal violetto colora tutto, anche le strutture basiche, ma non con la giusta affinità di carica, così
che, quando metto l’alcol, vado a decolorare le strutture basiche con cui il cristal violetto non è
riuscito a legarsi. A questo punto aggiungo la safranina 2.02.25 che è di solito rossa ed ha
un’affinità per le strutture basiche ed avrò una colorazione differenziale delle strutture acide e
basiche. I betteri gram+ appariranno blu, i gram- rossi. Riusciamo a vedere perché avviene questa
differenziazione. I batteri gram+ vengono colorati con il cristal violetto. Quando aggiungo il
mordenzante (a base di iodio), si forma questo grande complesso che precipita all’interno della
cellula. Una volta precipitato rimane all’interno della cellula (gram+) perché nei gram+ noi
abbiamo uno struttura, chiamata peptidoglicano, che non fa altro che intrappolare questo
complesso (cristal violetto + iodio). Nei gram- ciò non avviene poiché il peptidoglicano esiste ma il
batterio ha due membrane e quindi il colorante stazionerà inizialmente tra peptidoglicano e
membrana esterna. Quando noi andiamo ad aggiungere l’alcol, la membrana si dissolve e permette
a questi complessi di fuoriuscire. Di conseguenza, quando io aggiungo invece l’eosina, la safranina
(affine a strutture basiche) questa penetrerà all’interno del peptidoglicano e colorerà i gram-. Il
risultato finale sarà che i gram+ appariranno blu grazie al complesso violetto-iodio, mentre i gramperdono questo complesso e si coloreranno di rosso. Non tutti i batteri possono essere colorati con la colorazione Gram. I micobatteri, a causa della loro
struttura cerosa, sono impermeabili a questi coloranti. Viene utilizzata la colorazione di ziehlneelsen. Qual è il problema fondamentale? Sono caratterizzati sulla superficie cellulare da ceramidi
e fosfolipidi, quindi la colorazione diretta o selettiva, non riescono a penetrare la membrana. Per i
Scaricato da www.sunhope.it
batteri, chiamati acido resistenti, vengono utilizzate queste due colorazioni –un (?) e ziehl-neelsen.
Nel primo step si versa la fuxina basica sul nostro campione. Nel secondo step si fa evaporare la
fuxina riscaldando il campione con una fiamma per 5 minuti. Successivamente il campione viene
lavato con acqua. A questo punto bisogna decolorare con alcol acido fino a quando il colorante non
scompaia dal vetrino (ma non da alcune strutture che non sono acido resistenti che saranno state
colorate). Dopo un lavaggio si può contrastare con il blu di metilene ed infine si lava con acqua. Gli
organismi acido resistenti sono colorati in rosso, quelli non acido resistenti sono colorati in blu.
Qual è la differenza? Gli organismi acido resistenti (rossi) sono tali perché attraverso il
riscaldamento del campione essi sono riusciti a penetrare attraverso le membrane cerose. Grazie al
calore i microrganismi sono comunque penetrati da questo colorante rosso. Quando poi lo andiamo
a colorare, noi effettuiamo una semplice colorazione diretta, e tutti quelli che non sono acido
resistenti risulteranno colorati in blu. I passaggi fondamentali sono lo step 1 e lo step 2
(riscaldamento), il resto è una semplice colorazione diretta. Infine c’è una parte dedicata alle colorazioni speciali. Una colorazione chiamata Negativa, dove
normalmente l’organismo non viene colorato, ma viene colorato tutto quello che c’è intorno (come
il negativo della fotografia), di conseguenza risulteranno tutte le strutture batteriche proprio
perché non sono colorate. Un’altra colorazione speciale è quella delle spore. Come i batteri acido resistenti, richiede calore
per facilitare l’ingresso del colorante nella spora, perché, come sapete, la spora ha una membrana
estremamente impermeabile costituita da sostanze cerose. Infine esiste la colorazione per i flagelli. Anche in questo caso, come per la colorazione dei gram, si
utilizza un mordenzante per irrobustire la struttura flagellare. Quindi con formazione di un
complesso che precipita ed ispessisce il flagello. Scaricato da www.sunhope.it