Il reticolo endoplasmatico (RE) è una
struttura cellulare costituita da regioni
citoplasmatiche delimitate da membrana
che possono avere la forma di cisterne o
tubuli. Lo spazio interno che si identifica
tra le pieghe del reticolo è detto lume e
presenta una serie di enzimi che
catalizzano diverse reazioni chimiche.
Vi sono due tipologie di RE differenti per
morfologia e funzione e il cui prevalere
dell'uno sull'altro dipende dalla tipologia
cellulare di cui fa parte:
il reticolo endoplasmatico rugoso (RER o
REG)
e liscio (REL).
Il reticolo endoplasmatico rugoso è un organello
della cellula eucariote sia vegetale che animale;
è costituito da una serie di membrane piegate
una sull'altra a formare cisterne, mentre il
termine rugoso (o ruvido) si riferisce al fatto che
il versante citoplasmatico delle sue membrane è
punteggiato di ribosomi. Compito del ribosoma è
quello di sintetizzare proteine tramite la "sintesi
proteica". Una volta ottenuta la proteina, questa
viene impacchettata in una membrana
(gemmazione della vescicola di transizione) che,
trovatasi nel citoplasma può fuoriuscire dalla
cellula (esocitosi) o sostare nel citoplasma o
andare nell'apparato di Golgi dove verrà
modificata. Tutti i movimenti delle vescicole
possono essere compiuti tramite il
coordinamento del citoscheletro.
Reticolo endoplasmatico liscio
Il Rel è costituito da un sistema di sacche
tubolari. È il maggior responsabile della sintesi
dei lipidi, degli ormoni steroidei e del
metabolismo del glicogeno. Ha come compito
quello di detossificare sostanze altrimenti
dannose per l'organismo, come ad esempio
l'etanolo contenuto nelle bevande alcoliche. Per
questo motivo ritroveremo una rigogliosa
presenza di Rel in cellule epatiche. Il Rel inoltre
è sede primaria del metabolismo di fosfolipidi,
acidi grassi e steroidi, serve a sintetizzare i
lipidi utili per costruire e riparare tutte le
membrane della cellula.
Il REL ha anche la funzione di
immagazzinare ioni calcio. Nel tessuto
mscolare, questi ioni sono necessari per la
contrazione; quando un impulso nervoso
stimola una cellula muscolare, gli ioni
calcio passano dal reticolo liscio nel fluido
citoplasmatico, dove innescano la
contrazione della cellula.
RER in cellule di fegato umane
Golgi Complex
Also called Golgi body or Golgi apparatus
This is a curved membrane stack,
resembling a stack of pancakes
It packages proteins for export from the
cell
It also adds directions for the destination
of the protein package
Proteins only enter one side of the stack
(cis face), and exit on the other side (trans
face)
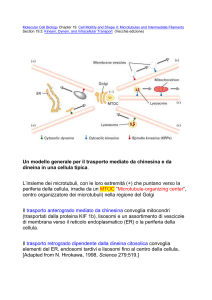
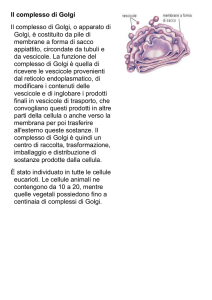
L'APPARATO DEL GOLGI
L'apparato del Golgi è formato da sacche
membranose impilate le une sulle altre, che
modificano proteine e lipidi, sintetizzano carboidrati
e "impacchettano" le molecole che devono essere
trasportate all'esterno della cellula. Anche se può
variare leggermente a seconda delle cellule studiate,
in linea di massima la sua struttura è pressoché
uniforme: è formato da dittiosomi, strutture formate
a loro volta da sacculi appiattiti e da piccole
formazioni cave, le vescicole golgiane. I sacculi
sono impilati strettamente gli uni agli altri e possono
trovarsi singolarmente nel citoplasma o associati nei
cosiddetti corpi golgiani. I sacculi che si trovano
localizzati in prossimità del nucleo sono detti
inferiori o prossimali e costituiscono la regione cis del
dittiosoma. I sacculi che invece si trovano localizzati in
prossimità della superficie cellulare sono detti superiori o
distali e costituiscono la regione trans dello stesso dittiosoma.
L'APPARATO DEL GOLGI
L'apparato del Golgi ha la funzione di rielaborare, selezionare ed
esportare i prodotti cellulari. Questo organulo può interagire con
altri (come il reticolo endoplasmatico rugoso) per indirizzare ed
etichettare certe vescicole contenenti prodotti cellulari verso la loro
destinazione, che può essere quello di confluire in altri organi o
ingranare nella membrana plasmatica e farne uscire il contenuto.
Possiamo fare un esempio con una proteina che deve
raggiungere un lisosoma. Inizialmente questa proteina viene
fornita di una specifica sequenza segnale che la indirizza nel
reticolo endoplasmatico rugoso. Qui, le viene rimossa la sequenza
segnale, sostituita con un oligosaccaride (cioè l'indirizzo in
etichetta), e la proteina ora si chiama glicoproteina. Gli enzimi del
Golgi modificano l'oligosaccaride aggiungendogli un gruppo
fosfato, processo chiamato fosforilazione. La proteina fosforilata,
si lega ad un recettore specifico; dopo viene racchiusa all'interno
di una vescicola mediante l'estroflessione della membrana
plasmatica; in questo modo la proteina resta divisa dal citoplasma
quindi può raggiungere il lisosoma.
Secrezione delle vescicole del Golgi
I lisosomi, vescicole (0.05 – 0.5 microns)
che si formano nell’apparato del Golgi,
sono essenzialmente sacchetti
membranosi (delimitati da una membrana
a doppio strato lipidico) che racchiudono
enzimi idrolitici, in grado, nella digestione
intracellulare, di tagliare in maniera
specifica e controllata grosse molecole.
Lisosomi
Gli enzimi idrolitici dei lisosomi sono detti
idrolasi acide e comprendono
proteasi (che tagliano le proteine),
nucleasi (che tagliano gli acidi nucleici),
glicosidasi( che tagliano i polisaccaridi)
lipasi ecc.
Se i lisosomi si rompono, la cellula stessa
viene distrutta, poiché gli enzimi che essi
contengono sono capaci di scindere tutti i
composti principali presenti nella cellula.
I lisosomi contengono ognuno idrolasi diverse,
che svolgono l’ampia serie di differenti funzioni
digestive.
Le idrolasi dei lisosomi sono sintetizzate nel
reticolo endoplasmatico, come le altre
proteine, e trasferite in seguito nel lume
dell’apparato del Golgi.
I lisosomi si formano per gemmazione (si
staccano come goccioline di membrana) dalle
cisterne più esterne dell’apparato del Golgi.
Bisogna distinguere fra due classi generali
di lisosomi:
i lisosomi primari, appena formati e non
ancora fusi con altre vescicole contenenti i
materiali da digerire; e
i lisosomi secondari, che derivano da
ripetute fusioni di lisosomi con altre
vescicole. In questo secondo tipo sono
contenuti enzimi, materiale da digerire e
materiale digerito.
Nella cellula si trovano altri organuli simili ai
lisosomi, i perossisomi, che ne differiscono
per il contenuto enzimatico. I perossisomi
contengono le catalasi, enzimi specializzati per
effettuare reazioni ossidative, che impiegano
l’ossigeno molecolare. Il perossisoma, con il
mitocondrio, è il principale sito di utilizzazione
dell’ossigeno nella cellula. Grossi perossisomi
delle cellule del fegato e del rene sono
importanti nel neutralizzare la tossicità di
numerose molecole, ossidandole (per esempio,
quasi la metà dell’alcol che beviamo viene
neutralizzata nei perossisomi).
Perossisomi e Lisosomi
il Citoscheletro
Il citoscheletro non è da considerarsi un
organulo cellulare in quanto non è racchiuso da
una membrana. Non bisogna però pensare che
esso sia di scarsa importanza per la cellula: si
può dire che il citoscheletro costituisca
muscolatura e ossatura della cellula. Le sue
funzioni sono di tipo strutturale (ed è
indispensabile quindi nelle cellule eucariotiche),
di protezione e di movimento sia intercellulare
che intracellulare.
Il citoscheletro è costituito da tre tipi di
filamenti proteici che si distinguono per
funzione e composizione proteica:
filamenti actinici o microfilamenti
filamenti intermedi
microtubuli
The Cytoskeleton
Microfilaments are involved with
movement (e.g. actin and myosin)
Microtubules are made of the protein
tubulin and give shape to the cell
Intermediate filaments have many
functions and components: keratin in skin,
hair and nails is an example
Filamenti actinici
la componente proteica di questo tipo di filamenti
è l'actina. L'actina è una proteina globulare che
lega ATP
Con il loro spessore di 6-7nm, sono i filamenti
citoscheletrici più sottili. Hanno una polarità
strutturale, ovvero hanno un'estremità +, dove
l'aggiunta di g-actina avviene velocemente e
contribuisce quindi all'allungamento del filamento,
ed una parte - che influisce poco
sull'accrescimento.
La polimerizzazione inizia lentamente con 3
molecole di actina che si legano tra loro. Nella
cellula la concentrazione di actina libera è molto
alta, quindi altre molecole di actina si legano a
questo polimero neoformato ed il processo diventa
man mano più veloce fino a che non si giunge ad
un punto di equilibrio con l'actina libera nella
cellula.
La miosina
è una proteina presente in tutte le cellule
eucariotiche. È dotata di attività ATPasica, cioè è in
grado di idrolizzare l'ATP e si lega all’actina
Esistono diverse isoforme di miosina all'interno delle
cellule. Nel complesso le varie isoforme funzionano
da "motori proteici"; in pratica accoppiano
l'idrolisi della molecola di ATP con cambiamenti
conformazionali che contribuiscono a generare la
forza meccanica per i vari tipi di motilità cellulare.
La struttura della miosina consiste in due parti
principali: la testa globulare, che lega la molecola di
actina ed è dotata di attività ATPasica (cioè in grado
di idrolizzare l'ATP) e da una coda, unita alla testa,
che consiste in due catene proteiche con
conformazione ad elica avvolte insieme.
Il complesso actina-miosina, nelle cellule
muscolari scheletriche dei Vertebrati,
forma una struttura caratteristica detta
sarcomero, da cui dipende la contrazione
delle fibre muscolari. La contrazione
muscolare è influenzata essenzialmente
dalla concentrazione intracellulare dello
ione calcio, ma anche da altre proteine,
come la tropomiosina, la troponina e la
nebulina
Filamenti intermedi
Chiamati così per il loro spessore (circa 10 nm)
intermedio tra quello dei microtubuli e quello dei
filamenti actinici. Le molecole che li
costituiscono sono filamentose e variano a
seconda del tipo di cellula. possiedono una
grande resistenza alla trazione e consentono
alla cellula di sopportare stress meccanici. A
differenza degli altri filamenti citoscheoletrici, i
filamenti intermedi non sono polarizzati e sono
più stabili. Una categoria di filamenti intermedi
presenti in tutte le cellule è quella delle lamìne,
ovvero quel particolare tipo di filamenti che va a
costituire la lamina nucleare
Microtubuli
Sono tubi proteici cavi del diametro di
25nm (diametro esterno) capaci di
autodemolirsi rapidamente in una sede e
ricostituirsi altrettanto velocemente in
un'altra. le loro pareti sono formate da 13
protofilamenti. Anche i microtubuli sono
polari. Sono composti da eterodimeri
formati da una molecola di tubulina-α e
una di tubulina-β. la tubulina è una
proteina capace di legarsi a GTP, ma solo
la tubulina-β può idrolizzare GTP a GDP.
Modello di interazioni citoscheletriche
Ciglia e flagelli
Speciali proteine possono stabilizzare in
maniera permanente i microtubuli che vanno
così a formare ciglia e flagelli.
Le ciglia servono per il movimento della cellula.
Generalmente sono numerose sulla superficie
cellulare. possono creare correnti nel liquido
intorno alla cellula in modo da indirizzare il cibo
verso il luogo in cui verrà digerito, come succede
per esempio nelle spugne. La parte interna di
un ciglio è detta assonema ed è costituito da
una membrana che racchiude 9 coppie di
microtubuli alla periferia più due micotubuli non
accoppiati al centro. Questa struttura è detta
9+2 e si ritrova in quasi tutte le forme di ciglia e
flagelli eucariotici, dai protozoi all'uomo
Il centriolo è una struttura cilindrica presente
nel citoplasma (in prossimità del nucleo) delle
cellule animali, in alcuni funghi, alghe e in
alcune piante. Un centriolo è formato da nove
triplette di microtuboli, ossia catene di
tubulina alfa e beta che si uniscono a formare
foglietti di protofilamenti, ripiegati su sé stessi.
I centrioli si trovano in coppia a costituire il
centrosoma e, solitamente, sono disposti tra di
loro a formare un angolo di 90°. Essi sono i
responsabili della produzione dei microtubuli
che costituiscono il fuso mitotico, hanno quindi
un'importante funzione nella divisione cellulare.
Microtubuli in un Neurone
il Nucleo
Il nucleo è un organulo che si trova
all'interno della cellula ed è sede di
importanti reazioni.
Il suo scopo è quello di contenere gli acidi
nucleici, provvedere alla duplicazione del
DNA, alla trascrizione e alla maturazione
dell'RNA.
Il nucleo è presente solo negli eucarioti ed
è delimitato da una doppia membrana
fosfolipoproteica in continuità con il RER
The Nucleus and Nuclear Pores