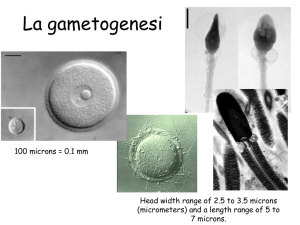
IL CICLO VITALE DEI MAMMIFERI
Cellule germinali primordiali
• 1. Le cellule germinali primordiali (PGC) colonizzano
le gonadi primitive che differenziano in testicoli ed
ovaie.
• 2. Le cellule germinali si moltiplicano e si trasformano
in goni (spermatogoni ed ovogoni) capaci di mitosi.
• 3. Con la gametogenesi (ovogenesi e
spermatogenesi), le cellule germinali entrano in
meiosi e diventano gameti.
• 4. La meiosi nella spermatogenesi inizia alla pubertà,
nell’ovogenesi inizia già nel feto e poi si blocca fino
alla pubertà
LA GAMETOGENESI
• FOLLICOLOGENESI o OVOGENESI
• SPERMATOGENESI
Cellule germinali primordiali (PGC)
Le cellule germinali primordiali (circa 50) si formano dall’epiblasto del
disco embrionale, migrano nella parete interna del sacco vitellino dove
compaiono alla fine della 3^ settimana vicino all’allantoide (a);
dalla 4^ alla 6^ settimana migrano verso le creste genitali (b)
proliferando (circa 5000 all’arrivo).
3^ sett.
Migrazione dalla 4^ alla 6^
settimana
Le cellule PGC sono riconoscibili perché più grandi delle cellule vicine. Sono
positive alla fosfatasi alcalina
• 1. Le PGC colonizzano le gonadi che differenziano
in ovaie e testicoli (ad es.: il gene SRY-Sex
determining Region Y-situato sul cromosoma Y,
codifica per il TDF-Fattore di Determinazione del
testicolo-una proteina che innesca lo sviluppo
maschile).
• 2. Le PGC si moltiplicano e si trasformano in goni
(spermatogoni ed ovogoni) cellule staminali capaci
di mitosi.
• 3. Con la gametogenesi (ovogenesi e spermatogenesi), le cellule germinali entrano in meiosi e
diventano gameti.
• 4. Differenze temporali: la meiosi nella
spermatogenesi inizia alla pubertà, nell’ovogenesi
inizia già nel feto e poi si blocca fino alla pubertà
Cronologia della gametogenesi
RIASSUMENDO:
Meiosi nel sesso ♂ e ♀
Nella ♀, le PGC si differenziano in ovogoni che vanno incontro a mitosi e poi
iniziano la meiosi (ovociti primari) già durante la vita fetale. Blocco degli ovociti
primari in profase I e ripresa della meiosi solo dopo la pubertà. Da un ovocito
primario deriva un solo ovocito maturo e tre globuli polari.
Nel ♂ , le PGC restano quiescenti fino alla pubertà, quando si differenziano in
spermatogoni che entrano in mitosi. Per tutta la vita adulta, gli spermatogoni
producono spermatociti primari, che vanno in meiosi. Da uno spermatocito
primario derivano 4 spermatidi (poi differenziano in spermatozoi).
Sviluppo
sessuale e
gametoge
nesi sono
controllati
dagli
ormoni
FOLLICOLOGENESI
o
OVOGENESI
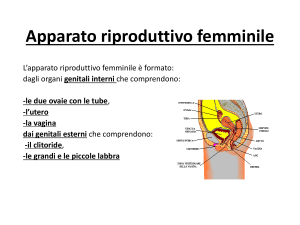
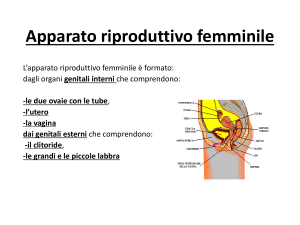
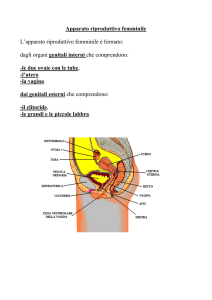
Anatomia del sistema riproduttivo femminile.
L’ovaio
A destra, sezione di ovaio nella zona corticale
(ingrandimento del riquadro rosso).
E, epitelio di rivestimento (basso) dell’ovaio;
TA, tunica albuginea;
F, follicoli;
FP, follicolo primordiale;
TF, teca follicolare;
ZP, zona pellucida;
ZG, granulosa;
O, ovocito.
il follicolo ovarico
• L’ovocito non è nudo ma è all’interno di un
“contenitore” di cellule epiteliali (follicolari).
• l’ovocito e il suo “contenitore” costituiscono il
follicolo ovarico.
• Fino alla pubertà nell’ovaio ci sono solo
follicoli quiescenti (f. primordiali)
Quanti follicoli?
Alla pubertà ~400.000
Follicologenesi
• Dopo la pubertà, ogni giorno un gruppo di
follicoli primordiali inizia a maturare
• Solo uno, in genere, ci riesce giungendo ad
essere ovulato dopo circa un anno di
maturazione (ovulazione=espulsione
dall’ovaio)
• Tra due successive ovulazioni passano
circa 28±7,5 gg.
L’ovaio ha le funzioni di
• Produrre gameti (ovociti)
• Produrre ormoni steroidei (estrogeni e
progesterone)
• Preparare l’utero ad una eventuale gravidanza
• Regolare la crescita degli organi riproduttivi
• Far sviluppare i caratteri sessuali secondari
(CSS)
• Regolare il ciclo mestruale
Ciclo mestruale o uterino
• E’ l’insieme delle modificazioni che
interessano ciclicamente la mucosa
uterina.
• L’evento più eclatante è la mestruazione,
cioè il distacco della parte superficiale
della mucosa uterina
Ciclo mestruale e follicologenesi
• Il tempo tra due successive mestruazioni è pari
anch’esso a 28±7,5 gg (è legato alle attività
dell’ovaio).
• Per comodità di conteggio, ciclo ovarico e
mestruale si fanno iniziare insieme e la
ovulazione, che si verifica a metà circa del ciclo
ovarico, lo divide in due fasi: follicolare (in cui
dominano gli estrogeni) e luteinica (in cui
domina il progesterone).
Nell’ovaio dopo la pubertà ci
sono diversi tipi di follicoli
I DIVERSI TIPI DI FOLLICOLI OVARICI
f. secondario
f. primordiali
f. di Graaf
f. primari
corpo luteo
ovulazione
stadi di maturazione di un follicolo
ovarico
• follicolo primordiale che si risveglia
(reclutamento)
• follicolo primario: dopo 150 gg dal
reclutamento
• follicolo secondario (pre-antrale): dopo 270
gg
• follicolo maturo (di Graaf): ultimo stadio,
dopo circa un anno
seguiamo lo sviluppo del follicolo
destinato ad ovulare
Follicolo primordiale
• Il follicolo primordiale è il tipo di follicolo
predominante nell’ovaio.
Ovocito: Ø 20 µm
Follicolo: Ø 25 µm.
E’ circondato da
cellule follicolari
appiattite.
FOLLICOLOGENESI:
fase pre-antrale
• Un gruppetto di follicoli primordiali (ciascuno
contiene un ovocito primario bloccato in profase
I) si risveglia (è reclutato) ogni giorno senza
intervento di ormoni ipofisari.
• Probabilmente la causa è l’interazione reciproca
tra ovocito (tramite BMP-15, FGF2, GDF9 (growth differentiation factor-9) , PDGF
(platelet-derived growth factor) ed altri fattori
ovocitari) e cellule follicolari e dello stroma
ovarico.
FOLLICOLOGENESI:
fase pre-antrale
• Dopo circa 150 gg si forma il follicolo
primario.
Follicoli primordiali (sono i follicoli più
numerosi nell’ovaio)
Follicoli primari
Si arriva allo stadio di
follicolo primario dopo
150 giorni dal
reclutamento
Lamina basale
Zona pellucida
formata da
glicoproteine ZP1,
ZP2, ZP3 e ZP4
Ricordiamo che gli ovociti
dei follicoli primordiali e
primari sono ancora in
diplotene della profase I
Follicolo primario:
Ovocito: Ø 70 µm
Follicolo: Ø 70-100 µm
Follicolo primario
Esistono due tipi di follicoli primari
(diametro fra 60 e 100 µm):
Lamina
basale
Follicolo primario multilaminare
Particolare di
follicolo
ovarico al ME
GIUNZIONI
GAP
Follicolo secondario o pre-antrale
(follicolo secondario di classe I*)
Si forma dopo circa 270 giorni dal reclutamento
E’ caratterizzato da:
• proliferazione continua delle cellule follicolari che formano una
membrana pluri- o pseudo-stratificata (detta granulosa) di circa 600700 cellule colonnari
• ispessimento della zona pellucida.
Dal follicolo primario al secondario sono coinvolti fattori di crescita
ovocitari (GDF-9, BMP-15; stimolano la proliferazione delle cellule
della granulosa?)
• Lo stadio è anche detto pre-antrale perché non si è ancora formato
l’antro follicolare entro il quale sporgerà l’ovocito
* i follicoli secondari sono classificati in VIII classi in base alle
dimensioni, vedi avanti
Cresce l’ovocita
primario
Fase
preantrale
Fase preantrale. La trasformazione da follicolo primordiale a follicolo primario
avviene con l’accrescimento dell’ovocito, formazione della granulosa e della teca
(interna ed esterna) del follicolo e produzione della zona pellucida.
Alla fine della fase pre-antrale sulle cellule della granulosa del follicolo 2° compaiono
recettori per FSH, prolattina, estrogeni ed androgeni e, sulle cellule tecali, recettori
per l’LH. L’espressione di tali recettori permette l’ulteriore maturazione del follicolo.
Cresce l’ovocita
primario
Fase
preantrale
Comparsa
dei
recettori
per LH
Comparsa
dei
recettori
per FSH
Fase preantrale. La trasformazione da follicolo primordiale a follicolo primario
avviene con l’accrescimento dell’ovocita, formazione della granulosa e della teca
(interna ed esterna) del follicolo e deposizione della zona pellucida.
Alla fine della fase pre-antrale sulle cellule della granulosa del follicolo 2° compaiono
recettori per FSH, prolattina, estrogeni ed androgeni e, sulle cellule tecali, recettori
per l’LH. L’espressione di tali recettori permette l’ulteriore maturazione del follicolo.
follicolo secondario
di classe II (antrale)
Nel follicolo secondario vediamo ora l’inizio
di formazione di spazi tra le cellule della
granulosa. Confluiranno poi nell’antro
follicolare entro il quale sporgerà l’ovocito.
cellule della granulosa
(sintesi di estrogeni)
membr. basale
cellule endocrine
della teca interna
teca esterna
Misure:
Follicolo di 120-200 µm di diametro
Ovocito di circa 100 µm di diametro
Vasi capillari nella teca interna
zona pellucida
ovocito in accrescimento
piccoli spazi isolati
(corpi di Call-Exner)
pieni di liquido.
Inizia a formarsi l’antro
Follicolo secondario antrale (precoce)
Spazi
tra le
cellule
(corpi
di CallExner)
ovocito
Struttura del follicolo secondario
Follicolo secondario tardivo
Durata del processo
• Da follicolo primordiale si passa a primario in
circa 150 giorni.
• Da follicolo primario si passa a secondario in
ulteriori 120 giorni circa.
• Al termine della fase pre-antrale della
follicologenesi si ottiene un follicolo definito
come follicolo secondario di classe I
(preantrale; per la classificazione vedi avanti).
Risultato del processo
6 unità strutturali diverse ma integrate:
• un ovocito in accrescimento (1) circondato dalla zona
pellucida (8);
• cellule della granulosa (2) disposte su 9 strati circa;
• una membrana basale (3);
• una teca interna (4);
• una teca esterna (5);
• una rete capillare entro il tessuto tecale (6).
(7) Corpi di Call-Exner
FOLLICOLOGENESI:
fase antrale
• Con la comparsa dell’antro si passa allo
stadio di follicolo antrale (follicoli secondari
di classe II-VIII)
FOLLICOLOGENESI: fase antrale
• E’ più veloce perché dipende dalle
gonadotropine ipofisarie e dagli ormoni
ovarici;
• il follicolo raggiungerà le dimensioni
massime, come follicolo maturo o di
Graaf, in 80 giorni.
• Nella fase antrale si distinguono 7 classi
follicolari a seconda della diversa
grandezza del follicolo.
Classificazione dei follicoli
2. Fase antrale
L’antro in formazione
si riempie di un fluido
viscoso composto da
proteoglicani e
trasudato del plasma
(contiene anche
estrogeni)
Follicolo secondario con antro
in formazione
Follicolo secondario al MES. Al centro:
ovocita circondato dalle cellule del
cumulo ooforo nella cavità dell’antro
Antro follicolare
Membrana granulosa
ZP
Teca follicolare
2. Fase antrale
L’antro:
fa aumentare il volume del follicolo
(circa 6 mm di diametro) ma non
dell’ovocito;
La causa? secrezione di liquido
antrale e proliferazione delle cellule
follicolari.
ATTENZIONE: Tutto ciò avviene
solo se la produzione di recettori
ormonali, di gonadotropine e di
ormoni steroidei (vedi avanti) sono
adeguate.
Tali produzioni si rinforzano
reciprocamente con meccanismo a
feedback positivo.
Sviluppo dei follicoli antrali fino al follicolo
maturo (di Graaf, di classe VIII)
La corona radiata
l’ovocito
teca
Follicolo di Graaf o follicolo maturo.
Il follicolo misurerà circa 20 mm e
l’ovocito 100-150 µm
Follicolo in degenerazione o atresico
Stadio iniziale di atresìa
follicolare con zona
pellucida collassata
Atresia follicolare
Nel periodo riproduttivo,
una donna ovula circa
400 ovociti. Durante un
ciclo riproduttivo un
gruppo di ovociti inizia
a maturare ma solo 1 o
2 completano
l’ovogenesi e sono
ovulati. Gli altri
subiscono un processo
degenerativo chiamato
atresìa che può
avvenire in qualunque
momento dello
sviluppo del follicolo.
I follicoli nella fase antrale
Come prima detto, nella fase antrale si distinguono 7 classi follicolari in base
alla diversa grandezza del follicolo e secondo la tabella:
il follicolo di classe I è quello secondario; il follicolo di classe II è detto antrale
perché compaiono i primi spazi che confluiranno nell’antro.
Il follicolo di classe VIII è quello più grande
Follicolo di Graaf
Ovocito
teca
Nella successiva fase pre-ovulatoria Il
follicolo misurerà circa 20 mm e l’ovocito
100-150 µm (in media 120 µm)
Follicolo di Graaf
Follicolo di classe VIII (ultimo stadio della fase antrale):
Caratteristiche:
• elevato numero di cellule della granulosa (circa 50-60
milioni) unite da giunzioni comunicanti;
• grosso antro entro cui sporge l’ovocita (diametro di circa
120 µm);
• teca ben sviluppata.
• Il follicolo di classe VIII al massimo dell’ampiezza (20-25
mm) è detto follicolo maturo o di Graaf.
• Ora, alla periferia dell’ooplasma compaiono i granuli
corticali
Granuli
corticali
Torneremo a
parlare del ruolo
dei granuli corticali
subito dopo la
fecondazione
le cellule della granulosa nel follicolo
di Graaf
•
•
Ovocita
•
teca
cellule della granulosa che
delimitano l’antro: formano la
parete del follicolo e poggiano
sulla membrana basale;
cellule della corona radiata:
circondano e nutrono l’ovocita
e costituiscono le cellule più
interne del cumulo ooforo;
cellule del cumulo ooforo:
costituiscono una sorta di
peduncolo che collega le
cellule antrali a quelle della
corona radiata.
androgeni ed estrogeni
due tipi di ormoni strettamente
collegati
: iniziale
Follicolo secondario: steroidogenesi
receptor
Durante la fase antrale le
cellule della teca interna
producono androgeni che la
granulosa aromatizza
trasformandoli in estrogeni
su stimolo del FSH. Le
cellule della granulosa
hanno recettori sia per FSH
che per estrogeni.
Nel follicolo 1° (tardivo) e 2°, le cellule follicolari (1) hanno solo recettori per FSH
mentre recettori per LH sono presenti sulle cellule della teca interna (2).
Steroidogenesi al termine della fase
antrale
Nel follicolo di Graaf, compaiono
recettori per LH sulle cellule della
granulosa e coesistono con
recettori per FSH (cala la sintesi di
recettori per FSH e per estrogeni).
L’acquisizione di recettori per LH è
fondamentale per:
1. iniziare a produrre progesterone
e favorire l’ovulazione;
2. la luteinizzazione del follicolo
dopo la sua rottura e l’ovulazione.
Follicolo di Graaf
Durata del processo
• Dopo due mesi dallo stadio di follicolo
secondario (pre-antrale) tra i follicoli antrali
formatisi finora avviene la selezione del
cosiddetto follicolo dominante, quello
che riuscirà ad ovulare.
• Il reclutamento* del follicolo che riesce ad
ovulare è avvenuto in realtà 20 giorni
prima della ovulazione (quindi durante la
fase luteinica del ciclo ovarico precedente)
-6 gg
0
14 gg (ovulazione)
La crescita esponenziale (20 gg)
1. Il reclutamento (dura 6 giorni),
• inizia nella fase luteinica tardiva del ciclo
precedente (la caduta di estradiolo,
progesterone ed inibina*, stimolano l’aumento di
FSH ed LH).
• È caratterizzata dalla maturazione di follicoli di
classe IV**, che non vanno in atresia, a follicoli
di classe V, che formeranno il pool di follicoli tra
cui sarà scelto quello destinato ad ovulare.
*l’inibina è un ormone proteico (della
granulosa) che inibisce la sintesi di FSH
**
6 gg
La crescita esponenziale (20 gg)
2. La selezione (dura 4 giorni),
• si verifica in fase follicolare precoce: 1-5 follicoli di
classe V diventano follicoli di classe VI.
• Non vanno in atresia perché:
-le cellule della granulosa si dotano di un maggior
numero di giunzioni comunicanti e di recettori per
FSH ed LH
-aumenta la vascolarizzazione della teca.
4 gg
La crescita esponenziale (20 gg)
3. La dominanza precoce (dura 8 giorni),
• si verifica nella fase follicolare intermedia,
durante la quale i pochi follicoli di classe VI
diventano di classe VII e poi tutti tranne uno,
vanno incontro ad atresia.
• Tale follicolo, detto dominante, diventerà di
classe VIII.
8 gg
La crescita esponenziale (20 gg)
4. La dominanza tardiva (dura 2 giorni),
• si verifica in fase follicolare tardiva (preovulatoria),
• il follicolo dominante di classe VIII
diventa follicolo maturo (del diametro
di 20-25 mm).
2 gg
Perché i follicoli selezionati non
vanno in atresia?
• le cellule della granulosa formano un maggior
numero di giunzioni comunicanti e di recettori per
FSH ed LH
• aumenta la vascolarizzazione della teca
• il follicolo dominante concentra nel suo fluido
estradiolo ed anche FSH.
• nel fluido follicolare il rapporto estrogeni/androgeni è
elevato*.
*NB:nei follicoli con elevate quantità di androgeni nel
liquor folliculi, il destino non è l’ovulazione ma
l’atresia.
Perché tra i follicoli reclutati uno
solo è quello dominante?
• Il follicolo dominante è quello che, tra
quelli reclutati, per condizioni metaboliche
ed endocrine (numero di recettori per FSH
sulla superficie), meglio risponde allo
stimolo di FSH e va incontro a tutte le
tappe maturative con modificazioni
morfologiche importanti.
Perché tra i follicoli reclutati e
selezionati uno solo ovula?
• Il follicolo dominante produce la inibina, che esercita un
feedback negativo sulla secrezione di FSH. La
mancanza dell’effetto stimolatorio di FSH sugli altri
follicoli reclutati, ne induce l’atresia (per apoptosi, cioè
morte cellulare programmata).
• Il follicolo dominante continua la sua crescita
producendo crescenti quantità di estrogeni che,
ulteriormente, esercitano un feedback negativo sulla
secrezione di FSH ipofisario con aumento della atresia
dei follicoli non dominanti.
• In questi ultimi il microambiente intrafollicolare è
iperandrogenico per l’incapacità di convertire gli
androgeni in estrogeni (carenza della attività
aromatasica).
Fase pre-ovulatoria
(circa 37 ore)
• La comparsa dei recettori per LH sulle
cellule della granulosa è condizione
necessaria ma non sufficiente perché il
follicolo sia in grado di ovulare. Occorre
che la concentrazione ematica di FSH ed
LH siano più alte dei valori fin qui
raggiunti.
3. Fase pre-ovulatoria
• Dura in media 37 ore e precede l’ovulazione.
E’ caratterizzata da un picco elevato di FSH
e da una ondata di LH dovuti all’azione degli
estrogeni sull’ipofisi (al di sotto di un certo
livello agiscono con feedback negativo; con
l’aumento di estrogeni tale livello e superato
ed il feedback diventa positivo).
3. Fase pre-ovulatoria
•
•
Durante questa fase si ha la ripresa della
meiosi ad opera del MPF (maturation
promoting factor o fattore che promuove la
fase M) sintetizzato dall’ovocito su
stimolazione del progesterone; l’ovocito si
sveglia dal blocco meiotico e si divide
formando l’ovocito secondario e il primo
globulo polare.
L’ovocito secondario inizia la seconda
divisione meiotica ma si arresta di nuovo in
metafase per azione del fattore citostatico
(CSF), sintetizzato poche ore dopo la
maturazione dell’ovocita. Riprenderà la meiosi
solo se fecondato.
3. Fase pre-ovulatoria
•
•
Durante questa fase si ha la ripresa della
meiosi ad opera del MPF (fattore che
promuove la fase M) sintetizzato dall’ovocito
su stimolazione del progesterone; l’ovocito si
sveglia dal blocco meiotico e si divide
formando l’ovocito secondario e il primo
globulo polare.
L’ovocito secondario inizia la seconda
divisione meiotica ma si arresta di nuovo (in
metafase II) per azione del fattore citostatico
(CSF), sintetizzato poche ore dopo la
maturazione dell’ovocita. Riprenderà la meiosi
solo se fecondato.
3. Fase pre-ovulatoria
•
•
•
Durante questa fase si ha la ripresa della
meiosi ad opera del MPF (fattore che
promuove la fase M) sintetizzato dall’ovocita
su stimolazione del progesterone; l’ovocita si
sveglia dal blocco meiotico e si divide
formando l’ovocito secondario e il primo
globulo polare.
L’ovocito secondario inizia la seconda
divisione meiotica ma si arresta di nuovo (in
metafase II) per azione del fattore citostatico
(CSF), sintetizzato poche ore dopo la
maturazione dell’ovocita. Riprenderà la meiosi
solo se fecondato.
Si ha il distacco dell’ovocito e della corona
radiata dal cumulo ooforo (36 ore prima
dell’ovulazione): il complesso ovocito/zona
pellucida/corona radiata galleggia libero nel
fluido follicolare;
3. Fase pre-ovulatoria
•
•
•
L’ovocito secondario inizia la seconda
divisione meiotica ma si arresta di nuovo (in
metafase II) per azione del fattore citostatico
(CSF), sintetizzato poche ore dopo la
maturazione dell’ovocito. Riprenderà la meiosi
solo se fecondato.
Si ha il distacco dell’ovocito e della corona
radiata dal cumulo ooforo (36 ore prima
dell’ovulazione): il complesso ovocita/zona
pellucida/corona radiata galleggia libero nel
fluido follicolare;
Come già detto, le cellule follicolari
esprimono i recettori per l’ormone
luteinizzante (LH) oltre ai già presenti
recettori per FSH. Questo induce il follicolo a
secernere progesterone, fatto critico per lo
sviluppo del corpo luteo (vedi oltre).
PRIMA DIVISIONE MEIOTICA
CONTROLLO DELLA MEIOSI
RA
Crossing-over
OMI
Nel topo, l’acido retinoico (RA)
RA
prodotto dal mesonefro e dalla rete
ovarii, fa entrare in meiosi gli ovogoni
( ovociti primari). Perciò l’RA è il
fattore che innesca la meiosi.
Il primo arresto della meiosi avviene
in diplotene (in uno stadio chiamato
dictiato o dictiotene) ed è causato da
un inibitore della maturazione
ovocitaria (OMI)
OMI prodotto dalle cellule
follicolari. L’arresto dura dalla vita
fetale alla pubertà.
CONTROLLO DELLA MEIOSI:
RA
RIASSUNTO
MPF
CSF
La 1^ ripresa è indotta da MPF
(maturation promoting factor) prodotto
dall’ovocito subito prima
dell’ovulazione (causa: il
progesterone).
Il secondo arresto (in metafase della
sperma II^ divisione meiotica) è causato da un
fattore citostatico (CSF)
tozoo
CSF ovocitario.
OMI
2^ ripresa solo se fecondato.
L’ovulazione
Corona radiata
Fluido viscoso dell’antro
(proteoglicani e trasudato del
plasma)
I cambiamenti periodici degli ovidotti: nel periodo della ovulazione, i padiglioni delle tube
uterine si avvicinano alle ovaie e le fimbrie scorrono sulla loro superficie per raccogliere
l’ovocito.
LA FASE LUTEINICA
• Dura circa 14 giorni:
il follicolo si trasforma in corpo luteo, una
ghiandola che inizia a secernere
progesterone.
2. Pieghe della
membrana
follicolare
contenenti
cellule follicololuteiniche.
Fase luteinica:
formazione del
corpo luteo
Gli spazi tra le
pieghe sono
occupati da
cellule tecaluteiniche, t.
connettivo e vasi
sanguigni
Il picco di LH induce la
trasformazione del
follicolo in corpo
3. Ingresso dei vasi
1. L’antro con il
coagulo di fibrina
è sostituito da
connettivo e vasi
Si formano cellule follicolo-luteiniche e
teca-luteiniche piene di lipidi e
derivanti dalle cellule della granulosa e
della teca interna.
luteo.
Fase luteinica: il
corpo luteo
Il corpo luteo
Le cellule del corpo luteo
che producono steroidi
mostrano tre
caratteristiche tipiche
(1) gocciole lipidiche;
(2) mitocondri con creste
tubulari;
(3) abbondante REL.
Cooperazione funzionale tra cellule teca-luteiniche e follicolo-luteiniche
1. LH stimola le cellule tecaluteiniche a produrre progesterone
ed androstenedione. Quest’ultimo
verrà traslocato alle cellule
follicolo-luteiniche per produrre
estradiolo (tramite α-aromatasi).
2. FSH ed LH stimolano le cellule
follicolo-luteiniche a produrre
estrogeni (a partire da
androstenedione) e progesterone.
3. LH, estrogeni e prolattina
mantengono in vita il corpo luteo.
La prolattina potenzia gli effetti
dell’estradiolo nei confronti
dell’accumulo e utilizzo del
colesterolo da parte delle cellule
follicolo-luteiniche.
Fase luteinica:
regressione del
corpo luteo
Se non c’è
fecondazione, il
corpo luteo
regredisce
(luteolisi) dopo
14 giorni per un
processo di
apoptosi
innescato dalla
diminuzione del
flusso sanguigno
e conseguente
ipossia nel corpo
luteo.
Fase luteinica:
regressione del
corpo luteo
La luteolisi
porta alla
formazione
del corpus
albicans
(tessuto
connettivo che
rimpiazza la
massa delle
cellule
luteiniche
Corpus albicans
Stroma ovarico con
vasi sanguigni
LH, prolattina ed
estradiolo contribuiscono
a tenere in vita il corpo
luteo ma questo
sopravvive solo per 14 gg
in assenza di
fecondazione.
L’hCG (human chorionic
gonadotropin) embrionale
sopprime il meccanismo di
autodistruzione del corpo
luteo (che funzionerà fino
alla 9^-10^ settimana)
L’hCG è rilevabile nel
plasma già all’8° giorno
dalla fecondazione
In sintesi…
hCG embrionale
no
fecondazione
=
regressione
fecondazione
=
salvataggio
REPPORTO TRA CICLO MESTRUALE E FOLLICOLOGENESI
Da follicolo primordiale a primario e poi
secondario passa circa un anno
Se si prende come giorno 1 il primo
giorno della mestruazione…
FASE LUTEINICA (gg 14-28)
OVU
LA
ZIONE
(gg 14)
Il corpo luteo,
inibendo la
produzione
dell’LH e
dell’FSH
ipofisari,
impedisce una
nuova
ovulazione.
interazioni tra ipotalamo, ipofisi ed ovaio durante la
fase luteinica.
Su questo
principio si
basa la pillola,
che funziona
come un
corpo luteo
artificiale.
Meccanismi
di controllo
del ciclo
ovarico