La Conduzione Nervosa
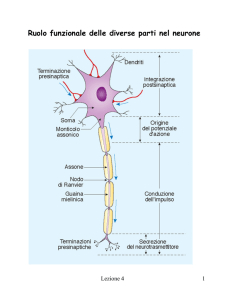
Struttura di una generica cellula nervosa
Forma e dimensioni cambiano molto a seconda della funzione (motoneuroni, interneuroni, neuroni
sensitivi). Elementi caratteristici comuni sono i dendriti (ramificazioni che permettono lo scambio
di “informazioni” con altre cellule in punti detti sinapsi) e l’assone (una struttura lunga e sottile per
la trasmissione dell’impulso elettrico). Nell’uomo, gli assoni hanno diametro compreso tra 1 e 20
µm. La lunghezza è molto variabile, e può arrivare al metro. In alcuni animali superiori gli assoni
sono ricoperti da cellule di Schwann che formano una guaina mielinica.
Modello Elettrico dell’Assone
Parametri Tipici di un assone
Resisitività assoplasma ρA
Capacità per area unitaria di membrana Cm
Resistenza di un area unitaria di membrana Rm
Raggio r
assone mielinico
2 [ohm][m]
5 10-5 [F][m]-2
40 [ohm][m]2
5 [µm]
R= ρ A
l
πr 2
assone amielinico
2 [ohm][m]
10-2 [F][m]-2
0.2 [ohm][m]2
5 [µm]
Se l’assone fosse lungo l=1 cm:
R=ρA l/(πr2)=2x0.01/(3.14x52x10-12)=2.5x108 ohm QUESTO E’ UN VALORE ELEVATISSIMO
Trasmissione dell’impulso elettrico in un circuito RC
A causa della elevata resistenza elettrica dell’assone, e delle forti correnti di dispersione, un impulso
elettrico generato ad una estremità si disperde velocemente, e non riesce quindi a propagarsi lungo
tutto l’assone. La situazione è migliore per gli assoni mielinici: la guaina mielinica riduce la
corrente dispersa e pertanto un impulso elettrico si può propagare per un tratto maggiore. Anche per
l’assone mielinico però la dispersione è tale per cui l’impulso si propaga solo per un beve tratto.
Questo modello elettrico è in grado di spiegare il comportamento dell’assone sotto soglia.
Per comprendere il funzionamento sopra soglia (la propagazione del potenziale d’azione) è
necessario considera altri fenomeni elettrici.
Concentrazioni Ioniche
I principali ioni che possono attraversare la membrana cellulare non si trovano alla stessa
concentrazione all’interno ed all’esterno della cellula. Tipicamente si osserva che:
Na+
K+
Cl(altri)-
(concentrazioni in moli per m-3)
Fluido Esterno
145
4
120
29
Fluido Interno
12
155
4
163
Inoltre, il potenziale elettrico all’interno dell’assone è più basso che all’esterno. Tipicamente:
Potenziale V
Fluido Esterno
0
Fluido Interno
-90 mV
Una differenza di potenziale ai capi di una membrana semipermeabile potrebbe giustificare una
diversa concentrazione ionica. La differenza di potenziale per una data concentrazione è data dalla
equazione di Nernst
equazione di Nernst
q(VI − VE ) = k B T ln
cE
cI
q è la carica dello ione; quindi q(VI-VE) è l’energia potenziale dello ione; in condizione di equilibrio
questa energia potenziale è uguale al lavoro necessario per spostare lo ione dalla zona a
concentrazione minore a quella a concentrazione maggiore (membro di destra dell’equazione). cE e
cI sono le concentrazioni, ln il logaritmo naturale, T la temperatura assoluta, kB una costante.
Applicando l’eq. di Nernst alle concentrazioni Cl-, Na+ e K- per valutare il potenziale di equilibrio
otteniamo:
per Cl- =-90 mV
questo valore è uguale al potenziale di riposo, quindi Cl- è in equilibrio.
per K+ =-98 mV
questo valore è un po’ minore del potenziale di riposo, quindi il flusso dovuto alla differenza di
concentrazione è un po’ maggiore di quello dovuto alla differenza di potenziale.
per Na+ = +66 mV
questo valore è COMPLETAMENTE diverso dal potenziale di riposo, ed il flusso dovuto alla
differenza di concentrazione non può assolutamente compensare quello dovuto al campo elettrico.
Correnti ioniche dovute a differenza di concentrazione IC e a differenza di potenziale IP. a) entrambe
le correnti entrano nella cellula; b) i due flussi si bilanciano esattamente; c) IC è leggermente
maggiore di IP, e ne risulta un piccolo flusso verso l’esterno.
Perché una tale distribuzione di concentrazioni possa conservarsi nel tempo, è necessaria la
presenza di un nuovo meccanismo cellulare: la pompa sodio-potassio.
Pompa Na-K
E’ un meccanismo di trasporto attivo che genera un flusso ionico a spese di un lavoro metabolico
della cellula. Trasporta ioni Na+ fuori dalla cellula, e per ogni ione spostato esternamente, ne
trasporta uno K+ all’interno. In questo modo NON TRASPORTA CARICA. Però la membrana è
100 volte più permeabile a K+ che a Na+, e quindi gli ioni potassio rientrano molto più velocemente
di quelli sodio. Si forma così un eccesso di carica positiva all’esterno della cellula. Per compensare
questo eccesso di carica, gli ioni Cl- attraversano la membrana trasferendosi all’esterno.
Quest’ultimo è un trasporto passivo, non causato direttamente dalla pompa sodio-potassio.
All’equilibrio è la differenza di concentrazione di Cl- tra interno ed esterno a determinare la
differenza di potenziale di –90 mV in base alla eq. di Nernst. Si noti che variazioni della
permeabilità della membrana a Na+ e/o a K+ causeranno variazioni nel potenziale a riposo.
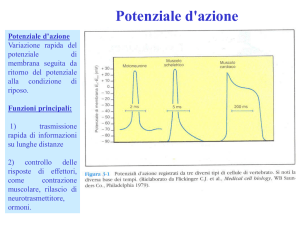
Potenziale d’azione
Se si applica uno stimolo elettrico sufficientemente elevato (sopra soglia) all’assone, si genera una
depolarizzazione che si trasmette lungo l’assone (potenziale d’azione). Lo stimolo elettrico sopra
soglia aumenta la permeabilità della membrana a Na+ di un fattore 1000! Quindi si genera un
improvviso flusso di cariche Na+ che rientra nell’assone. Per questo il potenziale di membrana
raggiunge valori prossimi al potenziale di equilibrio di Na+ (+66 mV). Subito dopo la permeabilità a
Na+ ritorna a valori normali, mentre aumenta quella a K+; per cui si osserva un flusso verso
l’esterno di K+ che porta il potenziale vicino al valore di equilibrio di K+ (-98 mV). La pompa
sodio-potassio, che non ha mai smesso di lavorare, riporta le concentrazioni ai valori a riposo, e
quindi il potenziale a –90 mV.
Propagazione del potenziale d’azione
A causa del potenziale a riposo di –90 mV, sulla parte esterna della membrana dell’assone si
accumuleranno cariche positive, e cariche negative sulla parte interna (i due lati della membrana
dell’assone si comportano come le due armature di un condensatore). Immaginiamo che sia
avvenuta una depolarizzazione sopra soglia in un certo punto dell’assone (b). Poiché il potenziale è
passato da –90 a +66 mV, si è invertita la distribuzione di cariche localmente. Per questo motivo si
creano due flussi di ioni: internamente. ioni positivi si allontanano dal punto di depolarizzazione,
mentre esternamente si avvicinano a questo punto. Quindi nei pressi del punto dove è avvenuta la
depolarizzazione, il potenziale cresce passando sopra soglia causando l’aumento della permeabilità
della membrana a Na+ (c). In questo modo il fronte di depolarizzazione si sposta. Il successivo
aumento di permeabilità della membrana al K+ e riduzione di permeabilità al Na+ riporta il
potenziale ai valori negativi (d). In questo modo si sposta un singolo fronte di depolarizzazione
lungo l’assone.
Nell’assone mielinico gli ioni possono passare attraverso alla membrana solo in corrispondenza dei
nodi di Ranvier. Quindi il fronte di depolarizzazione salta da un nodo all’altro. Questo permette una
trasmissione del potenziale d’azione molto più veloce. Inoltre l’energia metabolica necessaria per
riportare l’assone allo stato di riposo è assai inferiore.