L’IMPULSO NERVOSO
I neuroni utilizzano due tipi di segnali elettrochimici per diffondere l'impulso nervoso attraverso tutta la rete
nervosa:
I potenziali d'azione possono propagarsi senza distorsione da una estremità all'altra e si basano su
processi attivi; è un evento breve che si propaga senza alterazioni lungo tutto l'assone e lungo tutte le
fibre nervose con differente velocità. Queste infatti risultano più elevate negli assoni di grande
diametro rispetto a quelli di piccolo diametro.
I potenziali locali sono in grado di far svolgere alle singole cellule nervose funzioni integrative e di
dare origine al potenziale d'azione; sono graduati e possono estendersi solo su brevi distanze di solito
dell'ordine di 102 mm e hanno un ruolo essenziale nelle terminazioni nervose sensoriali, infine essi
sono passivi e sono soggetti all'attenuazione e alla distorsione.
Per comprendere i processi di trasmissione dei segnali elettrici è necessario conoscere prima alcuni aspetti
strutturali riguardanti l'assone o fibra nervosa. La fibra può essere considerata come un tubo riempito con
soluzione acquosa di sali, dissociati in ioni carichi negativamente e positivamente, separata dalla soluzione
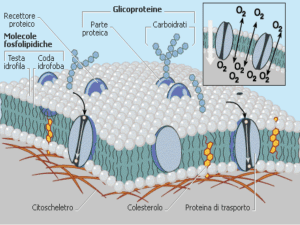
extracellulare per mezzo di una membrana. Il doppio strato lipidico della membrana dell'assone è,
analogamente a quello delle altre membrane cellulari, impermeabile agli ioni e a gran parte delle molecole
polari. L'impulso nervoso si propaga lungo tutto il prolungamento principale del neurone (assone) fino a dove
questo termina facendo “contatto” con un'altra cellula (neurone o fibra muscolare per esempio). Il punto di
giunzione della terminazione nervosa con un altro neurone si chiama sinapsi: qui avvengono una serie di
importanti processi che permettono la trasmissione dell'impulso all'altro neurone.
I neuroni che trasferiscono l'impulso nervoso verso la sinapsi prendono il nome di neuroni presinaptici; quelli
che lo trasferiscono a valle della sinapsi, neuroni postsinaptici. I due neuroni sono separati in realtà da uno
spazio extracellulare, detto fessura sinaptica, dell'ampiezza di circa 200 Å (ÅNGSTRÖM).
Il terminale assonico del neurone presinaptico è una struttura complessa, chiamata bottone sinaptico,
contenente un mediatore chimico (neurotrasmettitore) che si accumula in piccole vescicole.
Attivato il potenziale d'azione nel neurone presinaptico, viene liberato il neurotrasmettitore dal bottone
sinaptico, che si diffonde nella fessura sinaptica e si lega con siti recettori della membrana postsinaptica.
Questo legame rigenera il potenziale d'azione nel neurone postsinaptico.
Il passaggio degli ioni attraverso la membrana dipende dalla presenza di specifiche proteine, dette recettori.
Esse interagiscono con il neurotrasmettitore specifico (dopamina, serotonina, noradrenalina, acetilcolina..). Le
proteine implicate nel trasporto di ioni attraverso le membrane a livello sinaptico si distinguono in due
categorie:
Proteine che costituiscono canali di passaggio per determinati ioni. Esse si aprono e si chiudono a
seconda della conformazione assunta dalla proteina stessa, in funzione dei cambiamenti che avvengono
attorno ad essa. Queste proteine sono definite canali ionici e si possono suddividere in funzione del
meccanismo di regolazione dell’apertura in:
o canali sensibili al potenziale di membrana
o recettori-canale i quali regolano l'apertura o la chiusura dei canali a seguito del legame con
specifiche sostanze chimiche presenti fuori dalla cellula (ormoni o neurotrasmettitori) o nella
cellula stessa.
Proteine con funzione di pompa: ne è un esempio la pompa sodio-potassio che agisce contro
gradiente consumando energia, espellendo ioni sodio in cambio di potassio e determinando così una
ridotta concentrazione di sodio e un eccesso di potassio nell'ambiente intracellulare.
Il gradiente di concentrazione ionico, che si viene a formare grazie all’azione delle pompe sui due versanti della
membrana plasmatica, causa un eccesso di cariche negative all'interno del citoplasma, definito potenziale di
riposo. Possiamo così parlare di:
Membrane polarizzata, a riposo (interno negativo, da -50 a -100 mV)
Membrana depolarizzata, se in seguito ad uno stimolo diventa meno negativa all'interno rispetto
all'esterno.
Membrana iperpolarizzata, se diventa più negativa.
Grazie a dei microelettrodi è possibile misurare la differenza di potenziale tra l'interno e l'esterno della cellula.
Il potenziale di riposo è attorno a -70 mv. Se la membrana del neurone viene depolarizzata (per un impulso
elettrico o a seguito dell’apertura di canali di membrana, ad esempio per la presenza di un neurotrasmettitore) si
aprono canali voltaggio-dipendenti per il sodio, che lasciando entrare sodio (carico positivamente)
depolarizzano ulteriormente la membrana. Il processo si amplifica da solo perché la membrana depolarizzata
nelle zone adiacenti viene depolarizzata dal potenziale d'azione stesso e riproduce a sua volta un potenziale
d'azione. In questo modo si determina la propagazione dell'impulso nervoso finché la membrana è
completamente depolarizzata o addirittura l'interno della cellula diviene positivo rispetto all'esterno (+30 mv).
Successivamente, i canali al sodio si inattivano rapidamente pertanto la violenta depolarizzazione (potenziale
d'azione) si spegne in 2 millisecondi e la membrana non risulta stimolabile per alcuni ulteriori millisecondi.
L'impulso non può tornare indietro, a causa del periodo refrattario che segue al potenziale d'azione nel
segmento d'assone interessato. La propagazione dello stimolo avviene esclusivamente in una direzione grazie al
fatto che, nel tratto interessato dal potenziale d’azione, la pompa sodio-potassio sta operando
per riportare le condizioni a riposo.
QUINDI, PER MEGLIO COMPRENDERE POSSIAMO IN SINTESI DIRE CHE….
LA TRASMISSIONE DELL'IMPULSO NERVOSO
Tra la superficie interna e quella esterna della membrana cellulare del neurone esiste una differenza di
potenziale elettrico (potenziale di riposo, o potenziale di membrana): l'interno della cellula è caricato
negativamente rispetto all'esterno. Tale differenza è dovuta a diverse concentrazioni di ioni sodio (Na +) e
potassio (K+) ed è a sua volta prodotta da meccanismi attivi di trasporto degli ioni attraverso la membrana
(pompa sodio-potassio) e dalla diversa permeabilità della membrana plasmatica rispetto ai singoli ioni.
L'impulso nervoso determina una variazione della permeabilità della membrana, che si traduce in ultima analisi
in un'inversione improvvisa della carica elettrica della cellula. Questa variazione elettrica (che, come si è detto,
costituisce la "risposta") prende il nome di potenziale d'azione.
IL POTENZIALE D'AZIONE
Nella fase d'attivazione del potenziale d'azione, aumenta la permeabilità agli ioni sodio, che irrompono nella
cellula, mentre una modesta quantità di ioni potassio l'abbandona: il potenziale di riposo finisce per diventare
positivo all'interno e negativo all'esterno della cellula.
A questo punto diminuisce bruscamente la permeabilità agli ioni sodio, che fuoriescono in parte dalla cellula
sottraendo cariche positive, mentre aumenta quella agli ioni potassio. La somma totale di questi avvenimenti
ripristina il potenziale di riposo (cioè lo fa tornare negativo nel citoplasma cellulare).
Come risultato di queste depolarizzazioni e ripolarizzazioni attraverso la membrana cellulare si stabilisce un
segnale di natura elettrochimica, trasferibile lungo la fibra nervosa. In ultima analisi, il potenziale d'azione di
una regione della membrana cellulare innesca la produzione di un potenziale analogo in una zona limitrofa.
La conduzione dell'impulso nervoso nelle fibre mieliniche è detta saltatoria, poiché il potenziale d'azione salta
da un nodo di Ranvier all'altro. Inoltre, la guaina mielinica aumenta la velocità di propagazione dell'impulso
nervoso fino a 400 km/h. Nelle fibre amieliniche invece la modalità di conduzione è continua e molto più lenta.