
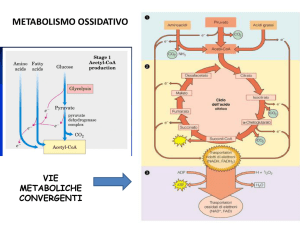
Ciclo di Krebs
1
Complesso della piruvato deidrogenasi (1)
Stechiometria della reazione finale
2
Complesso della piruvato deidrogenasi (2)
FAD
Diidrolipoil
deidrogenasi
Piruvato decarbossilasi
Diidrolipoil transacetilasi
E3
E1
E2
3
Complesso della piruvato deidrogenasi (3)
Le cinque tappe della decarbossilazione ossidativa
4
Ciclo di Krebs
ΔG°’=-32,2 kJ/mole
ΔG°’=-3,8 kJ/mole
ΔG°’=+13,3 kJ/mole
ΔG°’=0 kJ/mole
ΔG°’=-2,9 kJ/mole
5
ΔG°’=-33,5 kJ/mole
Citrato sintasi
Meccanismo
d’azione della citrato
sintasi
Centro ferro-zolfo dell’aconitasi
Meccanismo d’azione dell’isocitrato deidrogenasi
Succinil –CoA sintetasi
Unica reazione del ciclo di Krebs che porta
alla formazione di ATP con un meccanismo
analogo a quanto visto nella glicolisi
(fosforilazione al livello del substrato)
con la differenza che il substrato ad alto
potenziale di trasferimento del fosfato è legato
all’enzima tramite un’istidina
Fosforilazione al livello del substrato
10
Bilancio del ciclo di Krebs
I due C eliminati in ogni giro del
ciclo come CO2 non sono quelli
entrati come CH3-CO(lo saranno nei cicli successivi)
2,5 ATP
Per ogni giro del ciclo avremo
10 molecole di ATP per
molecola di Acetil-CoA ossidato
(nella fosforilazione ossidativa)
2,5 ATP
2,5 ATP
1,5 ATP
1 glucosio = 2 piruvato = 32 molecole ATP
32 x 30,5 kJ/mole = 976 kJ/mole (34% di resa)
2840 : 100 = 976 : X
X = 34 (efficienza in vivo ≈ 65%)
11
I componenti del ciclo sono importanti intermedi biosintetici
Il ciclo di Krebs è il cuore del metabolismo intermedio
(via anfibolica)
12
Reazioni anaplerotiche
Le reazioni anaplerotiche riforniscono il ciclo di Krebs dei suoi intermedi
La più importante è la
reazione catalizzata dalla
piruvato carbossilasi
13
Regolazione del ciclo di Krebs
Regolazione all’ingresso
(complesso piruvato deidrogenasi)
• Inibizione da prodotto (Acetil-CoA, NADH)
• Regolazione feed-back (ATP/AMP, NADH/NAD+)
• Regolazione covalente su E1 (chinasi /fosfatasi)
• Concentrazione Ca2+ (nel muscolo)
Regolazione delle prime tre tappe
intermedie irreversibili
• Disponibilità del substrato
(acetil-CoA, ossalacetato)
• Inibizione da prodotto
• Regolazione feed-back
(ATP/AMP, NADH/NAD+)
• Concentrazione Ca2+ (nel muscolo)
14
Ciclo del Gliossilato
Presente nei semi delle piante in
germinazione e in alcuni microorganismi.
Converte l’acetil-CoA proveniente dalla
degradazione degli acidi grassi in succinato
che poi, attraverso il ciclo di Krebs e la
gluconeogenesi, forma glucosio
Ciclo di Krebs nei mitocondri
15
Ciclo del Gliossilato
Relazione tra il ciclo del gliossilato
e il ciclo dell’acido citrico
Cellulosa
amminoacidi
Energia
Nucleotidi
Intermedi metabolici
16
Ciclo del Gliossilato
Il ciclo di Krebs e del gliossilato
sono regolati in modo coordinato
dalla isocitrato deidrogenasi
Regolazione covalente
Regolazione allosterica
17
Nei ruminanti la gluconeogenesi è molto attiva
Cellulosa
Glucosio
Cellulasi
batterica
Glucosio
Ossalacetato
Fermentazione
propionica
ATP + CoA
Propionato
AMP + PPi
Acil-CoA sintasi
Glucosio
Malato
Ciclo di
krebs
Fegato
18
Vitamina B12 (1)
E’ un complesso composto chimico organometallico (con un atomo di cobalto inserito in un
anello simile a quello del gruppo eme dell’emoglobina)
Tipo di reazione catalizzata
Anello corrinico
La carenza di Co2+ nell’alimentazione può portare a
forme di anemia, inappetenza e ritardo nella crescita
(caso di alcuni allevamenti bovini australiani)
19
Vitamina B12 (2)
La vitamina B12 è presente solo nella carne. Non può essere sintetizzata dagli organismi
animali ne vegetali ma solo da un limitato numero di microorganismi spesso presenti
nell’intestino degli animali superiori. Gli erbivori la ottengono dai batteri del loro stomaco
(i conigli ingeriscono periodicamente una parte delle loro feci per ottenere vitamina B12)
La carenza o il mancato assorbimento di vitamina B12 può causare gravi malattie.
•“Anemia perniciosa”(riduzione degli eritrociti, disfunzioni del SNC) dovuta alla carenza
del fattore intrinseco, una glicoproteina essenziale per l’assorbimento della vitamina.
Si può curare con l’assunzione di dosi elevate di vitamina B12
• Acidemia metilmalonica causata da carenza di L-metilmalonil-CoA mutasi (accumulo
di L-metilmalonil-CoA con conseguente diminuzione del pH, fatale nei primi anni di vita)
20








![11. Ciclo di Krebs [modalità compatibilità]](http://s1.studylibit.com/store/data/001120215_1-23827c0a1e611852a5ba14474c14bb9c-300x300.png)

