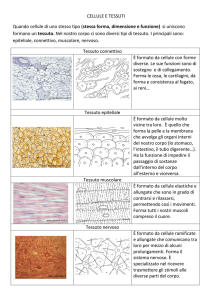
TESSUTO MUSCOLARE
La contrattilità è una proprietà della materia
vivente posseduta da tutte le cellule. Negli
organismi unicellulari o nelle cellule isolate di
metazoi la proprietà contrattile è legata al
citoscheletro, mentre negli organismi superiori la
funzione contrattile è devoluta ad un tessuto
particolarmente differenziato: il tessuto muscolare.
Sulla base delle caratteristiche fisiche delle miofibrille che mostrano
sempre una striatura longitudinale apprezzabile al M.E., il tessuto
muscolare può essere suddiviso in:
Tessuto muscolare striato, caratterizzato dalla presenza di striature
trasversali apprezzabili al M.O., distinguibile in:
¾scheletrico, volontario, che mostra contrazioni potenti e non ritmiche
¾cardiaco, involontario, con contrazioni veloci e ritmiche
Tessuto muscolare liscio, involontario, con contrazioni lente e ritmiche
Tessuto muscolare striato
scheletrico
E’ localizzato:
nei muscoli scheletrici di tutto il corpo
nella lingua, nel palato molle, nella
faringe, nella laringe,
in tratti più o meno estesi dell’esofago
di alcune specie, nel tratto terminale del
retto,
in tratti dell’apparato genitale,
nei muscoli mimici,
in alcuni muscoli dell’occhio e
dell’orecchio
Le fibre muscolari scheletriche sono
elementi multinucleati derivati dalla
fusione
di
mioblasti
embrionali
mononucleati, citologicamente parlando
sono quindi sincizi strutturali, hanno
forma cilindrica o poliedrica con le
estremità arrotondate. La lunghezza
delle fibre va da qualche mm a parecchi
cm, mentre la larghezza varia tra 10 e
100 µm.
Le fibre muscolari sono delimitate da una membrana detta sarcolemma che
racchiude il sarcoplasma nel quale sono immersi gli organuli cellulari. Il
sarcolemma è di fatto l’insieme della membrana plasmatica, della membrana
basale e di fibre reticolari. I nuclei, che possono arrivare a parecchie centinaia,
hanno localizzazione periferica. I mitocondri sono disposti parallelamente lungo
l’asse maggiore delle fibre. Gli altri organuli, il glicogeno e la mioglobina si trovano
sparsi nel sarcoplasma.
Le miofibrille sono strutture filamentose altamente differenziate di 1-2 µm di
spessore. Risultano caratterizzate da bande scure (A = anisotrope) e bande chiare
(I = isotrope) che si alternano. Ciascuna banda I risulta divisa in due parti dalla
linea Z o telofragma. Ciascuna banda A appare occupata nella parte centrale da
una sottile banda più chiara denominata banda H (stria di Hensen) attraversata
centralmente dalla linea M. La regione della miofibrilla compresa tra due linee Z è
denominata sarcomero e costituisce l’unità contrattile della miofibrilla stessa.
M
Le miofibrille sono costituite da miofilamenti distinti in:
spessi, costituiti da miosina, hanno un diametro di 10-20 nm ed una lunghezza di 1.5 µm.
Presentano, ad intervalli regolari, dei prolungamenti laterali detti ponti che assicurano il
contatto con i miofilamenti sottili durante la contrazione. Presentano inoltre al centro sottili
espansioni oblique di natura proteica che formano la linea M, occupano tutta la banda A. Il
miofilamento spesso è costituito dall’associazioni antiparallele di molecole di miosina. La
miosina rappresenta li 55% delle proteine totali del muscolo, ha forma di mazza da golf ed
è costituita da 6 polipeptidi (2 catene pesanti e 4 catene leggere) che formano una parte
lineare detta corpo, ed una parte globulare detta testa. Le molecole, sfasate di 14.3 nm,
sono disposte con la coda rivolta al centro del miofilamento e la testa verso le estremità.
sottili, costituiti da actina, tropomiosina e troponina, hanno un diametro di 5-7 nm ed
una lunghezza di 1 µm. occupano la banda I e si addentrano nella banda A fino al limite
della banda H.
L’actina costituisce il 25% delle proteine del
muscolo, esiste in forma globulare (G-actina), che
in presenza di Ca++ ed ATP polimerizza in actina
filamentosa (F-actina). Nel miofilamento sottile due
molecole di F-actina si avvolgono a doppia elica; a
livello della linea Z la polarità della F-actina si
inverte. L’actina è strettamente correlata con un
complesso tropomiosina-troponina.
troponina
G-actina
tropomiosina
F-actina
La tropomiosina rappresenta il 5%
delle proteine del muscolo, è costituita
da due catene avvolte ad elica ed è
localizzata nei due solchi della doppia
elica di actina per una estensione di
40nm. A far da ponte tra segmenti
consecutivi di tropomiosina si dispone
la troponina. La funzione della
tropomiosina
è
esclusivamente
regolativi in quanto inibisce lo
scorrimento delle teste miosiniche nelle
docce
actiniche.
La
troponina
costituisce il 5% delle proteine del
muscolo, non stabilisce alcun legame
diretto con l’actina, ma regola il
meccanismo
d’azione
della
tropomoisina.
E’
una
proteina
globulare costituita da 3 polipeptidi:
troponina T, che stabilisce il contatto
con la tropomiosina, troponina I, che è
implicata
nell’inibizione
della
formazione dei complessi actinomiosinici, troponina C che possiede un
sito di legame per gli ioni Ca++.
La linea Z è formata da filamenti di α-actinina,
che unisce i filamenti di actina tra loro, e da
desmina, che fa parte del citoscheletro e
coopera all’organizzazione dei miofilamenti in
ciascuna miofibrilla. La linea Z si trova solo nei
vertebrati e negli artropodi.
In un muscolo le fibrocellule muscolari striate
sono disposte parallelamente e sono tenute
insieme da tessuto connettivo. Ogni muscolo
è rivestito dall’epimisio che invia setti
connettivali destinati a circondare fasci di
fibre (perimisio), ogni singola fibra è rivestita
dall’endomisio in cui prevalgono le fibre
reticolari.
Nei Mammiferi le fibre muscolari che
compongono i muscoli sono di due tipi e
variano in percentuale a seconda del tipo di
muscolo:
9fibre rosse, a contrazione lenta e
sostenuta,
posseggono
sarcoplasma
abbondante rispetto alle miofibrille e ricco in
mioglobina e citocromi. Sono più corte e
sottili delle altre,
9fibre bianche, a contrazione rapida, sono
caratterizzate da scarso sarcoplasma e
numerose
miofibrille
disposte
uniformemente. Sono meno ricche in
mioglobina e citocromi delle rosse.
Il reticolo sarcoplasmatico è
rappresentato da REL, è costituito
da una serie di tubuli e canalicoli a
fondo
cieco
disposti
longitudinalmente rispetto all’asse
della
fibra
e
provvisti
di
anastomosi laterali, che formano
una rete intorno a ciascuna
miofibrilla. A livello della banda H
le anastomosi danno luogo alla
cisterna fenestrata dalla quale
partono dei tubuli longitudinali che
a livello del confine tra banda I e
banda A si trasformano in cisterna
terminale. Le cisterne terminali
che si affrontano sono in contatto
con il tubulo T (traverso) che è
una profonda invaginazione del
sarcolemma, l’insieme che si
forma viene denominato triade
sarcoplasmatica.
Meccanismo di contrazione
In sezione trasversale ogni miofilamento spesso è
circondato da 6 miofilamenti sottili, mentre gruppi
di 3 miofilamenti sottili sono circondati da 3
miofilamenti spessi.
Nelle miofibrilla a riposo il complesso
tropomiosina-troponina maschera i siti actinici di
attacco per le teste della miosina.
All’interno del reticolo sarcoplasmatico una
proteina specifica, la calsequestrina, lega gli ioni
Ca++ che vengono liberati nel citoplasma quando
la fibrocellula muscolare riceve l’impulso nervoso.
Il calcio si lega alla troponina C, questo determina
un cambiamento conformazionale della proteina
che porta all’allontanamento della tropomiosina dai
siti actinici per la miosina
I subframmenti S1 della meromiosina pesante si
attaccano ai siti specifici della G-actina e ruotano
assumendo
un’inclinazione
diversa
e
determinando lo scorrimento del filamento sottile
verso il centro del sarcomero.
Durante la contrazione l’accorciamento del
sarcomero porta alla riduzione, fino alla
scomparsa, della banda H e alla contemporanea
riduzione della banda I, mentre l’ampiezza della
banda A resta immutata.
Tessuto Muscolare striato cardiaco
Costituisce il miocardio cioè il tessuto muscolare cardiaco. E’ formato da cellule dette cardiociti,
mononucleate ed anastomizzate tra loro a formare una rete. Hanno forma cilindrica e spesso
terminano ad Y. Le loro dimensioni variano da specie s specie (5-19 µm di diametro). Presentano
la tipica bandeggiatura trasversale dovuta all’organizzazione in registro dei sarcomeri. Al limite tra
due cellule confinanti, a livello delle linee Z sono presenti dischi intercalari, a volte visibili al MO,
conosciuti anche come strie scalariformi e sono zone di contatto tra cardiociti contigui, formate da
segmenti trasversali in cui sono presenti desmosomi, fasce occludenti ed aderenti e gap junctions,
e segmenti longitudinali dove mancano i desmosomi.
Il sarcolemma
è costituito da una membrana plasmatica con uno spesso rivestimento
mucopolisaccaridico e si estende a formare un tubulo T, di diametro maggiore rispetto a quello
delle fibre scheletriche, a livello della linea Z dei sarcomeri. Il nucleo, di forma ovoidale, è
localizzato al centro della cellula. Il Golgi è situato in vicinanza del nucleo, i mitocondri sono
disposti in fila tra le miofibrille e occupano il 50% del volume della cellula. Il sarcoplasma
abbondante contiene glicogeno, gocciole lipidiche, miofibrille con caratteristiche analoghe a quelle
scheletriche, ma non raggruppate in fascetti.
Il reticolo sarcoplasmatico è
formato da elementi tubulari che
formano una rete tridimensionale.
A livello della linea Z il reticolo
contrae rapporti con li tubulo T
senza però formare cisterne
terminali complete, per cui si
forma una struttura denominate
diade sarcoplasmatica.
Nel Tessuto muscolare striato
cardiaco non tutte le cellule sono
raggiunte da fibre nervose, la
presenza di gap junctions fa sì
che la trasmissione dell’impulso
elettrico tra le cellule sia
immediata, questo tessuto si
comporta di fatto come un
sincizio funzionale.
Tessuto Muscolare liscio
Le fibrocellule muscolari lisce sono mononucleate con nucleo in posizione centrale,
di forma fusata, provviste di miofibrille disposte longitudinalmente, ma prive di
striature trasversali. Hanno dimensioni che variano tra 20-100 µm x 4-8 µm.
Presentano un rivestimento di natura glicoproteica ricco in fibre reticolari. Il REL è
poco sviluppato.
Le miofibrille hanno un diametro di 0.5-1 µm e presentano tre tipi di filamenti:
¾sottili, formati da actina, che differisce da quella del muscolo striato per sequenza
aminoacidica,
¾spessi, formati da miosina con minore attività ATPasica e più sensibile all’azione
del calcio che provocherebbe la polimerizzazione in filamenti al momento della
contrazione,
¾intermedi, rappresentati da vimentina nei vasi e desmina negli altri organi.
Nella fibrocellula muscolare liscia, l’apparato contrattile sembra organizzarsi al
momento della contrazione creando un ritardo eccitamento-contrazione.
Sono presenti due proteine regolative: la tropomiosina e l’α-actinina. L’azione della
troponina C nella modulazione del calcio è svolta dalla calmodulina.
Il tessuto muscolare liscio è localizzato nelle muscolaris mucosae delle tonache
mucose degli organi cavi, come pure nelle tonache muscolari di molti di essi. Si trova
inoltre localizzato nella parete di arterie, vene e vasi linfatici e in alcuni dotti escretori
ghiandolari.
Fisiologicamente la muscolatura liscia è divisa in due categorie:
9muscolatura liscia multiunitaria, in cui ogni fibrocellula riceve una terminazione motrice
(muscolatura vascolare),
9muscolatura liscia viscerale, in cui solo poche cellule risultano innervate, tuttavia la
presenza tra le cellule di gap junctions permette il passaggio rapido dell’impulso a tutta la
muscolatura.