Diversità nel ciclo cellulare tra cellule
dell’organismo adulto e cellule embrionali
La durata del ciclo varia molto,
nelle cellule embrionali le fasi
G1 e G2 sono quasi inesistenti
La durata della fase G1 è molto
variabile e condiziona la durata
complessiva del ciclo cellulare
Nella fase G1 la cellula compie le
sue normali funzioni metaboliche
proprie del suo stato di differenziamento,
cresce ed accumula ATP per entrare in
fase S e quindi in Mitosi
La fase S ha una durata costante
per le varie cellule di un organismo,
dipende dalla quantità di DNA
La fase S necessita di una
grande quantità di ATP per la
duplicazione del DNA e la
formazione della cromatina
Anche la fase G2 ha una durata costante, è il
completamento delle sintesi per l’allestimento del
meccanismo molecolare per la mitosi (M)
La fase G1 può durare ore, giorni, settimane o più a lungo,
dipende dal tipo cellulare
Quando la fase G1 si protrae a lungo, si parla di Go, cioè
una fase stazionaria di attesa, oppure di Gz se l’uscita dal
ciclo è irreversibile
Il ritorno da Go a G1 ed il
passaggio da G1 a S
richiede l’intervento di
messaggi ambientali,
chiamati
mitogeni o
fattori di
crescita
Tre tipi di cellule con diversa durata ciclo cellulare:
Cellule molto specializzate che hanno perso la capacità di
dividersi e si trovano in fase Gz, irreversibile. Le cellule
molto specializzate come le cellule nervose, le cellule
muscolari, i globuli rossi hanno perso per sempre la
capacità di dividersi
Cellule differenziate che normalmente non si dividono, ma
lo possono fare in risposta ad uno stimolo appropriato,
per esempio le cellule del fegato che si trovano in fase G0
ma che possono tornare in G1
Cellule attivamente proliferanti come le cellule staminali.
Hanno le fasi G1, S, G2 e Mitosi tutte le cellule che
possono proliferare, cioè le cellule embrionali e le cellule
staminali nei tessuti epiteliali e nel midollo osseo (globuli
rossi e bianchi), le cellule germinali maschili.
Divisione cellulare per
Mitosi
• tipica di tutte le cellule
sia somatiche che
germinali
• da una cellula si
formano due cellule con
lo stesso DNA
=> Mantiene la ploidia
della cellula
Divisione cellulare per Mitosi
• tipica di tutte le cellule
sia somatiche che germinali
• da una cellula si formano
due cellule con lo stesso DNA
=> Mantiene la ploidia della cellula
Quattro Fasi: 1a – Profase; 2a – Metafase; 3a – Anafase; 4a – Telofase
PROFASE :
-duplicazione dei centrioli
-condensazione della cromatina
per formare i cromosomi
-scomparsa di nucleolo e di
membrana nucleare
-i centrioli sono un centro per
l’organizzazione dei microtubuli
che formeranno il FUSO
MITOTICO
METAFASE :
i cromosomi si
allineano
all’equatore della
cellula, agganciati
alle fibre del fuso
mitotico che
partono dai
centrioli
A
N
A
F
A
S
E
TELOFASE :
i cromosomi si
decondensano, si
riforma la
membrana nucleare
e ricompare il
nucleolo
CITODIERESI
Un organismo DIPLOIDE riceve un cromosoma dal
genitore femmina (= materno) ed un cromosoma dal
genitore maschio (= paterno)
Una coppia di
cromosomi
corrispondenti
uno di origine
materna e l’altro
paterna formano
una coppia di
omologhi
Durante la fase S
ogni cromosoma
viene duplicato e
ogni copia completa
(cromatide) resta
unita all’altra per la
zona centrale del
centromero sino alla
fase M
Replicazione del DNA
semi-conservativa
Meccanismo di replicazione
del DNA di Watson e Crick
Il progredire della cellula nel ciclo cellulare dipende dal
superamento di tre checkpoint per entrare:
(1) in fase S,
(2) nella profase della Mitosi, (3) nell’anafase
• Evento fisico: Radiazioni X, g,
UV o particelle cariche di alta
energia ionizzano la materia
biologica cedendo energia
• Evento radiochimico:
Formazione di radicali liberi
• Evento biologico: Danno a
carico di molecole importanti
come DNA, membrane, etc.
Danno al DNA non riparato o mal riparato
MUTAZIONE
La riparazione del DNA danneggiato ha una funzione centrale non
solo nella protezione del genoma ma anche nella generazione della
diversità genetica che caratterizza gli individui di una stessa specie
Riparazione del DNA
ed Evoluzione
Mutazioni e ricombinazione sono
necessari per l’evoluzione, la
velocità di riparazione del DNA
la velocità di evoluzione
influenza
Riparazione efficace significa
evoluzione lenta ma specie stabili
I sistemi di riparazione del DNA negli Eucarioti hanno avuto il massimo
sviluppo nel periodo geologico pre-Cambriano
DNA: insufficiente riparazione del DNA è causa di patologie
invecchiamento cellulare associato ad accumulo di danni
Studi sperimentali sugli animali, in cui erano stati silenziati geni
coinvolti nella riparazione del DNA, hanno evidenziato un
accelerato invecchiamento, una manifestazione precoce di
malattie connesse con l'età avanzata ed una aumentata
suscettibilità a sviluppare
neoplasie
Patologie ereditarie
associate con difetti nella
riparazione del DNA
determinano prematuro
invecchiamento (come la
sindrome di Werner)
Disordini ereditari nella
riparazione del DNA sono
responsabili di diversi
disordini genetici, quali:
•Xeroderma pigmentoso:
ipersensibilità alla luce
solare/UV, che determina
aumentata sensibilità ai
carcinogeni e incidenza di
cancro alla cute ed
invecchiamento precoce
•Sindrome di Cockayne:
ipersensibilità a raggi UV e ad agenti chimici
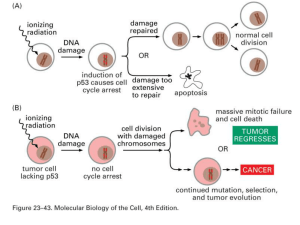
P53 (Tumor Suppressor Gene)
• Mutata (inattivata) in più del 60% di
tutti i tumori umani
• P53 regola (attiva o reprime) la
trascrizione di più di 50 differenti geni
• La quantità di P53 attivata aumenta
rapidamente quando c’è un danno al
DNA o si accumulano prodotti
intermedi della riparazione
APOPTOSI: è una opzione possibile per la Cellula
restare e
funzionare
apoptosi
L’apoptosi si manifesta
con alterazioni tipiche della
cellula che ne sta attuando il
programma genetico
ipertrofia
L’apoptosi viene spesso
attivata in risposta ad una
situazione di danno che
permane e che non
riesce ad essere
compensata
crescere e
dividersi
risposta allo stress
transdifferenziare
dedifferenziare
APOPTOSI
è una morte cellulare programmata, esecuzione di un complesso programma
genetico
l’altra forma di morte cellulare - necrosi – è invece una morte accidentale che si
realizza in modo caotico e senza chiamare in causa trascrizione e traduzione
la cellula in apoptosi si frammenta in corpi apoptotici che sono fagocitati dai
fagociti
APOPTOSI : quando?
nel corso dello sviluppo embrionale/fetale, l’apoptosi rimuove
le cellule indesiderate; per esempio quelle della membrana
interdigitale, circa metà dei neuroni in differenziamento,
i linfociti che possono aggredire il “self”
nei tessuti maturi che vanno incontro a rinnovamento, come le cellule del sangue
e quelle epiteliali, l’apoptosi è l’autoeliminazione delle cellule vecchie e non più
utili
alcune cellule difensive si sacrificano nell’espletamento della loro funzione di
protezione del corpo, ad esempio i granulociti neutrofili o i linfociti
le cellule infettate da virus impediscono la diffusione del contagio ad altre
cellule attivando il programma genetico dell’apoptosi
l’Apoptosi è usata nel continuo ricambio ciclico delle cellule dell’endometrio
uterino, del corpo luteo, dell’epitelio della ghiandola mammaria
Proliferazione
Morte cellulare
Omeostasi
Accumulo
Perdita cellulare
proliferazione di
cellule staminali
proliferazione con
differenziamento
Le cellule di un organismo multicellulare pur avendo gli stessi geni, sono in grado
di rispondere a segnali di differenziamento in modo variabile.
Le cellule sono plastiche, sono in grado di eseguire la loro funzione specializzata
pur mantenendo la capacità di adattarsi ai cambiamenti ambientali.
Questo si realizza attraverso molteplici meccanismi che coinvolgono la regolazione
diretta dello stato della cromatina in risposta a stimoli.
E’ ancora poco conosciuto come le vie di trasduzione del segnale possano
comunicare direttamente con cromatina per cambiare il paesaggio epigenetico, ma
è sempre più evidente la possibilità che la cromatina agisca come sito di
integrazione e di stoccaggio dei segnali.
Nei mammiferi, le informazioni comunicate agli organi sensoriali sono trasmessi al
sistema nervoso centrale, in cui l'informazione viene elaborata e convertita in una
risposta fisiologica.
A livello cellulare, le cascate di signalling trasmettono messaggi tramite
proteine effettrici che raggiungono il nucleo, dove opportuni fattori di trascrizione
attivano/inibiscono l’espressione genica. Questo evento può essere di natura
transitoria, ad esempio quando una cellula deve rispondere ad un evento acuto e poi
tornare al suo precedente stato stazionario.
Emerging roles for chromatin as a signal integration and storage platform - Nat Rev Mol Cell Biol. 2013 Apr;14(4):211-24. doi:
10.1038/nrm3545 - Badeaux AI, Shi Y.
A competitive protein interaction network buffers Oct4-mediated differentiation to promote pluripotency in
embryonic stem cells - Descalzo et al. - Mol Syst Biol. 2013 Oct 8;9:694. doi: 10.1038/msb.2013.49
Nelle cellule staminali embrionali, la
pluripotenza è mantenuta regolando l'attività di
un piccolo insieme di fattori di trascrizione, tra cui
sono particolarmente importanti (Oct4, Nanog,
TCF3, β-catenina) che controllano l'espressione
dell’auto-rinnovamento e dei geni per il
differenziamento
Il blocco dell'attività trascrizionale di Oct4 sembra
essere il principale determinante per uscire dalla
condizione di pluripotenza
From neural development to cognition: unexpected roles for chromatin - Ronan et al. - Nat Rev Genet. 2013 May;14(5):347-59. doi:
10.1038/nrg3413
La specializzazione che le cellule devono acquisire per
formare un organismo da uno zigote viene ottenuta mediante
cambiamenti graduali nell'espressione genica durante lo
sviluppo embrionale. Questi cambiamenti avvengono in
risposta sia a segnali extracellulari che a circuiti genetici
presenti nel nucleo della cellula.
I regolatori della cromatina contribuiscono ai cambiamenti
dinamici di espressione genica, ma anche a mantenere i destini
delle cellule, fornendo stati stabili ed ereditari di espressione
genica. Molti regolatori cromatina sono essenziali per processi
di sviluppo, compreso lo sviluppo del cervello.
Recenti studi di sequenziamento genomico su individui affetti
da disturbi dello sviluppo neurologico e psichiatrico hanno
evidenziato mutazioni in molti regolatori della cromatina.
The Use of Induced Pluripotent Stem Cells in Drug Development - Clin Pharmacol Ther. 2011 May;89(5):655-61. doi:
10.1038/clpt.2011.38 - Inoue H, Yamanaka S.
La tecnologia delle cellule staminali pluripotenti indotte (iPSC) sta
rivoluzionando la scienza medica, permettendo di esplorare i meccanismi di
molte malattie, fornendo nuovi bersagli molecolari terapeutici, e offrendo nuove
opportunità per la scoperta ed il testaggio di farmaci vecchi e nuovi
L’induzione di quattro geni
codificanti per fattori di
trascrizione (Oct3/4, Sox2,
Klf4, c-Myc) basta per
innescare la riprogrammazione
di cellule somatiche umane
e di topo per generare
cellule somiglianti alle rispettive ESC
Esistono già vari altri metodi alternativi per generare iPSC
Le cellule iPSC hanno risolto i problemi di natura etico/morale connessi con la
ricerca nel campo delle cellule staminali embrionali
La tecnologia delle iPSC ci sta mettendo sempre di più nelle condizioni di
sviluppare una efficiente medicina personalizzata
Utilizzando i sarcomi come modello sperimentale abbiamo dimostrato che:
(1) tumori solidi umani dal cariotipo complesso possono essere riprogrammati ad uno stato
pluripotente che soddisfa
tutti i criteri in vitro utilizzati
per definire le cellule staminali
pluripotenti generate da
cellule somatiche;
(2) sarcomi riprogrammati
possono essere differenziati
in tessuto connettivo maturo
e globuli rossi;
(3) la differenziazione terminale
dei sarcomi riprogrammati abolisce
in modo irreversibile il loro
potenziale oncogeno misurato
dalla capacità di ulteriore
proliferazione in vitro o negli
xenotrapianti.
Forse, la conclusione più sorprendente è che la riprogrammazione delle
cellule tumorali dimostra la capacità delle cellule tumorali di impegnarsi
nuovamente e completare l’esecuzione di una normale via di
differenziazione cellulare con conseguente perdita della cancerogenicità.
Reprogramming cancer cells: back to the future - Oncogene, 32, 2247–2248 (2013), doi:10.1038/onc.2012.349; publ online 6 august 2012 - Lang JY, Shi Y & Chin YE
Vascularized and functional human liver from an iPSC-derived organ bud transplant - Nature. 2013 Jul
25;499(7459):481-4. doi: 10.1038/nature12271 - Takebe et al.
La grave carenza di donatori di organi per il trattamento della insufficienza
d'organo in fase terminale rende urgente la necessità di generare organi
da cellule staminali pluripotenti indotte (iPSC).
Fino ad ora nessun laboratorio era riuscito a generare un organo
tridimensionale vascolarizzato come il fegato.
Si dimostra la generazione di un fegato umano vascolarizzato e funzionale
ottenuto dal trapianto di gemme di fegato formate da iPSC umane create in
vitro. Queste gemme si collegano ai vasi sanguigni entro 48 ore dal
trapianto e crescono portando alla formazione di tessuto simile al fegato
adulto che presentava le funzioni fegato-specifiche, come la produzione di
proteine e il metabolismo dei farmaci, con la possibilità di rendere non
necessaria la sostituzione del fegato del destinatario.
Questo è il primo lavoro che dimostra la generazione di un organo umano
funzionale da cellule staminali pluripotenti. Ulteriori sforzi dovranno essere
effettuati per tradurre queste tecniche in trattamenti per i pazienti, un
nuovo promettente approccio per la medicina rigenerativa.
Vascularized and functional human liver from an iPSC-derived organ bud transplant - Nature. 2013 Jul 25;499(7459):481-4.
doi: 10.1038/nature12271 - Takebe et al.
Generazione di gemme di fegato umano da
iPSC umane:
(a) Schema del disegno sperimentale;
(b) auto-organizzazione tridimensionali di
umane in colture di umana;
(c) confronto tra quando ottenuto dalle iPSC
(pannello superiore) rispetto alle culture
Generazione di fegato
umano vascolarizzato
in vivo
iPSC
umane
bidimensionali convenzionali
(pannello inferiore)
(d) presenza di reti endoteliali
nascenti all'interno delle gemme di
iPSC umane;
(e) analisi di PCR quantitativa di
espressione dei geni marcatori
epatici al sesto giorno di coltura.
Caratterizzazione funzionale
di gemme di iPSC di fegato
umano
Transcription factor-mediated reprogramming of fibroblasts to expandable, myelinogenic oligodendrocyte
progenitor cells - Nature Biotechnology, 31, 5 may 2013, 426-433 - Najm FJ, Lager AM, Zaremba A, Wyatt K, Caprariello
AV, Factor DC, Karl RT, Maeda T, Miller RH & Tesar PJ
Le terapie cellulari per le malattie della mielina, come la sclerosi
multipla e le leucodistrofie, necessitano di tecnologie per la
generazione di cellule progenitrici degli oligodendrociti funzionali. In
questo lavoro si descrive la conversione diretta di fibroblasti
embrionali e polmonari murini a cellule progenitrici degli
oligodendrociti indotte (iOPC) utilizzando set di otto o tre ben definiti
fattori di trascrizione. Le iOPC presentano una morfologia bipolare ed
un profilo di espressione genica globale coerente con le vere OPC. Le
iOPC possono essere espanse in vitro per almeno cinque passaggi,
mantenendo la capacità di differenziarsi in oligodendrociti.
Quando sono trapiantate in topi ipomielinati, le iOPC sono in grado
di inguainare gli assoni dell’ospite e di generare mielina compatta.
La conversione di linee di cellule somatiche a iOPC espandibili fornisce
una strategia per studiare il controllo molecolare della identità delle linee
degli oligodendrociti e può facilitare la costruzione di modelli animali per
lo studio delle malattie neurologiche e per arrivare a mettere a punto
terapie rimielinanti autologhe.
Figura 1 - ………….. (a) schema del disegno sperimentale e sviluppo temporale per la
riprogrammazione mediata da di fibroblasti Plp1-eGFP e R26-M2rtTA per formare iOPC,
espandibili ed in grado di differenziare in iOL. Il transgene di Plp1-eGFP modificato è
espresso sia nelle iOPC che negli oligodendrociti in vivo ……...
Il potenziale delle terapie cellulari per le malattie della mielina si basa sulla capacità di
generare cellule autologhe mielinogeniche da trapiantare.
La fonte cellulare più promettente per tali terapie sono le OPC. Gli oligodendrociti maturi
sono molto poco efficaci nel rimielinare gli assoni dell’ospite dopo il trapianto. Sebbene le
cellule staminali neurali e le cellule staminali neuronali indotte possano generare
oligodendrociti, l'efficienza di questo processo è piuttosto basso, e le cellule hanno una
propensione a formare neuroni ed astrociti. Al contrario, le iOPC appaiono limitate alla
generazione degli oligodendrociti. Abbiamo dimostrato che le iOPC si integrano nel
sistema nervoso centrale e mielinizzano gli assoni di topi congenitamente demielinati dopo il
trapianto in vivo. Tuttavia, affinché le iOPC possano avere rilevanza clinica, gli studi futuri
dovranno estendere questa strategia di riprogrammazione a cellule somatiche umane e
dimostrare una estesa mielinizzazione nel CNS ed un beneficio funzionale a lungo
termine nei destinatari del trapianto.
Transcription factor-mediated reprogramming of fibroblasts to expandable, myelinogenic oligodendrocyte progenitor cells - Nature
Biotechnology, 31, 5 may 2013, 426-433 - Najm et al.
Stem cells: A time to heal - Cassandra Willyard - Nature 503, S4–S6 (14 November 2013) doi:10.1038/503S4a Published online 13 November 2013
Le prime terapie con cellule staminali per le lesioni del midollo
spinale sono già in fase di sperimentazione in studi clinici, ma
restano l’incertezza scientifica e politica.
Regenerative medicine: Rebuilding the backbone - Mike May - Nature 503, S7–S9 (14 November 2013)
doi:10.1038/503S7a Published online 13 November 2013
I chirurghi possono aiutare a
fissare le vertebre danneggiate,
ma potrebbe un'infusione di
cellule in un materiale
bioingegnerizzato crescere per
sostituire una colonna
vertebrale danneggiata?
Developmental Neuroscience: Miniature human Brains - Nature. 2013 Sep 19;501(7467):319-20. doi: 10.1038/nature12552 Brüstle O.
Le cellule staminali indotte possono essere utilizzate per formare tessuti tridimensionali
che imitano il cervello umano in sviluppo. Questi tessuti forniscono un mezzo importante
per lo studio dei disturbi dello sviluppo neuronale. Cellule staminali embrionali
pluripotenti possono differenziarsi in tutti i tipi di cellule del corpo. Questo processo può
anche essere ottenuto in vitro, usando sia staminali pluripotenti (ES) che staminali
indotte (iPSC) ottenute dalla riprogrammazione di cellule epiteliali differenziate.
In condizioni adeguate, le iPSC umane possono auto-organizzarsi in organoidi cerebrali
che ricreano i primi passi della
formazione della corteccia cerebrale
del cervello umano , e quindi si prestano
a studi sullo sviluppo del cervello
e sui disturbi dello sviluppo neurale.
Recentemente era stata dimostrata la
formazione di tratti di corteccia cerebrale,
di intere strutture oculari nei primi stadi di
sviluppo ed anche retine stratificate a
partire da cellule umane e d murine.
Figura in alto: generazione di organoidi da cellule staminali umane pluripotenti che possono essere
derivate sia ES (cellule embrionali dalla massa cellulare interna della blastocisti di embrioni a 70200 -cellule) che staminali pluripotenti (iPS) indotte attraverso la riprogrammazione di cellule adulte.
Le cellule neurali derivate da cellule staminali pluripotenti si auto-organizzano in organoidi cerebrali
con architetture dei tessuti che ben ricordano la corteccia cerebrale umana.
Developmental Neuroscience: Miniature human Brains - Nature. 2013 Sep 19;501(7467):319-20. doi: 10.1038/nature12552
- Brüstle O.
Figura a sinistra: organoide umano generato in vitro in cui si vedono i
neuroni (fluorescenza verde e le cellule progenitrici (fluorescenza
magenta). La morfologia della regione superiore destra corrisponde a
quella della corteccia cerebrale e la regione in basso a sinistra ha una
morfologia simile a quella del plesso coroide. All’interno è presente una
cavità piena di liquido.
La corteccia cerebrale umana è evolutivamente il tessuto più
complesso del regno animale. Come nel cervello nativo, le aree
corticali segregano in diversi strati e queste peculiarità dello sviluppo corticale umano sono ben
riassunte negli organoidi.
Negli organoidi cerebrali le aree imitano le diverse regioni del cervello ma distribuite
casualmente, non hanno la forma e la complessiva organizzazione spaziale che si vede nel
cervello e sono privi di un sistema circolatorio che impedisce di avanzare alla complessa
architettura a sei livelli della loro controparte naturale.
Anche se con queste limitazioni, le potenziali applicazioni di tale modello in vitro sono
affascinanti e gli organoidi cerebrali possono fornire un modello per studiare i meccanismi di
sviluppo iniziale del cervello umano in laboratorio. Tali studi possono essere estesi ai disturbi
dello sviluppo neurale. Sono state utilizzate cellule iPSC derivate da fibroblasti della pelle di un
paziente con una forma genetica di microcefalia (disordine dello sviluppo neurale caratterizzato
da una sostanziale riduzione delle dimensioni del cervello che è difficile da studiare in modelli
animali a causa delle differenze di specie nello sviluppo del cervello). Rispetto agli organoidi
derivati da pazienti sani, quelli derivati da pazienti microcefalici avevano un minor numero di
cellule progenitrici e un aumento del numero di neuroni differenziati. Questa osservazione
indica il precoce differenziamento neuronale come meccanismo alla base di questa forma di
microcefalia.
The Use of Induced Pluripotent Stem Cells in Drug Development -Clin Pharmacol Ther. 2011 May;89(5):655-61. doi:
10.1038/clpt.2011.38 - Inoue H, Yamanaka S.
Medicina personalizzata
Il vantaggio evidente di
utilizzare iPSC piuttosto
che ESC è che le iPSC
possono essere
derivate da qualsiasi
individuo con relativa
facilità, consentendo in
tal modo lo sviluppo
di una piattaforma di
studio personalizzato
sul singolo genoma.
iPSC e cellule differenziate
da iPSC mantengono la loro identità personale, come un alter ego del paziente,
e possono essere usate per studiare le sue malattie con approccio personaspecifico per esaminare le differenze individuali nella risposta ai farmaci (vedi figura
in alto).