Biosintesi dei carboidrati
Gluconeogenesi: sintesi di glucosio da precursori non saccaridici
La riserva di glucosio dell’organismo (glucosio + glicogeno) è
sufficiente per circa 1 giorno.
La via gluconeogenetica converte il piruvato in glucosio.
I precursori non glucidici del glucosio vengono prima convertiti in piruvato o
entrano nella via a livello di intermedi successivi come l’ossalacetato e il
diidrossiacetone fosfato.
I principali precursori non glucidici sono il lattato, gli amminoacidi e il glicerolo.
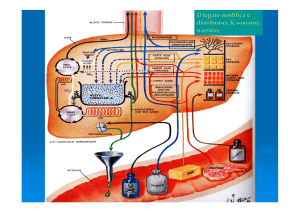
La gluconeogenesi ha luogo principalmente nel fegato, e in piccola quantità anche
nel rene.
La gluconeogenesi che avviene nell’encefalo, nel muscolo scheletrico e nel muscolo
cardiaco è solo di modesta entità.
La principale funzione della gluconeogenesi nel fegato e nel rene è di mantenere i
livelli ematici di glucosio sufficientemente alti da consentire all’encefalo e al
tessuto muscolare di avere sufficienti quantità di glucosio per soddisfare le
proprie esigenze metaboliche
Sintesi di carboidrati da precursori semplici
La via della gluconeogenesi
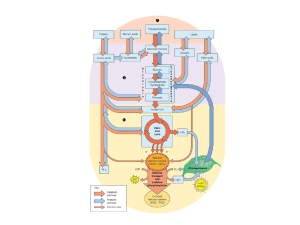
Confronto tra le vie glicolitica e gluconeogenetica
Il fosfoenolpiruvato si forma in due reazioni.
1) Il piruvato viene prima carbossilato ad ossalacetato a spese di 1 molecola di
ATP
piruvato + HCO3- + ATP Æ ossalacetato + ADP + Pi + H+
enzima: piruvato carbossilasi
- Questa è anche una reazione anaplerotica del ciclo di Krebs
2) l’ossalacetato viene poi decarbossilato e fosforilato a spese di un secondo
legame fosforico ad alta energia (GTP)
ossalacetato + GTP ' fosfoenolpiruvato + CO2 + GDP
enzima: fosfoenolpiruvato carbossichinasi
Il fosfoenolpiruvato si forma dal piruvato attraverso l’ossalacetato
Struttura a domini della piruvato carbossilasi
La biotina è un gruppo prostetico legato
covalentemente all’enzima, che serve da
trasportatore di CO2 attivato.
La carbossilazione del piruvato si svolge in tre stadi:
1) HCO3- + ATP ' Carbossifosfato + ADP
2) Enzima-biotina + Carbossifosfato ' CO2-enzima-biotina + Pi
3) CO2-enzima-biotina + piruvato ' enzima-biotina + ossalacetato
Il dominio di legame per la biotina della piruvato carbossilasi
La biotina è su un “guinzaglio flessibile” che le permette di muoversi tra
il sito dell’ATP e del bicarbonato e il sito del piruvato.
Il fosfoenolpiruvato si forma in due reazioni.
piruvato + HCO3- + ATP + GTP Æ fosfoenolpiruvato + ADP + GDP
+ Pi + H+ + CO2
ΔG0’ = 0,9 kj/mole
Per fosforilare 1 molecola di piruvato a fosfoenolpiruvato sono necessari 2 gruppi
fosforici ad alta energia
La piruvato carbossilasi è un enzima
mitocondriale mentre gli altri enzimi della via
gluconeogenetica sono citosolici.
L’ossalacetato viene trasportato nel citosol
sotto forma di malato.
Con il malato viene trasportato anche NADH.
2) Il fruttosio 6-fosfato si forma dal fruttosio 1,6-bisfosfato per idrolisi
dell’estere fosforico sull’atomo di carbonio C-1.
Fruttosio 1,6-bisfosfato + H2O Æ fruttosio 6-fosfato + Pi
ΔG0’ = -16,3 kj/mole
enzima: fruttosio 1,6 bisfosfatasi
3) Il glucosio libero si forma per idrolisi del glucosio 6-fosfato
glucosio 6-fosfato + H2O ¿ glucosio + Pi
ΔG0’ = -13,8 kj/mole
enzima: glucosio 6 fosfatasi
La glucosio 6-fosfatasi è presente nel fegato ma non nel muscolo e nel cervello.
Differenze enzimatiche tra la glicolisi e la gluconeogenesi
Glicolisi
Gluconeogenesi
Esochinasi
Glucosio 6-fosfatasi
Fosfofruttochinasi
Fruttosio 1,6-bisfosfatasi
Piruvato chinasi
Piruvato carbossilasi
Fosfoenolpiruvato carbossichinasi
La stechiometria della gluconeogenesi
2 piruvato + 4 ATP + 2 GTP + 2 NADH + 4 H2O ¿ glucosio + 4 ADP + 2 GDP
+ 6 Pi + 2 NAD+ + 2 H+
ΔG0’ = --9 kcal/mole
Per sintetizzare 1 molecola di glucosio dal piruvato vengono usati 6 legami
fosforici ad alta energia + 2 molecole di NADH
Nella glicolisi durante la conversione del glucosio a piruvato vengono prodotte
solo 2 molecole di ATP
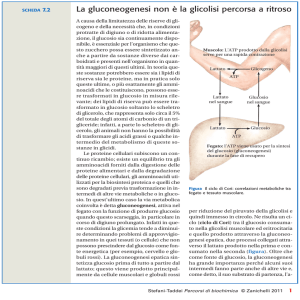
L’alanina ed il lattato prodotti dal muscolo scheletrico attivo sono uno dei
principali rifornimenti della gluconeogenesi.
Nel muscolo in attività l’elevato rapporto
NADH/NAD+ porta alla formazione
prevalente di lattato.
Nel fegato il lattato viene ossidato a
piruvato
per
il
basso
rapporto
NADH/NAD+ presente nel citosol degli
epatociti.
Il ciclo di Cori
La formazione del lattato nel muscolo sposta parte del carico metabolico dal
muscolo al fegato
Gli intermedi del ciclo dell’acido citrico e molti amminoacidi sono glucogenici
L’idrolisi dei triacilgliceroli di deposito produce anche glicerolo 3-fosfato, che può
entrare nella via gluconeogenetica dopo essere stato ossidato a diidrossiacetone fosfato
Ciclo dell’acido citrico
Ciclo del gliossilato
Al contrario degli animali, le piante possono
usare i grassi per sintetizzare glucosio
Le riserve di acidi grassi conservati nei semi sono convertite in saccarosio durante la
germinazione
La biosintesi del glucosio viene attivata
quando nei mitocondri vi è un eccesso di
Acetil-CoA.
Un eccesso di Acetil-CoA stimola la
piruvato carbossilasi.
L’Acetil-CoA aumenta quando la velocità
di produzione supera la velocità di
utilizzazione nel ciclo dell’acido citrico.
Il ciclo dell’acido citrico è inibito quando le
richieste energetiche della cellula sono
soddisfatte.
Il secondo punto di controllo della gluconeogenesi è la reazione catalizzata
dalla fruttosio 1,6 - bisfosfatasi.
L’enzima glicolitico corrispondente è la fosfofruttochinasi-1.
La regolazione ormonale (glucagone) della glicolisi e della gluconeogenesi nel
fegato è mediata dal fruttosio 2,6 bisfosfato.
Il fruttosio 2,6 bisfosfato attiva la fosfofruttochinasi 1 e stimola la glicolisi nel
fegato.
Il fruttosio 2,6 bisfosfato inibisce la fruttosio 1,6 bisfosfatasi e rallenta la
gluconeogenesi nel fegato.
Ruolo del fruttosio 2,6 bisfosfato nella regolazione della glicolisi e della gluconeogenesi.
Regolazione del livello di fruttosio 2,6- bisfosfato.
La concentrazione cellulare del modulatore fruttosio 2,6-bisfosfato viene determinata
dalla velocità della sua sintesi da parte della fosfofruttochinasi-2 (PFK-2) e dalla
velocità della sua degradazione operata dalla fruttosio 2,6-bisfosfatasi (FBPase-2).
Regolazione del livello di fruttosio 2,6- bisfosfato.
La fosfofruttochinasi-2 (PFK-2) e la fruttosio 2,6-bisfosfatasi fanno parte della stessa
catena polipeptidica (enzima tandem) e sono regolati in modo complementare e
coordinato dal glucagone.
Regolazione reciproca della gluconeogenesi e della glicolisi nel fegato.
Ciclo di Crebs se
l’ATP è scarso
Glucosio abbondante = ½ Fruttosio 2,6 bisfosfato = Glicolisi ½, Gluconeogenesi ¾
Digiuno prolungato = ¾ Fruttosio 2,6 bisfosfato = Glicolisi ¾, Gluconeogenesi ½
Biosintesi del glicogeno
Molte delle reazioni in cui gli esosi vengono trasformati o polimerizzati
utilizzano zuccheri legati a nucleotidi.
Formazione di uno zucchero legato a nucleotidi.
Sintesi del glicogeno
La catena del glicogeno viene allungata dalla glicogeno sintasi. Il residuo di
glucosio UDP-glucosio viene trasferito all’estremità non riducente di una
catena di glicogeno mediante un nuovo legame (α1Æ4).
Formazione di ramificazioni nella molecola di glicogeno
La glicogeno sintasi non può produrre i legami(α1Æ6) presenti ai punti di
ramificazione della molecola di glicogeno.
I legami(α1Æ6) vengono formati dall’enzima ramificante chiamato amilo
(1Æ4)(1 Æ6) transglicosilasi oppure glicosil-(4 Æ6)-transferasi, che catalizza
il trasferimento di un segmento terminale di 6 o 7 residui glucosidici
dall’estremità non riducente di una catena di glicogeno che abbia almeno 11
residui, al gruppo ossidrilico C-6 di un residuo di glucosio della stessa o di
una’altra catena localizzato in un punto più interno .
La glicogeno sintasi può quindi aggiungere altri residui glucosidici alla nuova
ramificazione
La glicogeno sintasi ha bisogno come
innesco (primer) di una catena di
poliglucosio (α1Æ4) preformata con
una lunghezza di almeno 8 residui
La glicogenina, una proteina, può
comportarsi da primer a cui legare il primo
residuo di glucosio (in corrispondenza del
residuo Tyr194) e anche da catalizzatore per
la sintesi di una catena nascente di
glicogeno.
La glicogenina forma poi un complesso
stabile con la glicogeno sintasi.
La glicogenina continua a far parte della
particella di glicogeno restando legata
covalentemente all’estremità a cui ha dato
origine.
La sintesi di una particella di glicogeno da parte della glicogenina
Regolazione coordinata della glicogeno sintasi e della glicogeno
fosforilasi
La fosforilazione ha effetti opposti sull’attività enzimatica della glicogeno
sintasi e su quella della glicogeno fosforilasi
Livelli di glucosio ematici bassi:
Glicogenolisi
+
+ glucosio in circolo
Gluconeogenesi
+
Glicolisi
-
Glicogeno sintesi
-
Livelli di glucosio ematici alti:
Glicogenolisi
-
Gluconeogenesi
-
Glicolisi
+
Glicogeno sintesi
+