
26/05/2013
FUNZIONE MICROTUBULI
(G. Karp: Biologia cellulare e Animale, EdiSES, 1998)
Citoscheletro
Scheletro interno o impalcatura che fornisce un
supporto strutturale e aiuta il mantenimento
della posizione degli organelli citoplasmatici
Parte del meccanismo che sposta materiali e
organelli da una parte all’altra della cellula
Elementi motori di ciglia e flagelli
Componenti principali del meccanismo
responsabile della mitosi e della meiosi (fuso
mitotico)
Microtubuli
BUSB
1
Microtubuli e organizzazione del citoplasma
‐2
2
Microtubuli: organizzazione di assoni e dendriti
http://scientia.wikispaces.com/Orga
nization+of+the+Nervous+System
(A) Schema di una cellula illustrando la tipica disposizione dei microtubuli (verde), reticolo
endoplasmatico (blu) e apparato di Golgi (giallo). Il nucleo è evidenziato in marrone e il
centrosoma in verde chiaro.
(B) Cellula marcata con anticorpi contro il reticolo endoplasmatico (panello superiore) o
microtubuli (panello inferiore). Proteine motore tirano il RE lungo i microtubuli,
estendendolo come un reticolato a partire dal suo collegamento all’involucro
nucleare.
(C) Cellula marcata con anticorpi contro l’apparato di Golgi (panello superiore) o contro i
microtubuli (panello inferiore). In questo caso delle proteine motore muovono l’apparato
di Golgi verso l’interno fino alla sua posizione vicino al centrosoma.
http://www.ncbi.nlm.nih.gov/books/NBK28420/figure/A4196/?report=objectonly
Microtubuli: Flagelli e cilia
http://www.ncbi.nlm.nih.gov/books/NBK28122/figure/A574/
Microtubuli: Eucarioti unicellulari ciliati
http://kentsimmons.uwinnipeg.ca/cm1504/cytoskeleton.htm
1
26/05/2013
Cellule ciliate dell’epitelio respiratorio
http://faculty.une.edu/com/abell/histo/histolab3c.htm
Schema dell’epitelio respiratorio
cellule ciliate
http://alexandria.healthlibrary.ca/documents/notes/bom/unit_
2/L32%20Histology%20of%20the%20Respiratory%20System
%201.xml
http://greaterimmunity.com/Files/respiratory_i
mmunity.html
Microtubuli: Cellule eucariotiche flagellate -2
Microtubuli: Cellule eucariotiche flagellate ‐1
Fuso mitotico ‐ 1
Microtubuli
Polimerizzazione
http://ehumanbiofield.wikispaces.com/Chromosomes+HW4+MC
12
2
26/05/2013
Polimerizzazione dei MT
http://www.ncbi.nlm.nih.gov/books/NBK21522/figure/A5432/
Microtubuli_comp2011Microtubuli
Microtubuli
13
Cinetica di crescita dei Microtubuli
INSTABILITA’ DINAMICA DEI MICROTUBULI
http://www.ncbi.nlm.nih.gov/books/NBK26862/?rendertype=figure&id=A2975
17
18
3
26/05/2013
L’instabilità dinamica è dovuta alle differenza
strutturali fra le estremità in crescita e in accorciamento ‐ 1
L’instabilità dinamica è dovuta alle differenza
strutturali fra le estremità in crescita e in accorciamento ‐ 2
Se la concentrazione libera di tubulina in soluzione si trova fra i valori critici per l’estremità positiva e quella negativa, l’estremità di un singolo microtubulo può subire transizioni fra uno stato di accrescimento e uno di accorciamento. Un microtubulo in accrescimento ha subunità contenenti GTP alla sua estremità, formando un cappello a GTP. Se l’idrolisi del nucleotide ha luogo più rapidamente dell’aggiunta di una nuova subunità, il cappello viene perso e il microtubulo inizia ad accorciarsi, un evento chiamato “catastrofe”.
Ma in ogni caso continueranno ad aggiungersi all’estremità in accorciamento delle subunità con GTP, e se un numero sufficiente di subunità viene aggiunto e si forma un nuovo cappello, la crescita del microtubuli riprende, un evento detto “salvataggio”.
Modello per le consequenze strutturali dell’idrolisi dell’GTP sulla rete di microtubuli. L’aggiunta di subunità di tubulina contenente GTP all’estremità di un protofilamento provoca la crescita in conformazione lineare che si può rapidamente impacchettare nella parete cilindrica del microtubulo. L’idrolisi del GTP dopo l’assemblaggio provoca l’alterazione delle conformazione delle subunità e tende a forzare i protofilamenti ad assumere una forma incurvata che è meno suscettibile ad impacchettarsi nella parete del microtubulo.
19
L’instabilità dinamica è dovuta alle differenza strutturali
fra le estremità in crescita e in accorciamento ‐ 3
In un microtubulo intatto, i protofilamenti fatti da subunità contenenti GDP sono costretti in una conformazione lineare dai diversi legami laterali all’interno della parete del microtubulo, a causa della presenza di un cappello stabile di subunità contenenti GTP. Tuttavia, la perdita del cappello di GTP permette alle subunità contenenti GDP di rilassarsi ad assumere la loro conformazione più incurvata. Ciò porta al progressivo disassemblamento del microtubulo. Sopra gli schemi di un microtubulo in crescita e in accorciamento, le fotografie in microscopia elettronica mostrano reali microtubuli in ciascuno di questi stati. Notare in particolare l’incurvatura, la disintegrazione dei protofilamenti contenentiGDP ad’una estremità del microtubulo in accorciamento.
Microtubuli
Proteine associate
21
“Microtubule‐associated proteins” (MAPs)
Sono proteine che si legano lateralmente ai microtubuli.
Come il farmaco tassolo, le MAPs possono stabilizzare i microtubuli contro il disassemblamento.
Un sotto‐insieme di MAPs può anche mediare le interazioni dei microtubuli con altre componenti cellulari.
o Queste MAPs sono cospicue nei neuroni, dove stabilizzano i fasci di microtubuli che formano l’asse centrale degli assoni e dei dendriti che si stendono dal corpo cellulare.
o La lunghezza del dominio che si proietta dal microtubulo può determinare quanto vicino i microtubuli rivestiti di MAPs si impachettano.
24
4
26/05/2013
Cell
overexpressing tau
Cell
overexpressing
MAP2
25
Citoscheletro degli assoni e dei dendriti
http://www.ncbi.nlm.nih.gov/books/NBK26809/figure/A3012/
26
Didascalia fig. Citoscheletro assone e dendriti ‐ 1
I citoscheletri degli assoni e dei dendriti differiscono sia in composizione che in organizzazione. Con un eccezione, tutte le proteine citoscheletriche sono sintetizzate su polisomi liberi nel corpo cellulare, e successivamente trasportate ai loro diversi compartimenti cellulari.
L’eccezione è la MAP2, che è la principale proteina associate ai microtubuli dei dendriti. Mentre una certa quantità di MAP‐2 è sintetizzata nel corpo cellulare, il mRNA della MAP2 è specialmente abbondante nel compartimento dendritico e si ritiene che una frazione significativa venga sintetizzata in quella sede. Si pensa che i microtubuli dei corpi cellulari, dendriti e assoni vengano nucleati nel “microtubule‐organizing center”(MTOC), e successivamente rilasciati nei dendriti o nell’assone.
http://www.ncbi.nlm.nih.gov/books/NBK28122/figure/A574/
Didascalia fig. Citoscheletro assone e dendriti ‐ 2
Didascalia fig. Citoscheletro assone e dendriti ‐ 3
Nel dendrite, i microtubuli spesso hanno polarità miste, con estremità sia (+) che (‐) distali rispetto al corpo cellulare. La conseguenza funzionale di tale organizzazione è incerta ma potrebbe spiegare perchè la forma dei dendriti si affusoli sempre di pù man mano essi si allontano dal corpo cellulare.
La MAP2 sembra essere assente dall’assone.
Viceversa, i microtubuli assonali sono orientate con la loro estremità (+) situate distalmente al corpo cellulare e mostrano una distribuzione uniforme lungo l’assone.
Sia i microtubuli che i neurofilamenti si fermano ed iniziano nell’assone piuttosto che proseguire all’interno verso il corpo cellulare.
Nonostante una certa quantità di proteina tau possa trovarsi nei corpi cellulari e nei dendrite, i microtubuli assonali hanno una gran quantità di proteina tau e la tau assonale è fosforilata diversamente.
I microfilamenti presentano un’organizzazione pù dispersa e possono essere difficilmente visualizzabili nel neurone maturo. Essi sono più abbondanti vicino alla membrana plasmatica e sono inoltre abbondanti nei terminali presinaptici e nelle spine dendritiche.
I neurofilamenti (filamenti intermedi) vengono largamente esclusi dai compartimenti dendritici ma sono abbondanti nei grandi assoni. La spaziatura dei neurofilamenti è sensibile al livello di fosforilazione,
GA: apparato di Golgi. 5
26/05/2013
Microtubuli
Proteine motore
http://neurologicalcorrelates.com/wordpress/wp-content/uploads/2008/01/tangles_big.jpg
31
34
Role of cytoskeletal motors beyond membrane transport.
From the following article:
Molecular motors
Manfred Schliwa and Günther Woehlke
Nature 422, 759-765(17 April 2003)
1, Retrograde transport of centrosomal
components.
2, Anterograde and retrograde
transport of intermediate filaments.
3, Anterograde and retrograde
transport of ribonucleoprotein (RNP)
complexes.
4, Myosin, kinesin and dynein motors
interact with components of the
microtubule plus-end complex
5, Anchorage of dynein at the actinrich cell cortex.
6, Interaction of a kinesin-like protein
with actin.
7, Catenin-mediated anchorage of
dynein at adherens junctions.
35
http://www.nature.com/nature/journal/v422/n6933/fig_tab/nature01601_F5.html
36
6
26/05/2013
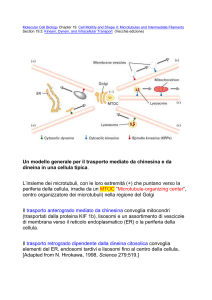
Modello generale per il trasporto mediato da chinesina e da dineina in una cellula tipica
Movimento assonale ‐ 1
L’insieme dei microtubuli, con le loro estremità (+) che puntano verso la periferia della cellula, irradia da un MTOC “Microtubule‐
Organizing Center”, centro organizzatore dei microtubuli) nella regione del Golgi.
Il trasporto anterogrado mediato da chinesina convoglia mitocondri (trasportati dalla proteina KIF 1b), lisosomi e un assortimento di vescicole di membrana verso il reticolo endoplasmatico (ER) o la periferia della cellula.
Il trasporto retrogrado dipendente dalla dineina citosolica convoglia elementi del ER, endosomi tardivi e lisosomi fino al centro della cellula.
Molecular Cell Biology Chapter 19. Cell Motility and Shape II: Microtubules and Intermediate Filaments Section 19.3. Kinesin, Dynein, and Intracellular
Transport (Vecchia edizione)
Un tipico neurone di Vertebrato La freccia indica la direzione in cui i segnali vengono convogliati. Il singolo assone conduce i segnali dal corpo cellulare all’esterno, mentre i multipli dendriti ricevono segnali dagli assoni di altri neuroni. I terminali nervosi finiscono sui dendriti o sul corpo cellulare di altri neuroni o di altri tipi cellulari come ad esempio il muscolo o le cellule ghiandolari. 37
38
Movimento assonale veloce
Didascalia figura precedente ‐ 1
Didascalia figura precedente ‐ 2
Illustrazione schematica di materiale associato a membrane nel trasporto assonale veloce. Il movimento assonale veloce rappresenta il movimento di organelli circondati da membrane lungo i microtubuli assonali sia in direzione anterograda che retrograda. Sono illustrati due principali classi di organelli racchiusi da membrane che sono sintetizzati e impacchettati mediante vie diverse. I polipeptidi delle vescicole sinaptiche sono tradotti su ribosomi legati al reticolo endoplasmatico con un processo in cui le proteine di membrana sono orientate in modo corretto all’interno del doppio strato lipidico e i polipeptidi da secrezione entrano nel lume del reticolo endoplasmatico. Questi polipeptidi sono ulteriormente processati all’interno dell’apparato di Golgi, dove hanno luogo le modificazioni appropriate post‐traduzionali e lo smistamento dei polipeptidi destinati all’assone. Una volta che questi polipeptidi sono impacchettati in organelli vescicolari, e le molecole motore appropriate sono presenti, gli organelli sono trasportati lungo l’assone utilizzaanfo i microtubuli assonali come rotaie a velocità di 200‐400 nm al giorno. Si ritiene che il movimento in direzione anterograda sia mediato dal motore molecolare chinesina, mentre la forza necessaria per muovere in verso retrogrado gli organelli dovrebbe essere generata dalla dineina citoplasmatica. Al contrario della sintesi dei polipeptidi vescicolari, i polipeptidi mitocondriali, che sono codificati dal genoma nucleare, sono sintetizzati su ribosomi citoplasmatici e contengono una sequenza “target” che indirizza i polipeptidi ai mitocondri. In seguito all’assemblaggio, i mitocondri si muovono lungo l’assone a velocità di circa 50‐100 mm al giorno. E’ possibile anche individuare mitocondri che si muovono indietro verso il corpo cellulare, ossia in direzione retrograda. La morfologia dei mitocondri trasportati in modo retrogrado è nettamente diversa da quelle dei mitocondri che si muovono in direzione anterograda e si pensa che essi rappresentino organelli degeneranti che non sono metabolicamente attivi. RER: reticolo endoplasmatico ruvido (“rough”). 41
7
26/05/2013
Movimento assonale lento
Didascalia figura precedente ‐ 1
Illustrazione schematica del movimento degli elementi citoscheletrici nel trasporto assonale lento. Il trasporto assonale lento rappresenta il movimento di costituenti citoplasmatici, incluso elementi citoscheletrici e enzimi solubili del metabolismo intermediario, a velocità di 0.2‐2 mm al giorno, che sono almeno due ordini di grandezza (102, ossia 100 volte) più lenti di quelli osservati nel trasporto assonale veloce.
Basic Neurochemistry Part Four. Growth, Development and Differentiation 28. Axonal Transport Discovery and
Conceptual Development of Fast and Slow Axonal Transport
44
Didascalia figura precedente ‐ 2
Come proposto nell’ipotesi strutturale e supportato da prove sperimentali, si presume che i componenti del citoscheletro siano trasportati lungo l’assone in forme polimerica, e non come subunità polipeptidiche individuali. I polipeptidi del citoscheletro sono tradotti su polisomi citoplasmatici e quindi assemblati in polimeri prima del trasporto lungo l’assone in direzione anterograda. Contrariamente al trasporto assonale veloce, nessun costituente del trasporto lento sembra essere trasportato in direzione retrograda. Nonostante la composizione dei polipeptidi nel trasporto assonale lento sia stata estesamente caratterizzata, le proteine motore responsabili del movimento di questi componenti citopalsmatici non sono ancora state identificate. Microtubuli
Centrosoma, centrioli
46
CENTROSOMA: Centro principale di organizzazione dei microtubuli (MTOC) nelle cellule animali. Comprende due centrioli circondati da una matrice elettron‐densa, il materiale pericentriolare (PCM). CENTRIOLO: Il centriolo canonico è un cilindro che comprende 9 triplette di microtubuli, ha circa ~0.5 μm di lunghezza e ha appendici nelle estremità distali dopo maturazione. Vi sono variazioni in questa struttura, in cui le triplette sono sostituite da singolette o doppiette e non vi sono appendici.
CORPO BASALE: Una struttura che si trova alla base delle cilia e flagelli degli eucarioti che organizza l’assemblaggio dell’assonema. I centrioloi possono dare origine a corpi basali o vice versa. La struttura del corpo basale è la stessa del centriolo; inoltre, i corpi basali hanno una zona di transizione nell’estremità distale che è contigua all’assonema. MATERIALE PERICENTRIOLARE. Materiale fibrillare che circonda I centrioli nel centrosoma e che nuclea la crescita di nuovi microtubuli. 8
26/05/2013
MICROTUBULE ORGANIZING CENTERS (MTOCs)
(Centri di Organizzazione dei Microtubuli)
Centrosoma
Controllano il numero e la polarità dei microtubuli che si formano
Controllano il numero di protofilamenti che formano la parete dei microtubuli (ad es. 13 o 11, come nell’assonema o nei centrioli e corpi basali)
Controllano il momento e il luogo dove i microfilamenti vengono assemblati
Hanno in comune la proteina γ‐tubulina nel materiale pericentriolare, critica per l’assemblaggio dei microtubuli
G. Karp: Biologia cellulare e Animale, EdiSES, 1998
http://www.ncbi.nlm.nih.gov/books/NBK26809/figure/A2999/?report=objectonly
49
MICROTUBULE ORGANIZING CENTERS (MTOCs)
(Centri di Organizzazione dei Microtubuli)
Controllano il numero e la polarità dei microtubuli che si formano
Controllano il numero di protofilamenti che formano la parete dei microtubuli (ad es. 13 o 11, come nell’assonema o nei centrioli e corpi basali)
Controllano il momento e il luogo dove i microfilamenti vengono assemblati
Hanno in comune la proteina γ‐tubulina nel materiale pericentriolare, critica per l’assemblaggio dei microtubuli
G. Karp: Biologia cellulare e Animale, EdiSES, 1998
50
Centriolo
Struttura cava, cilindrica che si trova nel centrosoma o MTOC della maggior parte delle cellule eucariotiche.
Due centrioli formano una zona centrale a forma di L che è composto di corti microtubuli e da diverse proteine accessorie.
Il più “vecchio” dei due centrioli, chiamato “centriolo madre” inizia l’assemblaggio del più giovane, o “centriolo figlio”, durante la fase di duplicazione dei cromosomi che precede la divisione cellulare.
I centrioli sono coinvolti nell’assemblaggio del fuso mitotico e della citocinesi, fasi in cui aiutano ad organizzare il materiale pericentriolare contenente diversi γ−TuRC.
Tuttavia, i centrioli non nucleano direttamente i microtubuli. Si pensa che l’inizio della formazione dei microtubuli abbia luogo nel materiale pericentriolare e che essi si leghino alle appendici del centriolo “madre”.
Una struttura specializzata simile al centriolo, il corpo basale, è un componente essenziale alla base di tutte le cilia e flagelli.
http://manual.blueprint.org/Home/glossary‐of‐terms/mechano‐glossary‐‐c/glossary‐centriole
Struttura del centrosoma
Nelle cellule che non si stanno dividendo, il MTOC è noto come centrosoma, che consiste in una copia a forma di L di centrioli e di materiale centriolare associato.
http://courses.washington.edu/conj/cell/cell.htm
Il “più vecchio” dei due centrioli ha proteine aggiuntive che formano come delle appendici lungo la superficie esteriore.
Il materiale pericentriolare contiene numerosi γ‐TuRC che nucleano la disposizione di mcirotubuli. Centrioli
I centriloi hanno i MTs organizzati in una struttura simile a quella del corpo basale che si trova alla base di cilia e di flagelli.
http://micro.magnet.fsu.edu/cells/centrioles/centrioles.ht
ml
http://manual.blueprint.org/Home/glossary-of-terms/mechano-glossary--c/glossary-centriole
9
26/05/2013
Iniziazione dell’assemblaggio dei MTs
La nucleazione dei MTs è sfavorevole nelle condizioni normali che si trovano nella maggior parte delle cellule viventi.
Perciò i MTs sono nucleati da un complesso di γ‐tubulina e di altre componenti proteiche note come il “γ‐
tubulin ring complex” (γ‐TuRC). Il MTOC contiene numerosi γ‐TuRC e questa regione della cellula è il sito principale di nucleazione dei MTs.
Il γ‐TuRC nuclea e incapuccia le estremità (‐) e (+) dei nuovi filamenti fornendo siti di legame stabili per i dimeri di tubulina. http://manual.blueprint.org/Home/glossary-of-terms/mechanoglossary--m/mechano-glossary-microtubules/glossarytubulin/glossary-microtubule-initiation
I dimeri di tubulina usano sopratutto interazioni longitudinali per legarsi fra di loro e al γ−TuRC durante la fase di nucleazione.
Man mano la lunghezza del protofilamento aumenta, le interazioni laterali fra i protofilamenti creano una stabilità addizionale che porta ad un MT chiuso.
Microtubuli
ORGANIZZAZIONE DEL CITOPLASMA
(a) Nelle cellule animali in interfase, le estremità (−) della maggior parte dei MT si trova vicino al MTOC. Allo stesso mdo, i MT dei flagelli e delle cilia hanno le loro estremità (−) in continuità con il corpo basale, che funge come il MTOC per queste strutture. (b) Quando le cellule entrano in mitosi, la rete dei MTs si riarrangia, formando un fuso mito co. Le estremità (−) di tutti i MT del fuso puntano verso uno dei due MTOCs, o poli, come vengono chiamati nelle cellule mitotiche. http://www.ncbi.nlm.nih.gov/books/NBK21710/figure/A5469/?report=objectonly
http://www.ncbi.nlm.nih.gov/books/NB
K21580/figure/A5417/?report=objecton
ly
Struttura di neuroni tipici dei mammiferi
(c) Nelle cellule nervose, le estremità (−) dei MT assonali sono orientate verso la base dell’assone. Tuttavia, i MT dei dendriti hanno polarità miste. http://www.ncbi.nlm.nih.gov/books/NB
K21580/figure/A5417/?report=objecton
ly
(d) Nelle cellule vegetali, che contengono numerosi MTOCs, i MTs si allineano lungo la corteccia cellulare. Reti di MTs incapucciano le estremità in crescita di una cellula vegetale. Le frecce indicano la direzione di conduzione dei potenziali di azione negli assoni (rosso). (a) Interneuroni multipolari. Ciascun neurone ha dendriti con ramificazioni profuse, che ricevono segnali nelle sinapsi con diverse centinaia di altri neuroni, e un singolo e lungo assone che si ramifica lateralmente alla sua estremità. (b) Un motoneurone che innerva una cellula muscolare. Tipicamente, i motoneuroni hanno un singolo e lungo assone che si estende dal corpo cellulare fino alla cellula effettrice. Nei, motoneuroni dei mammiferi uno strato isolante di mielina di solito riveste tutte le parti dell’assone all’eccezione dei nodi di Ranvier e terminali assonici. (c) Un neurone sensitivo in cui l’assone si ramifica subito dopo che esce dal corpo cellulare. Il ramo periferico trasporta l’impulso nervoso dalla cellula recettrice fino al corpo cellulare che è localizzato nella radice dorsale del ganglio vicino al midollo spinale; il ramo centrale trasporta l’impulso dal corpo cellulare al midollo spinale a al cervello. Entrambi i rami sono strutturalmente e funzionalmente degli assoni, tranne che nelle pozioni terminali, nonotante il ramo periferico conduca impulsi verso, piuttosto che dal corpo cellulare
60
10
26/05/2013
Microtubuli
Cilia, flagelli, assonema
62
Battito ciliare
Ritmo metacronale del battito delle
cilia
Le cilia sulla superficie di uno protozoo ciliato battono in onde metacronali in cui le
cilia di una data fila sono nello stesso stadio del ciclo di battito, ma quelle delle file
adiacenti sono in stadi diversi. RS: cilia nel “recovery stroke” (battito di recupero);
ES: cilia in battito di potenza effettivo.
http://kentsimmons.uwinnipeg.ca/cm1504/cytoskeleton.htm
65
11
26/05/2013
Corpuscoli basali e altri MTOCs ‐1
I microtubuli che formano l’assonemas di un ciglio o di un flagello prendono origine da una struttura detta corpo basale che si trova alla base dell’organello. I corpi basali hanno la stessa struttura di un centriolo e infatti corpi basali e centrioli danno orgini gli uni agli altri. Il flagello di uno spermatozoo, ad es., si forma da un corpo basale derivato da un centriolo che faceva parte del fuso mitotico dello spermatocita da cui lo spermatozoo ha preso origine.
67
68
Movimenti di cilia e flagelli
69
http://www.ncbi.nlm.nih.gov/books/NBK9833/figure/A1846/
70
Struttura di cilia e flagelli
71
72
12
26/05/2013
Dineina diffettosa
‐ Notare la perdita di organizzazione strutturale
http://kentsimmons.uwinnipeg.ca/cm1504/cytoskeleton.htm
Corpuscoli basali e altri MTOCs ‐2
Altri tipi di cellule hanno tipi differenti di MTOC. Nei funghi, l’MTOC principale appare come una struttura discoidale, detta corpo polare del fuso ed é inserita nell’involucro nucleare. I microtubuli che si estendono dal corpo polare del fuso verso l’esterno, formano parte del citoscheletro citoplasmatico, mentre i microtubuli che crescono verso l’interno fanno parte del fuso mitotico (che in questi organismi si forma all’interno del nucleo).
Sindrome di Kartagener
Incapacità di sintetizzare microtubuli
Cilia e flagelli sono immobili
Infezioni respiratorie croniche
Sterilità maschile
Le piante sono prive sia di centrosomi che di centrioli; il principale MTOC di una cellule vegetale é costituito da porzioni di materiale situato sulla superficie esterna dell’involucro nucleare da cui emergono i microtubuli del citoscheletro.
75
UniVe
La struttura dei flagelli differerisce fra (la maggior parte dei) procarioti e degli eucarioti
Microtubuli
Fuso mitotico
78
13
26/05/2013
Fuso mitotico
http://www.ncbi.nlm.nih.gov/bo
oks/NBK26934/figure/A3350/
http://www.microscopyu.com/staticgallery/featuredmicroscopist/deerinck/deerinckimage13.html
80
Fuso mitotico
Formazione del fuso mitotico
http://www.ncbi.nlm.nih.gov/books/N
BK9932/figure/A1825/
81
http://www.ncbi.nlm.nih.gov/books/NBK9932/figure/A1830/
I centrioli e i centrosomi si duplicano durante l’interfase.
Durante la profase della mitosi, i centrosomi duplicati si separano e si muovono verso le estremità opposte del nucleo. L’involucro nucleare si disaggrega, e i microtubuli si riorganizzano per formare il fuso mitotico. I microtubuli del cinetocore si legano ai cromosomi condensati, mentre i microtubuli polari si sovrappongono al centro della cellula, e i microtubuli astrali si estendono verso la periferia della cellula.
Nella metafase i cromosomi condensati si allineano al centro del fuso. Fuso mitotico - 1
Microtubuli
Farmaci
http://ehumanbiofield.wikispaces.com/Chromosomes+HW4+MC
84
14
26/05/2013
Colchicina e tassolo: Farmaci specifici per i microtubuli
Questi e altri farmaci che interferiscono con l’assemblaggio e il disassemblaggio normale dei microtubuli hanno un effetto
antimitotico che è particolarmente devastante nelle cellule in rapida divisione, come le cellule tumorali (e cellule normali quali quelle del midollo osseo, dell’intestino e o dell’epidermide). Molecular Cell Biology 19. Cell Motility and Shape II: Microtubules and Intermediate Filaments 19.2. Microtubule Dynamics
85
and Associated Proteins
86
http://www.ncbi.nlm.nih.gov/books/NBK26862/table/A2992/
87
Cultured human epithelial cells form primary cilia (green:
cilia; red: basal body).
Microtubuli
SEMINARIO: Cilia primarie o immobili
http://www.kidneyresearchcenter.org/index.php/fuseaction/scaleup/scaleupID/162/Cilia.htm
http://deepspace.jpl.nasa.gov/dsn/images/picture_70_bw_lg.jpg
15
26/05/2013
Cilia primarie (immobili)
Le cilia primarie/immobili si trovano in praticamente tutte le cellule del corpo.
Rispetto alle cilia mobili, di solito è presente un solo un cilio
primario per cellula.
Negli organi sensoriali (es. occhio, naso) sono presenti cilia primarie specializzate:
Il segmento esterno della cellule fotorecettrici bastoncelli dell’occhio è collegato al corpo cellulare da un cilio specializzato immobile.
Anche Il “dendritic knob” dei neurono olfattivi, in cui si trovano i recettori dell’odorato, contiene cilia immobili (circa 10 cilia per “dendritic knob”).
http://en.wikipedia.org/wiki/Cilium
Cilia primaria: cosa le rende diverse dalle cilia mobili?
Mancano della paia centrale di microtubuli, il che spiegherebbe la mancanza di mobilità.
In più sembrano anche essere privi di dineina, uno dei motori molecolari necessari per la motilià.
Inoltre, alcune cilia primarie non si proiettano oltre la superficie cellulare e la maggior parte, se non tutte, sono molto corte.
Che funzione svolgono allora questi organelli se non protrudono fuori dalla cellule nè sono mobili?
Molti scienziati hanno ritenuto che potessero essere semplicemente organi vestigiali senza alcun ruolo reale, mentre altri hanno notato che le cilia primaria sembrano essere presenti in zone chiave che coinvolgono la visione, l’udito, e altre forme di input sensoriali.
Negli anni1990 gli scienziati cominciarono a guardare alle cilia primarie con interesse sempre crescente ma la sorgente dell’interesse è venuta da un candiato alquanto improbabile.
Cilia primaria ‐ 1
Per molto tempo quasi tutta l’attenzione è stata data alle cilia mobile perchè la loro funzione era immediatamente osservabile.
Ma gli scienziati, già dal 1868 con Kowalevsky, hanno riportato la presenza di cilia singole (immobili) in una gran diversità di cellule dei vertebrati.
Queste cilia solitarie e apparentemente non funzionali sono in realtà molto più diffuse del tipo mobile.
Ad esempio, nell’Uomo solo alcuni tipi cellulari hanno cilia mobili, in particolare gli spermatozoo, nelle cellule epiteliali dei bronchi e degli ovidotti, e le cellulem ependimali che rivestono le vescicole cerebrali.
Viceversa, praticamente tutti gli altri tipi cellulari hanno un cilio primario.
http://www.nature.com/scitable/topicpage/the‐primary‐cilium‐an‐orphan‐organelle‐finds‐14228249
Chlamydomonas e il trasporto intraflagellare
Chlamidomonas: Modello classico per studiare la struttura dei flagelli.
Sistema di trasporto
intraflagellare (IFT): sistema
di trasporto intracellulare
che costruisce e mantiene
cilia e flagelli.
Una delle proteine coinvolte
nella funzione di IFT è la stessa coinvolta nella
malattia del rene policistico
(Polycystic kidney disease, PKD) nei topi.
Le cellule che rivestono il nefrone hanno
cilia primarie.
I topi con PKD non sono in grado di assemblare le cilia come si deve. La PKD è la più importante patologia
renale nell’Uomo.
http://www.nature.com/scitable/topicpage/the‐primary‐cilium‐an‐orphan‐organelle‐finds‐14228249
Le cilia come recettori sensoriali ‐ 1
Le cilia primarie nelle cellule normali rispondono al segnale meccanico
prodotto da un fluido che fluisce per iniziare la rapida captazione di calcio.
Le cellule PKD, che mancano di cilia primarie, non mostrano alcuna
risposta al flusso. I complessi di Bardet‐Biedl contengono una gran
varietà di proteine tipo rivestimento che si possono
legare e traportare diverse proteine recettrici verso il
cilio.
Quando una proteina di indirizzamento al cilio si collega, l’intero complesso si
muoverà verso la membrana del cilio primario e permetterà al recettore di inserirsi nella membrana. http://www.nature.com/scitable/topicpage/the‐primary‐cilium‐an‐orphan‐organelle‐finds‐14228249
16
26/05/2013
Christensen et al., 2007
Christensen et al., 2007
Christensen et al., 2007
Christensen et al., 2008
17
26/05/2013
Christensen et al., 2008
Christensen et al., 2008
18
26/05/2013
19
26/05/2013
Singla & Reiter, 2006
http://www.sciencemag.org/content/313/5787/629/F1.large.jpg
Singla & Reiter, 2006
• Primary cilia are highly structured and are found in many organisms and on many cell types. (A) Electron micrograph of the primary cilium of a canary brain radial glia (69). (B) Schematic showing structure of the basal body and primary cilium [modified from (6, 70)]. (C) The green alga Chlamydomonas showing flagella (green, arrow) and basal body (red). Nuclei are blue. [(D) to (L)] Scanning electron and immunohistological images of primary cilia (arrows) of (D) the mouse node, (E) the mouse neural tube, emanating from basal bodies (red), (F) the Xenopus neural tube, (G) the zebrafish
neural tube, (H) a mouse neurogenic astrocyte, (I) a mouse embryonic epidermal cell, (J) a mouse somite, (K) mouse embryonic stem cells, and (L) mouse astrocytes expressing glial fibrillary acidic protein (red). Also shown in (H) are motile ependymal cell cilia (arrowhead). Scale bars, 1 μm [(A), (C), and (D)] and 10 μm [(E) to (L)]. http://www.sciencemag.org/content/313/5787/629/F1.expansion.html
20








![[ XI ] INDICE GENERALE Riconoscimento e interazioni tra cellule](http://s1.studylibit.com/store/data/001175494_1-9062ea75af318561914dbe68aa0e27ca-300x300.png)