Anteprima Estratta dall' Appunto di
Biochimica
Università : Università degli studi di Padova
Facoltà : Medicina
Indice di questo documento
L' Appunto
Le Domande d'esame
ABCtribe.com e' un sito di knowledge sharing per facilitare lo scambio di materiali ed informazioni per lo studio e la
formazione.Centinaia di migliaia di studenti usano ABCtribe quotidianamente per scambiare materiali, consigli e
opportunità Più gli utenti ne diffondono l'utilizzo maggiore e' il vantaggio che ne si può trarre :
1. Migliora i tuoi voti ed il tempo di studio gestendo tutti i materiali e le risorse condivise
2. Costruisci un network che ti aiuti nei tuoi studi e nella tua professione
3. Ottimizza con il tuo libretto elettronico il percorso di studi facendo in anticipo le scelte migliori per ogni esame
4. Possibilità di guadagno vendendo appunti, tesine, lezioni private, tesi, svolgendo lavori stagionali o part time.
www.ABCtribe.com
ABCtribe.com - [Pagina 1]
L' Appunto
A partire dalla pagina successiva potete trovare l' Anteprima Estratta dall' Appunto.
Se desideri l'appunto completo clicca questo link.
Il numero di pagina tra le parentesi tonde si riferisce a tutto il documento contenente l'appunto.
Sull'appunto stesso in alcuni casi potrete trovare una altra numerazione delle pagine che si riferisce al solo
appunto.
Grafico dei voti medi per questo
esame:
Grafico dei giorni medi per la
preparazione di questo esame:
Grafico Copyright © ABCtribe.com. Vietata la
riproduzione.
Grafico Copyright © ABCtribe.com. Vietata la
riproduzione.
Clicca qui per i grafici aggionati su Biochimica >>
ABCtribe.com - [Pagina 2]
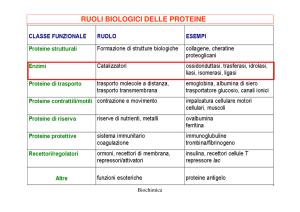
GLI ENZIMI
1. Definizione.
Un enzima è un catalizzatore (acceleratore) di reazioni biologiche. Un catalizzatore è una sostanza
che è in grado di accelerare lo svolgimento di una reazione chimica diminuendo l'energia richiesta
per farla iniziare (energia di attivazione) aumentando, in ultima analisi, la velocità della reazione in
oggetto. Il catalizzatore partecipa alla reazione, ma non venendo “consumato” ne esce inalterato e
quindi può essere riutilizzato. Nel caso di reazioni all’equilibrio, l’enzima accelera il
raggiungimento dell’equilibrio* di una reazione chimica (spontanea!) senza spostarlo. Quindi
accelera (da 107 a 1014 volte!) della stessa entità sia la reazione in un senso che quella in senso
opposto:
A+B
C+D
om
Quasi tutti gli enzimi sono proteine (alcuni RNA hanno attività catalitica).
e.c
*L’equilibrio (dinamico) di reazione è raggiunto quando le concentrazioni relative di reagenti e
prodotti non cambiano più nel tempo.
AB
Ct
rib
2. Meccanismo d’azione.
Quando un substrato S viene convertito in prodotto P (a minore contenuto energetico rispetto a S,
altrimenti la reazione sarebbe termodinamicamente impossibile e nessun enzima sarebbe in grado di
farla avvenire!) le molecole di S passano attraverso uno stato intermedio tra S e P (lo stato di
transizione , S*) caratterizzato da un contenuto energetico maggiore di S. La differenza tra il
contenuto energetico di S e quello di S* è detta energia di attivazione di S. L’enzima è in grado di
legare S* (ha maggiore affinità per S* che per S) e il legame E-S* abbassa l’energia di
attivazione : la formazione di legami tra E ed S* libera energia (energia di legame E-S*) e abbassa
il contenuto energetico di E-S*. Quindi statisticamente, nell’unità di tempo, più molecole di S
riusciranno a superare la barriera di attivazione e procedere lungo la direzione di reazione.
L’enzima lega S* a livello di un sito preciso sulla superficie della proteina che si chiama sito attivo:
questa tasca, di solito superficiale, corrisponde esattamente per forma e disposizione di cariche
elettriche e/o residui idrofobici (dei gruppi R degli amminoacidi che foderano il sito attivo) a S*.
Si può immaginare che S* sia la chiave e il sito attivo sia la serratura specifica per quella chiave.
I legami che si formano tra S* e il sito attivo sono di solito legami deboli (ponti H, legami ionici e
idrofobici) anche se in alcuni enzimi si tratta di legami covalenti.
ABCtribe.com - [Pagina 3]
L’esistenza di un sito attivo dove si lega il substrato e dove avviene la sua trasformazione in
prodotto determina alcune proprietà comuni a tutti gli enzimi:
specificità, saturabilità e inibibilità.
Specificità: ogni enzima riconosce specificamente il/i substrato/i e non altre molecole, anche
chimicamente simili a S.
Saturabilità: quando tutti i siti attivi di tutte le molecole enzimatiche presenti sono legate al
substrato (saturate) la velocità di catalisi è massima e non può aumentare ulteriormente, anche
aggiungendo altro substrato.
Inibibilità: una molecola che assomigli moltissimo al substrato (per forma, distribuzione di cariche,
ecc.) e che si leghi al sito attivo può spiazzare il vero substrato dal legame con il sito attivo e inibire
l’attività enzimatica (inibizione competitiva). L’aggiunta di un eccesso di substrato, però, rende
reversibile l’inibizione. Molti farmaci agiscono per inibizione enzimatica.
om
3. Cinetica enzimatica.
L’attività catalitica degli enzimi è caratterizzata da due parametri cinetici che consentono di
confrontare l’azione di enzimi diversi e la loro affinità per il substrato. Questi due parametri sono la
costante di Michaelis-Menten (Km) e la velocità massima (Vmax). Con l’aumentare della
concentrazione di S ([S]) la velocità di catalisi (prodotto/tempo) aumenta secondo una funzione
iperbolica fino a un valore massimo (Vmax), raggiunto quando tutti i siti attivi disponibili sono
saturati da S.
AB
Ct
rib
e.c
Cinetica enzimatica degli enzimi a singola subunità
Da questo grafico si può derivare il valore di Km, cioè la [S] alla quale V = ½ Vmax.
Dato che né il valore di Vmax né quello di Km possono essere calcolati in modo preciso da una
curva iperbolica, si preferisce trasformare questa curva in una retta (prendendo il reciproco di ogni
valore), dall’equazione della quale si possono calcolare precisamente -1/Km e 1/Vmax.
Vmax = velocità massima di catalisi, in condizioni di saturazione dell’enzima (dipende dalla [E])
Km = concentrazione di S a cui la velocità di catalisi è metà della massima (non dipende dalla [E]).
Km è una misura dell’affinità dell’enzima per S: maggiore la Km (che si misura in molarità),
minore l’affinità.
Un inibitore competitivo (si lega al sito attivo al posto di S) aumenta la Km e lascia invariata la
Vmax (un eccesso di S spiazza l’inibitore e l’enzima raggiunge comunque la sua Vmax, anche se ad
una [S] maggiore rispetto alla condizione senza inibitore).
ABCtribe.com - [Pagina 4]
Un inibitore non competitivo (si lega all’enzima in un punto diverso dal sito attivo, ma induce una
modifica conformazionale della proteina enzimatica tale per cui S non si può più legare al sito
attivo) abbassa Vmax ma lascia invariata Km (affinità dell’enzima per S).
Enzimi allosterici.
Allosterico significa “altro sito” (oltre al sito attivo): gli enzimi allosterici, oltre al sito attivo dove si
lega il substrato, possiedono un altro sito dove si lega il regolatore (attivatore o inibitore). Il legame
del regolatore modifica la conformazione del sito attivo e quindi modifica l’attività catalitica
dell’enzima. Quasi tutti gli enzimi allosterici sono formati da più di una subunità proteica: in questi
casi può verificarsi un effetto cooperativo tra i siti attivi presenti sulle diverse subunità.
La cooperatività di legame con il substrato si ha quando una subunità proteica dell’enzima lega S e
induce una modifica conformazionale nelle altre subunità che ne aumenta l’affinità per S. La
conseguenza è che piccole variazioni della [S] inducono grandi variazioni della velocità di catalisi.
La cinetica enzimatica è descritta allora da una curva sigmoide anziché iperbolica.
AB
Ct
rib
e.c
om
Cinetica enzimatica degli enzimi allosterici che mostrano cooperatività di legame E-S
La cooperatività è un meccanismo per regolare l’attività enzimatica sulla base della disponibilità di
S, ma gli enzimi hanno anche altri meccanismi di regolazione, come descritto sotto.
4. Classi di enzimi.
Gli enzimi vengono suddivisi in 6 classi sulla base del tipo di reazione che catalizzano.
Classe 1: ossido-reduttasi o deidrogenasi. Catalizzano reazioni di ossido-riduzione, con l’aiuto di
specifici coenzimi vitaminici (NAD+, NADP + o FAD) che fungono da trasportatori di elettroni.
Classe 2: transferasi. Catalizzano reazioni di trasferimento di gruppo. Ad esempio, le cinasi
trasferiscono un fosfato dall’ATP ad una molecola accettrice (o viceversa):
X + ATP ↔ XP + ADP ;
possono essere reazioni reversibili o irreversibili a seconda del ∆G di idrolisi del legame X-P
confrontato con il legame ADP-P (il “potenziale di trasferimento del fosfato”).
Classe 3: idrolasi. Catalizzano la rottura di un legame per introduzione di una molecola d’acqua:
XY + H2 O → X-H + Y-OH ; sono sempre reazioni irreversibili.
Classe 4: liasi. Catalizzano reazioni di addizione a doppi legami, del tipo:
X + Y ↔ XY (NON richiede energia!) ; sono reazioni reversibili.
Classe 5: isomerasi. Catalizzano reazioni (reversibili) di isomerizzazione.
Classe 6: ligasi o sintetasi. Catalizzano la formazione di nuovi legami chimici con dispendio di
energia (ATP): X + Y + ATP → XY + ADP + Pi ; sono sempre reazioni irreversibili.
ABCtribe.com - [Pagina 5]
Gli enzimi più importanti per lo studio del metabolismo energetico sono le ossidoreduttasi e le
cinasi. Attraverso reazioni di ossidazione dei substrati metabolici (glucoso, fruttoso, acidi grassi) si
formano i coenzimi ridotti (NADH e FADH2 ) che, nella catena respiratoria mitocondriale, saranno
utilizzati per generare il gradiente protonico necessario alla sintesi dell’ATP.
Le cinasi catalizzano le reazioni di trasferimento del fosfato dall’ATP su glucoso e fruttoso (che
servono ad attivare questi zuccheri e ne consentono l’utilizzazione metabolica) e catalizzano
reazioni di trasferimento del fosfato da molecole fosforilate sull’ADP, con formazione di ATP,
nelle cosiddette “reazioni di fosforilazione a livello del substrato” nella glicolisi.
om
5. Meccanismi catalitici.
Gli enzimi agiscono attraverso uno o più dei seguenti meccanismi:
• distorsione del substrato: per legarsi al sito attivo il substrato deve essere leggermente distorto
(S*) e questo favorisce la successiva trasformazione in prodotto;
• riduzione entropica: diminuzione dei moti molecolari relativi dei due substrati che devono
reagire, perché legati entrambi nel sito attivo;
• desolvatazione: aumento della forza delle interazioni deboli (legami ionici, idrofobici, ponti H)
tra 2 substrati per allontanamento delle molecole d’acqua intorno a loro;
• riduzione della diffusione dei prodotti intermedi: nei complessi multienzimatici, i prodotti
intermedi non abbandonano il complesso proteico (aumenta la velocità e l’efficienza catalitica).
e.c
Molti enzimi richiedono cofattori per funzionare: ad es. ioni metallici o coenzimi. Se legati
covalentemente i cofattori sono detti gruppi prostetici.
AB
Ct
rib
Tutti i coenzimi (molecole non proteiche necessarie alla funzione dell’enzima) contengono nella loro molecola
una parte vitaminica: il nostro organismo non è in grado di sintetizzare le vitamine, che devono essere assunte
con l’alimentazione. Una volta assorbite, le nostre cellule sono in grado di trasformarle nei corrispondenti
coenzimi. Data la fondamentale importanza degli enzimi nel consentire il metabolismo si capisce perché la
carenza di vitamine sia una condizione incompatibile con la vita.
Coenzimi vitaminici
VITAMINA
COENZIMA
Tiamina (B1)
Acido pantotenico (B5)
Piridossina (B6)
Biotina (H)
Acido Folico (B9)
TPP
Coenzima A (CoA)
Piridossalfosfato (PLP)
Biocitina
Acido tetraidrofolico (THF)
Acido nicotinico (B3)
Nicotinamide adenina dinucleotide
(NAD)
Flavin adenina dinucleotide
Riboflavina (B2)
ABCtribe.com - [Pagina 6]
ENZIMA/FUNZIONE
Piruvato deidrogenasi
Trasportatore di acili
Transaminasi
Carbossilasi
Trasferimento unità
monocarboniose
Deidrogenasi (trasferimento di
elettroni)
Deidrogenasi (trasferimento di
elettroni)
Questo documento e' un frammento dell'intero appunto utile come
anteprima. Se desideri l'appunto completo clicca questo link.
ABCtribe.com - [Pagina 7]
Preparati con le domande di ABCtribe su Biochimica.
1. calcolare il numero di molecole di atp prodotte durante la completa ossidazione a co2 e acqua
del triacilglicerolo avente acido steaerico in po
Risposta:
devi sapere quanti atomi di carbonio ci sono nell acido stearico (è un acido saturo o insaturo?)
saputo questo sai quanti cicli servono per ossidarlo tutto,
dalla teoria sai quanti ATP si prosucono per un ciclo ossidativo, moltiplichi per 2 perchè di acido stearico
ce ne sono 2, poi fai lo stesso ragionamento con l'acido oleico e sommi il tutto.
al numero ottenuto sommi che il glicerolo entra in glicolisi facendo questa reazione
glicerolo--------->gliceraldeide e riduzione di NAD+--->NADH + H+
gliceraldeide----->gliceraldeide3 fosfato e spesa
[Clicca qui >> per continuare a leggere].
2. se devo ossidare una molecola di succinilcoa a co2 e h2o entra nel cicl
Risposta:
2 scetil CoA + 2ADP + 6NAD+ + 2FAD --> 4CO2 + 2ATP + 6NADH + 2FADH2
ne
[Clicca qui >> per continuare a leggere].
* Carica Appunti,Domande,Suggerimenti su : Biochimica e guadagna punti >>
* Lezioni Private per Biochimica >>
Avvertenze:
La maggior parte del materiale di ABCtribe.com è offerto/prodotto direttamente dagli studenti (appunti, riassunti,
dispense, esercitazioni, domande ecc.) ed è quindi da intendersi ad integrazione dei tradizionali metodi di studio e non
vuole sostituire o prevaricare le indicazioni istituzionali fornite dai docenti. Il presente file può essere utilizzato in base
alle tue necessità ma non deve essere modificato in alcuna sua parte, conservando in particolare tutti i riferimenti
all’autore ed a ABCtribe.com; non potrà essere in alcun modo pubblicato tramite alcun mezzo, senza diverso accordo
scritto con l’autore ed i responsabili del progetto ABCtribe.com. Per eventuali informazioni, dubbi o controversie
contattate la redazione all’indirizzo [email protected].
ABCtribe.com - [Pagina 8]