La storia di un uomo nei nove
mesi che precedono la sua
nascita è probabilmente più
interessante e comprende eventi
molto più importanti di tutti
quelli dei settant’anni che
seguono.
S. Taylor Coleridge 1885
Lo scopo finale di tutte le
relazioni amorose, siano esse
comiche o tragiche, è in realtà
più importante di tutti gli altri
scopi della vita umana. Quello a
cui si riferisce è niente di meno
che la composizione della
generazione successiva.
A. Schopenhauer (citato da C. Darwin
1871)
Il Ciclo Cellulare
Nel corso della loro vita tutte le cellule subiscono
una serie di cambiamenti molecolari e morfologici
definiti complessivamente ciclo cellulare.
Esso indica il periodo che intercorre tra l’origine di
una cellula da una precedente divisione ed il momento
in cui questa si divide nuovamente.
Il ciclo cellulare comprende quattro fasi sequenziali
denominate G1, S, G2 e M, oltre ad una fase G0. Nel
loro insieme le fasi G1, S, G2 costituiscono
l’interfase, mentre la fase M indica il processo di
divisione cellulare per mitosi o per meiosi. Le fasi G1
e G2 (G=Gap=intervallo) sono considerate fasi di
riposo in quanto in esse non c’è duplicazione del DNA.
Tuttavia, durante queste fasi, la cellula è
metabolicamente attiva e compie le sue funzioni
specializzate preparatorie alle successive fasi del
ciclo. Durante la fase S (S=sintesi) si attuano la
duplicazione del DNA e la sintesi di istoni, che
portano alla replicazione cromosomica.
L’intero ciclo cellulare si svolge in un periodo compreso tra alcune ore e
qualche giorno, in rapporto alla fase G1 la cui durata varia a seconda della linea
cellulare considerata.
Le cellule che vanno incontro ad un profondo differenziamento o che non si
dividono ulteriormente (es. neuroni e cellule muscolari cardiache) prolungano
indefinitamente la fase G che pertanto viene detta G0. Altri tipi cellulari,
come le cellule epiteliali e gli epatociti, possono rientrare nel ciclo cellulare
dalla fase G0 e procedere con la divisione mitotica in seguito ad appropriati
stimoli quali: fattori di crescita, sostanze mitogeniche, segnali da altre cellule
e dalla matrice extracellulare.
Le cellule della linea germinale
(spermatogoni ed ovogoni) dopo
una serie di cicli cellulari in cui si
dividono per mitosi, vanno
incontro ad un ciclo che termina
con una divisione meiotica.
Le cellule somatiche e le cellule germinali (spermatogoni ed ovogoni) di
ogni specie hanno un corredo cromosomico completo per cui sono
denominate diploidi ed indicate con 2n.
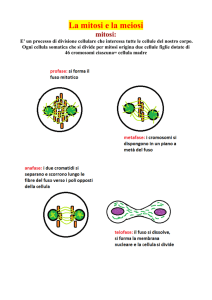
Mitosi
Il termine mitosi è usato per descrivere la divisione equazionale di una
cellula con formazione di due nuove cellule con lo stesso corredo
cromosomico della cellula progenitrice da cui sono derivate. La mitosi è
essenziale per lo sviluppo embrionale e per riparare e rimpiazzare i
tessuti durante la vita. La mitosi può essere divisa in quattro fasi:
profase, metafase, anafase e telofase.
Durante la profase (A, B), i cromosomi, formati dai cromatidi fratelli
spiralizzano ed i nucleoli si disgregano. Nel citoplasma 2 coppie di
centrioli duplicati durante l’interfase, cominciano a formare il fuso
mitotico distinto in un fuso mantellare ed uno cromosomiale, responsabile
del successivo movimento dei cromatidi verso i poli opposti della cellula in
divisione. Segue una breve pro-metafase (C), caratterizzata dalla
disgregazione dell’involucro nucleare, e quindi la metafase propriamente
detta (D), caratterizzata dall’organizzazione dei cromosomi all’equatore
della cellula. Il cinetocore, un complesso proteico che si forma sui
centromeri alla fine della profase, agisce come una piattaforma di
attacco per i microtubuli del fuso. Si forma quindi un complesso
microtubulo-cinetocore che consente il movimento dei cromosomi che si
posizionano a metà strada tra i due poli della cellula, in una regione
denominata piastra metafasica o equatoriale. Ogni cromatide fratello è
attaccato al centrosoma mediante il proprio complesso microtubulocinetocore. Nell’anafase (E) si distinguono due stadi: anafase A in cui le
coppie di cromatidi fratelli si separano sincronicamente per la scissione
dei centromeri e per l’accorciamento dei complessi microtubulocinetocore. I due set di cromatidi appena separati vengono trascinati
verso i due poli opposti della cellula. Nell’anafase B lo spostamento dei
due cromatidi verso i poli opposti è accompagnato dall’allontanamento dei
poli. Nella telofase (F), i due gruppi di cromosomi identici, raggruppati ai
rispettivi poli, decondensano; inoltre, quelli con costrizione secondaria
ripristinano il nucleolo, attorno a ciascun set di cromosomi si ricostituisce
un involucro nucleare e termina la cariocinesi. Dopo la formazione dei
nuovi involucri nucleari, un anello contrattile di actina stringe la
membrana cellulare e divide il citoplasma in modo da separare
completamente le due cellule figlie. Quest’ultimo processo definito
citocinesi o citodieresi, normalmente porta alla formazione di due cellule
figlie di uguali dimensioni; occasionalmente differenti quantità di
citoplasma ed organuli possono essere distribuiti tra le due cellule figlie.
A
C
E
B
D
F
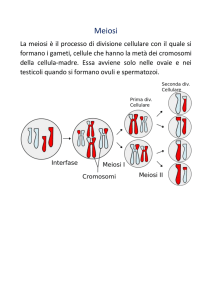
Meiosi
E’ il processo di divisione riduzionale, esclusivo delle cellule della linea
germinale e rappresenta l’ultimo evento di divisione della
gametogenesi.
La meiosi porta alla formazione di gameti aploidi a partire da cellule
diploidi in quanto consiste di una sola fase S seguita da due divisioni
consecutive. In conseguenza, dalla cellula di partenza diploide, si
formano, in due tappe, quattro cellule aploidi. Le due divisioni,
separate da una breve intercinesi, sono indicate come I e II
divisione meiotica, ognuna delle quali si può suddividere in fasi
corrispondenti a quelle della mitosi: profase, metafase, anafase e
telofase.
Tuttavia nella meiosi queste fasi presentano caratteristiche peculiari
anche se la cellula, che entra in profase I, presenta la stessa quantità
di DNA di una cellula che si avvia alla mitosi.
La profase della I divisione meiotica (A-D), durante la quale
avvengono molti eventi intracellulari cruciali, è molto lunga e può
essere ulteriormente divisa in cinque sottostadi: leptotene, zigotene,
pachitene, diplotene e diacinesi.
Questi sottostadi prendono il nome dagli eventi salienti che si
verificano nel corso della profase:
• leptotene, inizio della condensazione della cromatina che continua
per tutta la profase fino a formare cromosomi spessi e corti (A);
• zigotene, appaiamento dei cromosomi omologhi duplicati per formare
le tetradi, ad opera di un complesso di natura proteica detto
complesso sinaptinemale che persiste fino allo stadio successivo (B);
•pachitene, scambio di materiale genetico, per un processo di
frattura e successiva saldatura tra i cromatidi non fratelli dei due
cromosomi omologhi, definito crossing-over (C);
• diplotene, separazione dei cromosomi omologhi che restano ancora in
contatto nei punti in cui si è verificato il crossing-over (chiasmi) (D);
• diacinesi, condensazione completa dei bivalenti con separazione degli
stessi a livello dei chiasmi (terminalizzazione dei chiasmi).
E’ da notare che solo nella spermatogenesi, il processo di
citocinesi, successivo sia alla I che alla II telofase meiotica,
divide equamente il citoplasma tra le cellule. Nell’ovogenesi, al
contrario, dopo la I telofase una delle due cellule risultanti
trattiene la maggior parte del citoplasma divenendo ovocita
II, la più piccola delle due cellule è denominata I globulo
polare. Una distribuzione ineguale di citoplasma viene
mantenuta anche tra l’ovocita secondario ed il II globulo
polare, mentre nel caso in cui anche il I globulo polare vada
incontro alla seconda divisione meiotica, lo scarso citoplasma a
disposizione viene equamente ripartito.
In metafase I, le coppie di cromosomi omologhi si attaccano mediante i
loro cinetocori ai microtubuli del fuso e si posizionano in piastra
equatoriale. I centromeri dei due omologhi sono orientati verso i poli
opposti del fuso, mentre i due cromatidi di ciascuno omologo sono ancora
uniti a livello del centromero. Durante l’anafase I (E), le diadi, costituite
ognuna dai due cromatidi di ognuno degli omologhi, si spostano ai poli
opposti della cellula. A differenza di quanto avviene in anafase mitotica,
ai poli opposti della cellula migrano singoli cromosomi della coppia di
omologhi, ma ancora costituiti da due cromatidi che possono avere
effettuato il crossing-over. A questo punto la distribuzione dei
cromosomi omologhi di derivazione paterna e materna è casuale, ed è alla
base del principio mendeliano dell’assortimento casuale. In telofase I,
intorno a ciascun set di cromosomi separati si forma l’involucro nucleare,
mentre ha luogo la citodieresi. Le due cellule figlie derivate dalla prima
divisione meiotica contengono un numero aploide di cromosomi (un
rappresentante di ogni coppia di omologhi) con una quantità 2c di DNA
dal momento che ogni cromosoma è ancora costituito da due cromatidi.
Dopo una breve fase di riposo, chiamata intercinesi o interfase (F),
durante la quale si ha la formazione di un nuovo fuso, ciascuna cellula
figlia inizia la seconda divisione meiotica. La profase II è più breve
della profase I; il nucleo contiene un set di diadi non interamente
despiralizzate, composte ognuna da una coppia di cromatidi fratelli
connessi mediante un unico centromero. La metafase II è simile alla
metafase I poiché i cromosomi sono posizionati in piastra equatoriale dal
complesso microtubulo-cinetocore. In questo caso il centromero si è
sdoppiato in due cinetocori ognuno su ciascuno dei singoli cromatidi.
Questo consente ai microtubuli di attaccarsi a ciascun cromatide.
Durante l’anafase II (G) i singoli cromatidi legati al complesso
microtubulo-cinetocore migrano verso i poli opposti della cellula in
divisione. Alla fine della telofase II si riformano gli involucri nucleari
attorno a ciascun set di cromatidi ed il citoplasma si divide nuovamente.
Pertanto alla fine della citodieresi le cellule terminali della meiosi
posseggono n cromosomi ed una quantità c di DNA e sono quindi aploidi,
ma il loro corredo genico è rinnovato dal crossing-over e dalla
ripartizione casuale dei cromosomi (H).
GAMETOGENESI
I gameti sono cellule aploidi altamente differenziate derivanti, nel corso dello sviluppo embrionale, da cellule
germinali primordiali. Esse sono caratterizzate da un costituente citoplasmatico, il plasma germinale,
preesistente nell’uovo prima della fecondazione.
Nel corso della segmentazione (della gastrulazione nei Mammiferi), dallo zigote si formano numerose cellule
somatiche ed un limitato numero di cellule germinali primordiali in cui viene segregato il plasma germinale.
In sede extraembrionale (nel topo si rendono visibili nell’endoderma del sacco vitellino ad una settimana
dalla fecondazione, mentre nell’uomo alla terza settimana) si assiste alla proliferazione di tali cellule che
attraverso il circolo sanguigno, migrano attraverso il mesentere dorsale nei territori gonadici in sviluppo. Le
cellule germinali primordiali o protogoni sono mitoticamente attive fino alla nascita nella femmina e per tutta
la vita nel maschio. Le cellule germinali, definite spermatogoni nelle gonadi maschili e ovogoni nelle gonadi
femminili, vanno incontro ad una sequenza di sviluppo simile, che viene indicata come processo di
gametogenesi.
Durante
la
gametogenesi
avvengono
sostanzialmente 3 tipi di eventi anche se con
alcune differenze quantitative, qualitative e
cronologiche tra i due sessi:
• proliferazione mitotica dei goni, (fino alla
nascita nella femmina in cui tutti gli ovogoni
entrano in profase I; per tutta la vita nel maschio
in cui gli spermatogoni dalla maturità sessuale in
poi, possono dividersi sia per mitosi che entrare
in meiosi divenendo spermatociti primari);
• divisione meiotica, (discontinua e completa
solo in seguito a fecondazione nella femmina;
continua e completa nel maschio);
• trasformazioni morfo-fisiologiche dei futuri
gameti, (auxocitosi nella femmina durante la
profase I; spermiogenesi nel maschio dopo la
meiosi).
Ovogenesi
Spermatogenesi
Mitosi e Meiosi iniziano una volta
per l’intera popolazione cellulare di
ovogoni
Mitosi
e
continuamente
popolazione
spermatogoni
Meiosi
iniziano
per parte della
cellulare
di
La meiosi si arresta e riprende in
una popolazione cellulare più
esigua
La meiosi procede in modo continuo
Ogni meiosi produce solo 1 gamete
e solo in seguito a fecondazione,
che può avvenire mesi o anni più
tardi rispetto all’inizio della meiosi
Ogni meiosi produce 4 gameti e la
meiosi si completa in giorni o
settimane
Il differenziamento del futuro
gamete avviene nella profase I,
quando l’ovocita è diploide
Il differenziamento del gamete
avviene dopo la meiosi, a carico
dello spermatidi aploide
Tutti i cromosomi subiscono una
ricombinazione durante la profase I
I cromosomi sessuali sono esclusi
dalla ricombinazione durante la
profase I
Nella I e nella II citocinesi
una
delle due cellule risultanti trattiene
la maggior parte del citoplasma
Nella I e nella II citocinesi
il
citoplasma è equamente diviso tra le
cellule
Tutti gli eventi legati alla gametogenesi sono finalizzati all’acquisizione da parte dei gameti di una
struttura e una fisiologia in grado di conciliare tre funzioni:
• sopravvivere in ambienti diversi da quello delle gonadi;
• riconoscere cellule omologhe dell’altro sesso e cooperare in eventi legati alla fecondazione;
• fornire adeguate quantità di materiale nucleare e citoplasmatico per lo sviluppo del nuovo organismo.
OVOGENESI
Nei Vertebrati l’ovogenesi è di tipo follicolare in
quanto, terminata la fase di moltiplicazione, ogni
ovogonio si associa a cellule somatiche di forma
appiattita per formare il follicolo ovarico
primordiale. Poco prima della nascita tutti gli
ovogoni si differenziano in ovociti primari:
duplicano il DNA, iniziano la fase di
accrescimento e entrano nella profase I della
meiosi. Poco prima o subito dopo la nascita, a
seconda della specie, ogni ovocita entra in una
prolungata fase di riposo allo stadio di diplotene
della profase I, detta dictiotene. Essa dura fino
alla maturità sessuale dell’animale, quando lo
sviluppo riprende per induzione da parte delle
gonadotropine. Nei Mammiferi il processo di
ovogenesi procede in parallelo col processo di
follicologenesi,
pertanto,
durante
l’attività
sessuale delle diverse specie animali, a regolari
intervalli di tempo, avviene la maturazione di uno
o più follicoli e dei relativi ovociti. Inoltre, sia
durante la fase proliferativa, che durante quella di
riposo, una cospicua quota di follicoli primordiali
va incontro ad atresia (nella specie umana da
2000 protogoni migranti dal sacco vitellino, si
arriva a 6-7 milioni di ovogoni al 5° mese di vita
intrauterina, mentre alla pubertà la donna ha da
200.000 a 400.000 ovociti; in una vitella di tre
mesi sono presenti circa 75.000 follicoli, in una
vacca di tre anni circa 20.000, in una di nove anni
circa 2.500).
follicolo atresico
Follicolo di Graaf
ovulazione
corpo luteo
follicolo secondario
follicolo primario
follicolo primordiale
corpo luteo in regressione
FECONDAZIONE
La fecondazione è il processo attraverso cui lo spermatozoo capacitato e l’ovocita si fondono per
formare lo zigote, che rappresenta il nuovo individuo.
La fecondazione consente il ripristino del numero diploide di cromosomi e la determinazione del sesso
dell’individuo; inoltre è fondamentale per la variabilità biologica grazie all’integrazione delle
caratteristiche ereditarie materne e paterne. L’integrazione del materiale genetico paterno e materno dei
due pronuclei viene indicata come singamia.
La fecondazione può essere esterna o interna. Nei Mammiferi è interna e nonostante durante il coito
vengano deposti milioni di spermatozoi nelle vie genitali femminili, solo alcune centinaia raggiungono la
sede della fecondazione ed uno solo partecipa alla singamia.
Il
coinvolgimento
di
più
spermatozoi nella fecondazione,
definito polispermia, è tollerato in
Pesci, Anfibi e Uccelli, anche se
gli spermatozoi in eccesso
comunque degenerano. Invece
nei Mammiferi, la presenza di
spermatozoi soprannumerari è
un evento anomalo che porta alla
morte dell’embrione.
Ripresa della
meiosi
Corona radiata
Zona pellucida
Granuli corticali
Reazione
acrosomiale
Membrana
ooplasmatica
Scoppio dei granuli
corticali
Ingresso dello
spermatozoo
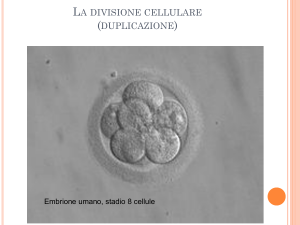
SEGMENTAZIONE
La segmentazione è l’evento che porta il nuovo individuo dalla condizione di
unicellularità alla condizione di pluricellularità, attraverso una serie di divisioni
mitotiche che si interrompono in genere quando viene ripristinato il rapporto
nucleo/citoplasmatico tipico della specie.
L’embrione entra in segmentazione allo stadio di zigote e al termine del
processo è divenuto una blastula formata da cellule chiamate blastomeri.
La modalità attraverso cui avviene la segmentazione è legata alla quantità di
tuorlo presente nell’uovo. Uova con scarsa o moderata quantità di tuorlo
segmentano completamente (segmentazione oloblastica), mentre uova con
grande quantità di tuorlo segmentano parzialmente (segmentazione
meroblastica).
Il tuorlo è un adattamento evolutivo dell’embrione che pertanto può svilupparsi senza assunzione esterna
di cibo.
Gli animali che hanno uova con poco tuorlo (oligolecitiche), come il riccio di mare (Echinoderma) o
l’Anfiosso (Cordato), hanno uno sviluppo embrionale molto breve con la formazione di una larva in grado
di muoversi e nutrirsi.
Passando ai Vertebrati, dagli Anfibi ai Pesci e poi ai Rettili, si osserva un progressivo aumento di tuorlo
che si accompagna alla riduzione e poi alla scomparsa dello stadio larvale. Infatti, alcuni Anfibi con uova
contenenti una discreta quantità di tuorlo (mesolecitiche), presentano segmentazione totale e formano un
girino come stadio larvale; altri Anfibi adattati alle zone desertiche, con uova contenenti grandi quantità di
tuorlo (macrolecitiche) come quelle dei Pesci e dei Sauropsidi, hanno segmentazione parziale e
l’embrione si può sviluppare interamente a spese del tuorlo. Per contro i Mammiferi Placentati, con uova
prive di tuorlo (alecitiche), hanno segmentazione totale ma sviluppano, accanto all’embrione, la placenta,
un annesso che, attraverso scambi materno-fetali, provvede al nutrimento dello stesso.
L’echidna trattiene l’uovo nell’utero fino
allo stadio di morula e successivamente
viene deposto e sistemato in una tasca
cutanea dove dopo la schiusa, il piccolo
viene allattato
L’ornitorinco depone le uova (1-3) in una tana
nella quale dopo la schiusa i piccoli
permangono per circa due settimane allattati
dalla madre
I Mammiferi sono considerati filogeneticamente derivati da
antenati correlati ai Rettili primitivi che avevano probabilmente
uova macrolecitiche simili a quelle degli attuali Sauropsidi.
Durante la filogenesi i Mammiferi sono diventati vivipari in
quanto la madre forniva nutrimento all’embrione attraverso la
placenta. Non essendoci necessità di tuorlo ai fini nutrizionali,
questo è progressivamente scomparso e le uova sono divenute
più piccole. Questo aspetto dello sviluppo evolutivo può
facilmente essere evidenziato nelle tre sottoclassi dei
Mammiferi:
Prototeri o Monotremi
Metateri o Marsupiali
Euteri o Placentati
Infatti i Prototeri depongono uova macrolecitiche ed il tuorlo
costituisce il nutrimento dell’embrione.
I Metateri sviluppano uova con moderato contenuto di tuorlo;
quest’ultimo viene però espulso per cui il nutrimento
dell’embrione è sostenuto, in utero, da una pseudo-placenta.
Infine gli Euteri hanno uova prive di tuorlo ed il nutrimento
dell’embrione in utero è sostenuto esclusivamente dalla
placenta.
In parallelo alla diminuzione del tuorlo, si assiste al passaggio
da una segmentazione parziale ad una segmentazione totale.
GASTRULAZIONE
In tutti i Vertebrati al termine della segmentazione si forma una blastula, più o meno cava e colma di liquido circondata
da cellule organizzate in un foglietto detto blastoderma. Tali cellule vanno incontro ad una serie di spostamenti, correlati
a modificazioni morfologiche, che comportano anche un profondo cambiamento della forma della blastula.
La gastrulazione è il processo che porta al riordino delle cellule del disco embrionale prima in due
(stadio didermico) e poi in tre (stadio tridermico) strati concentrici denominati foglietti germinativi che
dall’esterno all’interno sono rappresentati da ectoderma, mesoderma ed endoderma, dai quali
deriveranno tutti i tessuti e gli organi dell’individuo.
Il riordino delle cellule avviene attraverso movimenti morfogenetici irreversibili ed il processo non comporta in genere
aumento volumetrico dell’embrione, anche se l’attività mitotica è comunque intensa. Si assiste però a modificazioni
metaboliche in relazione all’iniziale differenziamento.
Sebbene i tipi di gastrulazione varino notevolmente nelle diverse specie animali, anche in relazione al tipo di blastula
che si è formata alla fine del processo di segmentazione, i meccanismi primari di tale processo sono conservati
filogeneticamente e sono ripetutamente utilizzati in varie tappe dello sviluppo.
La gastrulazione è il risultato di una combinazione dei seguenti meccanismi di movimento:
1. espansione di cellule in superficie in modo da ricoprire e circondare quelle più interne o epibolia;
2. movimento di divisione di uno strato di cellule in due lamine parallele o delaminazione;
3. movimenti di aggregazione cellulare o di addensamento conseguenti ad aumento di adesione tra le
cellule o ad aumento locale delle divisioni mitotiche.
4. spostamento di cellule dalla superficie verso l’interno.
Questo spostamento di cellule verso l’interno può avvenire per:
• invaginazione, ripiegamento di una porzione del blastoderma o di una lamina cellulare verso
l’interno, simile all’infossamento di una palla di gomma quando viene schiacciata in un punto
circoscritto o embolia;
• involuzione, ripiegamento e scorrimento di cellule in modo da accollarsi alla superficie interna dello
strato superficiale;
• ingressione o immigrazione, migrazione di singole cellule del blastoderma all’interno dell’embrione.
Differenziamento dei foglietti embrionali
Ectoderma
E’ il foglietto embrionale più
esterno, ha la forma di un disco
epiteliale in continuità con
l’ectoderma amniotico e si divide
in tre territori principali:
l’ectoderma di rivestimento
il tubo neurale
la cresta neurale
Mesoderma
E’
il
foglietto
embrionale intermedio
e si divide in cinque
territori principali:
Placca precordale
Cordomesoderma
Mesoderma
parassiale
Mesoderma
intermedio
Mesoderma laterale
Endoderma
L’endoderma comincia
il suo differenziamento
formando
il
tubo
intestinale primitivo.
(fegato, pancreas)
Durante lo sviluppo si succedono due periodi indicati
come periodo embrionale e periodo fetale.
Il periodo embrionale va dalla fecondazione al
momento in cui il nuovo organismo è nettamente
definito nel suo aspetto esteriore e nelle sue parti
costitutive.
A questo segue il periodo fetale, che si estende fino
al termine della gravidanza, nel quale si perfeziona e
si completa l’organogenesi.
ANNESSI EMBRIONALI: aspetti evolutivi
Gli annessi embrionali sono strutture che si formano da estensioni dei foglietti embrionali nella
cosiddetta area extraembrionale e permettono lo sviluppo dell’embrione e del feto.
Nei Vertebrati acquatici a fecondazione esterna, come la maggior parte dei Pesci Teleostei e degli
Anfibi, lo sviluppo embrionale avviene all’interno della membrana dell’uovo circondata da un esile
strato gelatinoso, cosicchè gli scambi gassosi e l’allontanamento dei cataboliti avvengono facilmente
tra l’embrione e l’acqua circostante. Il tuorlo, necessario alla nutrizione dell’embrione, viene
inglobato nel citoplasma delle cellule dell’endoderma (Anfibi) o avvolto dai tre foglietti endoderma,
mesoderma e ectoderma (Teleostei) che insieme formano la parete del cosiddetto sacco del tuorlo o
vitellino. Questo costituisce l’unico annesso presente nei Pesci.
L’espansione dei Vertebrati sulle terre emerse ha richiesto, tra i tanti adattamenti, che gli embrioni
fossero particolarmente protetti dalla disidratazione e dalla forza di gravità. E’ stato quindi ricreato
un ambiente acquatico dove gli embrioni potessero svilupparsi: l’amnios con il liquido amniotico in
esso contenuto. Pertanto i Rettili, gli Uccelli e i Mammiferi possiedono questo nuovo annesso e
sono definiti amnioti.
Inoltre i Vertebrati terrestri ovipari (la maggior parte dei Sauropsidi e i Mammiferi Monotremi) hanno
sviluppato degli involucri a funzione protettiva e nutritizia attorno alle uova, quali l’albume, la
membrana testacea e il guscio calcareo. Tali involucri, rendendo difficoltosa l’eliminazione delle
sostanze di scarto e la respirazione, hanno determinato la necessità della formazione di altri due
annessi embrionali: il corion e l’allantoide.
La viviparità, che comporta l’instaurarsi di un rapporto tra l’ovidutto o l’utero materno e l’embrione, è
sporadica nei Vertebrati acquatici (alcuni squali) e nei Rettili (camaleonti), ma raggiunge il successo
evolutivo nei Mammiferi Metateri e Euteri, con rapporti sempre più stretti tra madre ed embrione. Tali
rapporti sono resi possibili da profonde modificazioni del corion e della mucosa uterina che insieme
costituiscono la placenta, organo che attua i necessari scambi metabolici tra madre ed embrione
rendendo pertanto inutile la presenza di tuorlo nelle uova di questi animali.