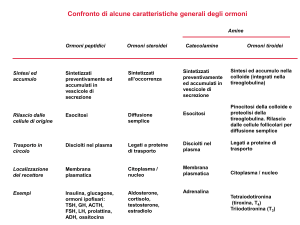
ORMONI
Gli ormoni vegetali sono molecole segnale presenti in tracce.
Variazioni della loro concentrazione e della sensibilità dei
tessuti bersaglio mediano tutta una serie di processi di
sviluppo delle piante.
Le cellule bersaglio sono quelle che possiedono i recettori
per l’ormone. I recettori sono molecole, di solito proteine, che
legando l’ormone attivano una serie di eventi, la cosiddetta
via di trasduzione del segnale, che porta alla risposta
fisiologica.
Le piante non possono muoversi, quindi sono soggette a
interazioni con fattori ambientali da cui non possono
sfuggire, quindi i processi di sviluppo devono tenere conto
anche di queste interazioni.
ORMONI
La principale attenzione è rivolta ai sei principali tipi di
ormoni conosciuti (auxine, gibberelline, citochinine, acido
abscissico, etilene, brassinosteroidi) ma altri composti sono
di recente stati aggiunti alla lista di quelli che hanno una
funzione regolatrice nello sviluppo delle piante (es.
poliammine, acido salicilico, acido giasmonico,
oligosaccaridi, sistemina)
Le vie di trasduzione del segnale nelle piante sono per molti
versi simili a quelle presenti negli animali.
ORMONI
AUXINE
ORMONI
AUXINE
ORMONI
BIOSINTESI DELL’ACIDO 3-INDOLACETICO
ORMONI
BIOSINTESI DELL’ACIDO 3-INDOLACETICO
ORMONI
BIOSINTESI DELL’ACIDO 3-INDOLACETICO
ORMONI
INATTIVAZIONE DELL’ACIDO 3-INDOLACETICO
ORMONI
DEGRADAZIONE DELL’ACIDO 3-INDOLACETICO
ORMONI
DOSAGGIO DELLE AUXINE
- TEST DI CURVATURA DEL COLEOTTILE D’AVENA
- TEST DI ALLUNGAMENTO DI SEGMENTI
ORMONI
DOSAGGIO DELLE AUXINE
- TEST RIA
- TEST ELISA
- TLC o HPLC o GC + MS
ORMONI
TRASPORTO DELL’ACIDO 3-INDOLACETICO
IL TRASPORTO DELL’AUXINA E’ POLARE
ORMONI
TRASPORTO DELL’ACIDO 3-INDOLACETICO
IL TRASPORTO DELL’AUXINA E’ POLARE
ORMONI
TRASPORTO DELL’ACIDO 3-INDOLACETICO
IL TRASPORTO DELL’AUXINA E’ POLARE
NEL FUSTO IL MOVIMENTO E’ BASIPETO
da cellule meno differenziate a più differenziate
NELLA RADICE IL MOVIMENTO E’ ACROPETO
da cellule più differenziate a meno differenziate
ORMONI
TRASPORTO DELL’ACIDO 3-INDOLACETICO
AUX1
PIN
TIBA - NPA
ORMONI
PERCEZIONE E TRASDUZIONE DEL SEGNALE
DELL’AUXINA
ORMONI
EFFETTI DELLE AUXINE
- DISTENSIONE CELLULARE (allungamento, ingrossamento)
- FOTOTROPISMO
- GRAVITROPISMO
- DOMINANZA APICALE
- RITARDO DELL’ABSCISSIONE
- SVILUPPO DELLE GEMME FIORALI
-FORMAZIONE DELLE RADICI
LATERALI
- INDUZIONE DELL’ATTIVITA’
DEL CAMBIO C.V.
LE AUXINE SINTETICHE FUNZIONANO DA ERBICIDI
ORMONI
DISTENSIONE CELLULARE
La stimolazione
richiede energia
È inibita da
cicloesimmide e da
inibitori di RNA
L’auxina esercita effetto su segmenti
piuttosto che su piante intere
Le cellule della radice sono più
sensibili: l’effetto diventa inibitorio a
concentrazione minore
Il lag è di circa 10 min ed è
indipendente da altri fattori
ORMONI
DISTENSIONE CELLULARE - MECCANISMO
L’auxina aumenta la plasticità della parete cellulare
Secondo l’ipotesi della crescita acida l’auxina induce la
crescita attivando una pompa protonica del plasmalemma
che estrude protoni nella parete acidificandola e aumentando
la sua plasticità
I tamponi acidi quindi
dovrebbero produrre lo
stesso effetto, almeno a
breve termine
ORMONI
DISTENSIONE CELLULARE - MECCANISMO
Inoltre composti diversi dall’auxina che provocano
estrusione di protoni dovrebbero stimolare la crescita
esattamente come l’auxina
Lo fa la FUSICOCCINA, una fitotossina fungina che
stimola l’estrusione rapida di protoni e induce
l’accrescimento momentaneo in sezioni di coleottile
FUSICOCCIN: WHO IS IT?
ORMONI
DISTENSIONE CELLULARE - MECCANISMO
Quale conseguenza provoca l’acidificazione della parete?
I legami idrogeno che tengono legate le catene polisaccaridiche potrebbero essere indeboliti da proteine di parete
definite ESPANSINE: la partecipazione di proteine sarebbe
dimostrata dal blocco della stimolazione causato da
condizioni che denaturano le proteine (calore, proteasi)
ORMONI
DISTENSIONE CELLULARE - MECCANISMO
ORMONI
FOTOTROPISMO
MODELLO CHOLODNY-WENT
ORMONI
FOTOTROPISMO
Il fototropismo è controllato dalla luce blu. I fotorecettori
sono due: fototropina 1 e 2. Si tratte di due enzimi
appartenenti alla famiglia delle proteina chinasi con azione
autofosforilante. Il gradiente di fosforilazione sembra indurre
la migrazione dell’auxina verso il lato in ombra.
ORMONI
FOTOTROPISMO
Il fototropismo è controllato dalla luce blu. I fotorecettori
sono due: fototropina 1 e 2. Si tratte di due enzimi
appartenenti alla famiglia delle proteina chinasi con azione
autofosforilante. Il gradiente di fosforilazione sembra indurre
la migrazione dell’auxina verso il lato in ombra.
ORMONI
GRAVITROPISMO
Il modello
Cholodny-Went
può essere applicato anche al
gravitropismo
La percezione
dello stimolo
gravitropico è
nella cuffia
della radice
ORMONI
GRAVITROPISMO
Il gravitropismo della
radice è legato al
movimento degli
statoliti (amiloplasti)
ORMONI
GRAVITROPISMO
Il ruolo dell’auxina
ORMONI
DOMINANZA APICALE
Ancora incerto il meccanismo con cui
l’auxina determina la dominanza
apicale. Forse reprime geni della
sintesi di citochinine.
Recentemente sono stati individuati
prodotti derivati da carotenoidi
(strigolattoni) che agendo insieme
all’auxina inibirebbero lo sviluppo
delle gemme laterali.
ORMONI
RADICAZIONE
Individuati in
Arabidopsis thaliana
geni che controllano la
formazione di radici
laterali
ORMONI
GIBBERELLINE
Originariamente scoperte
in colture di riso in
Giappone come sostanza
prodotta da un fungo
patogeno
Gibberella fujikuroi
(Fusarium moniliforme)
ORMONI
BIOSINTESI DELLE GIBBERELLINE
ORMONI
BIOSINTESI DELLE GIBBERELLINE
ORMONI
INATTIVAZIONE DELLE GIBBERELLINE
Il controllo del livello di gibberelline libere è più che altro
basato su reazioni di coniugazione con glucosio
(glucosilazione)
DETERMINAZIONE DELLE GIBBERELLINE
HPLC-MS o saggi biologici
ORMONI
EFFETTI DELLE GIBBERELLINE
Hanno effetto su
piante intere
Stimolazione della
crescita di mutanti
nani
Viene stimolata sia la
distensione che la
divisione cellulare. La
distensione è dovuta
ad allentamento della
parete ma non con
acidificazione
dell’apoplasto.
ORMONI
EFFETTI DELLE GIBBERELLINE
Induzione della
fioritura in
sostituzione del
fotoperiodo lungo
ORMONI
EFFETTI DELLE GIBBERELLINE
Induzione della
fioritura in
sostituzione delle
basse temperature
(vernalizzazione)
ORMONI
EFFETTI DELLE GIBBERELLINE
Induzione dello
allungamento della
guaina fogliare in
piantine di riso
ORMONI
EFFETTI DELLE GIBBERELLINE
Mobilizzazione delle riserve amilifere delle cariossidi di
graminacee
ORMONI
EFFETTI DELLE GIBBERELLINE
Mobilizzazione delle riserve
amilifere delle cariossidi di
graminacee
ORMONI
EFFETTI DELLE GIBBERELLINE
ORMONI
EFFETTI DELLE GIBBERELLINE
Produzione di frutti
partenocarpici
ORMONI
CITOCHININE
Ormoni particolarmente deputati al controllo della divisione
cellulare ma che esercitano anche altri effetti quali il ritardo
della senescenza e l’annullamento della dominanza apicale
ORMONI
CITOCHININE: BIOSINTESI
ORMONI
CITOCHININE: METABOLISMO
ORMONI
CITOCHININE: EFFETTI
Le citochinine accelerano il ciclo cellulare agendo sui fattori
coinvolti nelle diverse fasi del ciclo (cicline e chinasi ciclinadipendenti).
ORMONI
CITOCHININE: EFFETTI
Sviluppo dei calli
ORMONI
CITOCHININE: EFFETTI
effetto del rapporto auxina/citochinine
ORMONI
CITOCHININE: EFFETTI
Gli effetti del rapporto auxina/citochinine sono confermati da
esperimenti con Agrobacterium tumefaciens
Tumore del colletto provocato
da Agrobacterium tumefaciens
ORMONI
INDUZIONE DEL TUMORE DA AGROBACTERIUM
ORMONI
IL T-DNA DI AGROBACTERIUM
ORMONI
AGROBACTERIUM COME VETTORE DI
TRASFORMAZIONE DELLE PIANTE
ORMONI
CITOCHININE: EFFETTI
ritardo della senescenza
pianta transgenica di
tabacco che esprime il
gene ipt di
Agrobacterium sotto il
controllo del promotore
CaMV 35S.
ORMONI
CITOCHININE: EFFETTI
Frutti di pomodoro trasformati con il gene ipt sotto il
controllo del promotore specifico 2A11. Paradossalmente,
dato che le citochinine ritardano la senescenza, le aree rosse
contengono sei volte più citochinine delle aree verdi
ORMONI
CITOCHININE: EFFETTI
Convogliamento di nutrienti
ORMONI
CITOCHININE: EFFETTI
Sviluppo dei cloroplasti
ORMONI
CITOCHININE: EFFETTI
Espansione dei cotiledoni