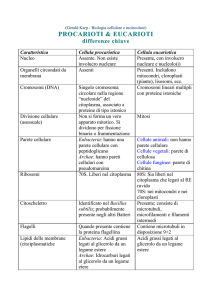
 Cellula vegetale")
04/04/17
Rispetto alla cellula animale, le
principali peculiarità della cellula
vegetale sono:
(a) un involucro rigido esterno
al plasmalemma (parete),
(b) specifiche connessioni
intercellulari (plasmodesmi),
(c) un apparato vacuolare di
grandi dimensioni,
(d) organelli detti plastidi, tra
cui i cloroplasti,
(e) un citoscheletro che mostra
architettura e dinamica
particolari
(f) gliossisomi.
La presenza della parete cellulare nelle piante impone che tutte le
cellule siano interconnesse da ponti citoplasmatici: i
plasmodesmi. Questi sono costituiti da un tubulo di reticolo
endoplasmatico rivestito dal plasmalemma. Le dimensioni
massime delle molecole che normalmente passano attraverso i
plasmodesmi è circa 1000 Da, lo stesso che per le gap junctions
degli animali.
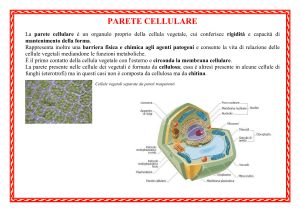
La parete cellulare
E un rivestimento continuo, più o meno rigido,
assemblato esternamente alla membrana cellulare
utilizzando precursori prodotti dalla cellula. Una parete
è presente in molte cellule sia di organismi procariotici
che eucariotici:
- Batteri (Gram + e Gram -) principale componente:
peptidoglicano mureina
- Molte alghe (Rodofite, Feofite, Clorofite) - principale
componente: cellulosa
- Funghi - principale componente: chitina
- Piante - principale componente: cellulosa
Batterio
Fungo
PARETE CELLULARE
Funzioni della parete cellulare:
1) protezione
Perché le cellule delle piante hanno la
parete cellulare?
La principale funzione della parete cellulare è quella
protettiva. La protezione cellulare è fondamentale nelle
piante, perché esse dipendono da nutrienti presenti
nell ambiente in forma dispersa (luce, CO2, ioni minerali)
ed hanno perciò l esigenza di stabilire un ampia
superficie di contatto con l esterno. Gli animali sono
eterotrofi, dipendono da nutrienti presenti in forma
concentrata (materiale organico) e la maggior parte delle
loro cellule non è a contatto con l esterno ma con un
ambiente interno molto stabile e controllato.
PARETE CELLULARE
2) sostegno (scheletro)
3) trasporto
Parete cellulare delle piante ha tre componenti
fondamentali: (1) cellulosa – componente fibrillare
- (associata con emicellulose), (2) polisaccaridi
pectici, o pectine, (3) proteine strutturali.
Sono presenti anche numerosi enzimi, che rendono la parete
una parte altamente dinamica e reattiva della cellula vegetale.
Spesso, nei rivestimenti esterni della pianta presenti sostanze
lipidiche (cutina, suberina (nel sughero) e cere) per la
riduzione di perdita di acqua.
PARETE CELLULARE
1
04/04/17
La cellulosa, un fondamentale componente della
parete cellulare della cellula vegetale è un
omopolisaccaride costituito da glucosio
La cellulosa
Singola catena di cellulosa
(poli- β-1-4-glucano) e catene
associate tramite legami
idrogeno in una microfibrilla.
Grado di polimerizzazione: da
2000 a 15000 unità di glucosio.
Nelle piante una microfibrilla
consiste mediamente di 36
molecole di cellulosa.
PARETE CELLULARE
Le pectine sono polisaccaridi eterogenei acidi o neutri
PARETE CELLULARE
Le pectine
(costituiti da zuccheri diversi, il più comune acido
galatturonico). In presenza di ioni calcio, le pectine formano
gel compatti e fortemente adesivi.
Nella frutta sono abbondanti nelle pareti cellulari e, durante
la maturazione, vengono idrolizzate da enzimi come le
pectinasi.
Vengono utilizzate nell industria alimentare come
gelificante nella preparazione di marmellate.
ACIDO GALATTURONICO
PARETE CELLULARE
Le emicellulose sono polisaccaridi ramificati di matrice che
interagiscono con la cellulosa mediante legami idrogeno. Le
emicellulose costituiscono circa il 40-45% delle pareti cellulari
primarie.
PARETE CELLULARE
I diversi componenti della parete sono assemblati in
una struttura composita costituita dalla parte fibrillare
(cellulosa) e da una matrice (pectine). Le emicellulose,
costituite da differenti monosaccaridi e a struttura non
fibrillare ma ramificata, stabiliscono la connessione fra
fibrille di cellulosa e matrice.
Modello di parete nelle
angiosperme dicotiledoni
PARETE CELLULARE
Dettaglio della parete di
cellula vegetale
PARETE CELLULARE
2
04/04/17
La parete durante lo sviluppo cellulare
La parete nelle giovani cellule in divisione è sottile ed estensibile
durante la crescita che accompagna il differenziamento cellulare. Il
suo spessore resta però invariato perché la cellula continua a
deporre nuovo materiale di parete durante l espansione. La parete
formata nel corso della fase di espansione cellulare è detta PARETE
PRIMARIA, comprende un sottile strato centrale, LAMELLA
MEDIANA la quale ricca di pectine funge da collante fra le cellule.
Parete secondaria
Una volta raggiunte le dimensioni definitive, la cellula completa
il proprio differenziamento e inizia a funzionare come cellula
adulta. La maggior parte delle cellule del corpo della pianta
possiede solo una parete primaria. Alcuni tipi cellulari, però,
completano il differenziamento deponendo una parete
secondaria all interno della parete primaria. Si tratta in
particolare di cellule specializzate nella funzione di sostegno e
conduzione. Queste cellule spesso dopo aver depositato la
parete secondaria muoiono.
Lamella
mediana
PARETE CELLULARE
PARETE CELLULARE
PLASTIDI
I plastidi sono organuli tipici delle piante,
delimitati da due membrane e contenenti DNA.
Internamente ai plastidi si differenziano sistemi di
membrane.
Oltre ai cloroplasti le piante possiedono plastidi non
fotosintetici, localizzati in tessuti e organi specifici.
Sono presenti cloroplasti anche nei protisti
fotosintetici (alghe).
I plastidi maturi sono classificati in base ai pigmenti o
altre molecole presenti:
-CLOROPLASTI
-CROMOPLASTI
-LEUCOPLASTI
PLASTIDI
I plastidi derivano sempre da plastidi
preesistenti. Nelle piante le giovani cellule degli
apici meristematici contengono piccoli plastidi
immaturi, detti proplastidi. I proplastidi si
differenziano in cloroplasti, amiloplasti o
cromoplasti o altri tipi di plastidi specializzati.
I proplastidi possono
contenere numerose copie
(50-150) del genoma
plastidiale. I proplastidi si
dividono e il numero di
copie del genoma per
plastidio tende a diminuire
durante il differenziamento
PLASTIDI
Nel corso dello sviluppo della pianta sono frequenti
interconversioni di un tipo plastidiale in un altro. Per esempio, la
buccia verde dei frutti assume diverse colorazioni nel corso della
maturazione a causa della conversione dei cloroplasti in
cromoplasti. L inverdimento dei tuberi della specie Solanum
tuberosum, causato dall esposizione alla luce, riflette la
conversione degli amiloplasti in cloroplasti. In generale, i
cromoplasti rappresentano una forma senescente di plastidio, non
più convertibile.
I plastidi contengono DNA circolare di tipo procariotico
(130-150 Kb nelle piante). Questo DNA contiene alcune
decine di geni che codificano per RNA ribosomiale e per
alcune proteine plastidiali, tra cui la subunità maggiore
della Rubisco. La maggior parte delle proteine
plastidiali, però, è codificata da geni nucleari.
Origine dei cloroplasti: teoria
endosimbiontica
amiloplasto
cromoplasto
cloroplasto
PLASTIDI
Parete
secondaria
proplastidio
PLASTIDI – TEORIA ENDOSIMBIONTICA
3
04/04/17
Il cloroplasto (e gli organelli simili) probabilmente deriva
da un cianobatterio simbionte acquisito per fagocitosi dal
progenitore unicellulare delle alghe verdi (da cui le
piante derivano). L acquisizione del cloroplasto è
avvenuta dopo che, attraverso un processo simile, erano
stati acquisiti i mitocondri (endosimbiosi).
Eucariote
fagotrofo
anaerobio
Batteri aerobi,
diventano mitocondri,
presenti in (quasi)
tutti gli eucarioti oggi
viventi
Cianobatteri,
diventano cloroplasti,
presenti negli
eucarioti fotosintetici
insieme ai mitocondri
Genoma di cloroplasti e mitocondri
Mitocondri e cloroplasti posseggono un proprio DNA
e propri ribosomi per cui sono in grado di
sintetizzare i vari tipi di RNA e di conseguenza,
anche se un numero molto ridotto, di proteine.
Esistono inibitori specifici delle due sintesi
proteiche. Quella citosolica è inibita dalla
cicloeximide mentre la sintesi proteica degli
organuli no.
La sintesi proteica dei mitocondri e dei cloroplasti è
inibita da cloroamfenicolo, tetraciclina ed
eritromicina, mentre quella citosolica non lo è.
PLASTIDI – TEORIA ENDOSIMBIONTICA
Una caratteristica importante del
genoma dei cloroplasti e dei
mitocondri è che esso è costituito da
DNA circolare (che nei mitocondri di
cellule vegetali è molto più grande
rispetto a quello dei mitocondri di
cellule animali perché contiene molti
introni).
Genoma dei cloroplasti
Il genoma dei cloroplasti è
fortemente rassomigliante a quello
dei batteri, esso contiene circa 120
geni che codificano proteine dei
fotosistemi I e II, dell ATP-sintetasi
e componenti di complessi
multienzimatici.
PLASTIDI – TEORIA ENDOSIMBIONTICA
Il doppio strato lipidico ha composizione asimmetrica
che differisce a seconda del tipo di membrana
PLASTIDI – TEORIA ENDOSIMBIONTICA
PLASTIDI – TEORIA ENDOSIMBIONTICA
I cloroplasti
nelle piante si trovano soprattutto nel tessuto fotosintetico
(clorenchima) delle foglie e dei giovani fusti. Il cloroplasto
contiene materiale trasparente, detto STROMA, e, immersi in
questo, numerosi sacculi membranosi appiattiti, i TILACOIDI, la
cui organizzazione spaziale è diversa nei vari gruppi di eucarioti
fotosintetici. Nelle piante i tilacoidi sono disposti in pile, visibili
anche al microscopio ottico e dette grani.
PLASTIDI – CLOROPLASTI
4
04/04/17
La principale, ma non unica, funzione del cloroplasto è la fotosintesi,
cioè la riduzione del biossido di carbonio a glucosio usando come
fonte di energia la luce.
nCO2 + nH2O → (CH2O)n +n O2
- CH2O solitamente n=6 e il monosaccaride che si forma è il glucosio
La fotosintesi si articola in DUE FASI. La prima, detta FASE LUMINOSA, compie
la conversione dell energia luminosa in energia chimica (sotto forma di ATP e
NADPH). La fase luminosa ha sede nei tilacoidi e ad essa partecipano
direttamente i pigmenti fotosintetici. Nella seconda fase, detta FASE OSCURA,
l ATP e il NADPH sono usati per ridurre il biossido di carbonio a glucosio. La
fase oscura è catalizzata da enzimi presenti nello stroma.
Il glucosio prodotto per fotosintesi è accumulato nel
cloroplasto sotto la forma, osmoticamente inattiva, di
amido, secondo la reazione:
n glucosio + n ATP → (α-1-4 glucosio)n + n ADP + n Pi
L amido accumulato nel cloroplasto durante il giorno è
detto amido primario.
Amido
primario
PLASTIDI – CLOROPLASTI
PLASTIDI – CLOROPLASTI
AMILOPLASTI
L amido primario, accumulato nel cloroplasto
durante il giorno, viene riconvertito in zucchero
solubile (saccarosio, disaccaride costituito da
glucosio+fruttosio) durante la notte, trasportato
nei tessuti di riserva (soprattutto fusto e radice)
e qui ricondensato come amido negli amiloplasti
(amido secondario). Gli amiloplasti sono plastidi
privi di pigmenti fotosintetici, specializzati
nell accumulo di amido. Sono abbondanti nei
tessuti di riserva di fusto, radice, semi.
ACCUMULO DI AMIDO
Sezione trasversale di una
giovane foglia colorata con il
reattivo di Wagner (IodioIoduro di potassio)
Cellule in coltura
Gli amiloplasti fanno parte dei plastidi non
pigmentati detti genericamente leucoplasti.
Sezione di radice
PLASTIDI – AMILOPLASTI
PLASTIDI – AMILOPLASTI
L AMIDO
L amido è composto da due polimeri: l amilosio (che ne costituisce circa il
20%) e l'amilopectina (circa l'80%). In entrambi i casi si tratta di polimeri
del glucosio, che si differenziano l'uno dall'altro per la struttura. L'amilosio è
un polimero lineare che tende ad avvolgersi ad elica, in cui le unità di
glucosio sono legate tra loro con legami glicosidici (1 4) (tra C1 di una
unità e C4 dell'unità successiva). La doppia elica è stabilizzata da legami a
idrogeno. L'amilopectina è un polimero ramificato che presenta catene di
struttura simile all'amilosio che si dispongono a formare una struttura
ramificata; ogni 24-30 unità di glucosio, infatti, si innestano catene laterali
attraverso legami (1 6).
AMILOSIO E AMILOPECTINA
PLASTIDI – AMILOPLASTI
PLASTIDI – AMILOPLASTI
5
04/04/17
I Cromoplasti
sono plastidi non fotosintetici che accumulano grandi
quantità di carotenoidi gialli, arancione o rossi. I
cromoplasti conferiscono uno di questi colori alle parti
della pianta in cui si trovano (di solito fiori, foglie
senescenti, radici e frutti).
Possono svilupparsi dai cloroplasti verdi già esistenti
mediante perdita delle membrane interne dei
cloroplasti e di clorofilla e accumulo dei carotenoidi,
come avviene durante la maturazione di molti frutti.
Petalo di ranuncolo
PLASTIDI – CROMOPLASTI
PLASTIDI – CROMOPLASTI
CAROTENOIDI
CAROTENOIDI
In natura sono presenti più di 500 diversi carotenoidi.
Anche alcuni batteri e funghi sintetizzano carotenoidi.
Tutte le molecole di carotenoidi sono molecole idrofobiche,
presentano una lunga catena idrocarburica e ad entrambe
le estremità un anello a 6 atomi di carbonio. In totale le
molecole di carotenoidi hanno 40 atomi di carbonio.
Esistono due classi di carotenoidi: CAROTENI E
XANTOFILLE
I CAROTENI sono costituiti solo da atomi di carbonio e
idrogeno – il più comune è il β-carotene.
Le XANTOFILLE hanno anche uno o più atomi di ossigeno –
una delle molecole più comuni è la luteina.
http://science.jrank.org/pages/5303/Plant-Pigment-Carotenoids.html
CAROTENOIDI
I CROMOPLASTI
cellula dell'epidermide
PLASTIDI – CROMOPLASTI
6
04/04/17
CONVERSIONE DI CLOROPLASTI IN CROMOPLASTI
CONVERSIONE DI CLOROPLASTI IN CROMOPLASTI
Durante il processo di maturazione dei frutti,
contemporaneamente alla conversione di cloroplasti in
cromoplasti, si ha in genere, il consumo di amido
accumulato.
GLIOSSISOMI
GLIOSSISOMI
ORGANULI SPECIFICI
DELLE CELLULE
V E G E TA L I I N C U I
AVVIENE IL CICLO DEL
G L I O S S I L AT O : Q U I
AVVIENE PARTE
DEGRADAZIONE DEGLI
ACIDI GRASSI IN
CARBOIDRATI.
Piccoli organuli delle cellule vegetali scoperti solo nel
1961. Si possono considerare perossisomi specializzati
Presenti in particolar modo nei tessuti dove sono
accumulati acidi grassi (semi).
GLIOSSISOMI - CICLO DEL GLIOSSILATO
VACUOLI
Perché le cellule vegetali hanno grandi vacuoli?
I vacuoli sono vescicole delimitate da membrana detta TONOPLASTO
e ripiene di liquido acquoso detto SUCCO VACUOLARE. Le piante
usano i vacuoli come un mezzo per aumentare le dimensioni cellulari
e quindi accrescersi. La crescita per accumulo di acqua nelle cellule è
un sistema molto economico per produrre un ampia superficie di
contatto con l ambiente.
I vacuoli sono molto piccoli nella fase giovanile e si accrescono nel
corso del differenziamento cellulare.
Assumono un ruolo chiave durante la germinazione dei semi, consentendo in
questo modo di fornire glucosio quando la pianta non è ancora in grado di
produrlo mediante la fotosintesi. Cosi si può formare la radice e il germoglio.
VACUOLI
7
04/04/17
Le cellule vegetali sono di solito molto più
grandi delle cellule degli animali. Ciò è dovuto
soprattutto alla presenza dei vacuoli.
Un esempio: lo sviluppo del frutto di pesco
Cellule di frutto di
Prunus persica
fotografate allo stesso
ingrandimento in
momenti successivi
dello sviluppo. Le
divisioni cellulari si
arrestano quando la
giovane pesca ha le
dimensioni di circa 1
cm2 (a). Poi ad
aumentare sono solo
le dimensioni cellulari,
soprattutto per
l espansione del
vacuolo.
a
10-20 µm
30-100 µm
VACUOLI
VACUOLI
RUOLO di RISERVA di ACQUA:
Fenomeno della PLASMOLISI
IL TURGORE CELLULARE
La cellula delle piante, provvista di parete, assorbe acqua
fino ad una condizione di equilibrio, detta TURGORE
CELLULARE.
acqua
pura
La perdita d’acqua per osmosi nella cellula vegetale è nota
come PLASMOLISI. Durante la plasmolisi il plasmalemma
si stacca dalla parete tirando con sé i plasmodesmi.
VACUOLI – TURGORE CELLULARE
VACUOLI – TURGORE CELLULARE
Soluzione Esterna IPERTONICA
Soluzione
ISOTONICA
Il vacuolo presente nelle cellule delle piante
presenta sostanziali affinità con i lisosomi
delle cellule animali. Il succo vacuolare è una
soluzione di ioni e piccole molecole organiche
ed ha un pH decisamente acido (pH 3 – 5). Il
succo vacuolare, inoltre, contiene numerosi
enzimi litici capaci di idrolizzare proteine,
lipidi e acidi nucleici.
vacuolo
(pH acido)
citoplasma
(pH neutro)
parete
VACUOLI – TURGORE CELLULARE
VACUOLI
8
04/04/17
Come in altre membrane biologiche, le proteine
integrali presenti nel tonoplasto hanno una
distribuzione asimmetrica, sporgendo più sul lato
vacuolare (V) che su quello citoplasmatico (C). Tra le
proteine integrali è compresa una pompa di
protoni (H+-ATPasi).
Le pompe H+-ATPasi e H+-PPasi idrolizzano
adenosina trifosfato (ATP) e pirofosfato
inorganico (PPi) generando una forza protonica
per il trasporto di ioni e metaboliti attraverso il
tonoplasto.
VACUOLI
VACUOLI
I vacuoli probabilmente hanno meccanismi di
formazione diversi in diversi tipi cellulari. In
molti casi essi appaiono derivare da piccoli
rigonfiamenti locali di cisterne di reticolo
endoplasmatico (provacuoli), che
successivamente si espandono. I vacuoli sono
perciò uno dei pochi organelli cellulari che
possono essere prodotti de novo.
Il vacuolo, oltre ad
accumulare l acqua in
eccesso, può funzionare
come sito di accumulo
di sostanze di riserva
(corpi proteici nei semi) o
sostanze di rifiuto (es.
cristalli di ossalato di
calcio).
VACUOLI
VACUOLI
Probabilmente nella stessa cellula possono essere presenti
tipi diversi di vacuolo. Questo è stato dimostrato in cellule
di semi, nelle quali si osservano grossi vacuoli pieni di
materiale proteico (V1) e numerosi piccoli vacuoli con un
contenuto acido (V2) .
VACUOLI
Il vacuolo può anche partecipare alla difesa
della pianta nei confronti di predatori erbivori
accumulando sostanze tossiche: METABOLITI
SECONDARI. Fra queste si annoverano
numerosi glicosidi
VACUOLI
9
04/04/17
MICROTUBULI E MICROFILAMENTI DI ACTINA - architettura e dinamica
molto particolari.
MICROFILAMENTI DI ACTINA
1- L interazione actina e miosina responsabile del movimento del citoplasma
- CICLOSI CITOPLASMATICHE- o di organelli come i cloroplasti per
posizionarli in funzione della intensità luminosa.
2- controllo della migrazione delle vescicole di esocitosi, processo
importante per la crescita apicale.
MICROTUBULI
1- si irradiano dal nucleo formando citoscheletro corticale
2- esistono numerosi MTOC, associati all involucro nucleare e forse al
plasmalemma
3- in interfase : CITOSCHELETRO CORTICALE orienta le microfibrille
di cellulosa della parete in espansione. Inizio mitosi: Citoscheletro
corticale sostituito da BANDA PREPROFASICA nella regione dove si
formerà il setto di divisione tra le due cellule figlie.
4- mitosi: FUSO MITOTICO ANASTRALE (per assenza di centrioli). Il
fuso mitotico controlla la ripartizione dei cromatidi nei due nuclei.
5- telofase: FRAGMOPLASTO in posizione centrale fra i due nuclei;
controlla la formazione del setto di divisione (nuova parete cellulare).
CITOSCHELETRO VEGETALE
Come negli animali e negli altri eucarioti, le cellule delle piante
contengono un citoscheletro di tubulina e di actina. L architettura
e la dinamica del citoscheletro nelle piante sono però molto
particolari.
Nelle cellule vegetali pienamente differenziate il citoplasma
contiene un reticolo di microfilamenti di actina, in parte legati al
plasmalemma, mentre sulla superficie degli organelli
citoplasmatici è presente una specifica forma di miosina.
L interazione ATP-dipendente tra actina e miosina è responsabile
del movimento del citoplasma (ciclosi citoplasmatiche) o di
specifici organelli (per es. i cloroplasti).
Microfilamento
di actina
plasmalemma
Le ciclosi citoplasmatiche sono regolarmente presenti nelle
cellule delle piante: esse servono a rimescolare il
citoplasma e facilitare la diffusione dei soluti. Il
citoscheletro actina-miosina è coinvolto anche nel
controllo della posizione di alcuni tipi di organelli
cellulari, in particolare i cloroplasti. Questo permette alla
pianta di reagire a diverse intensità di illuminazione.
Distibuzione dei cloroplasti nelle cellule della foglia di
Arabidopsis a bassa (sinistra) o alta (destra) intensità
luminosa. Se la luce è molto intensa i cloroplasti si
dispongono in modo da coprirsi l un l altro e proteggersi
dal rischio di danni fotochimici.
Il citoscheletro microtubulare delle piante presenta una struttura e
una dinamica altamente distintive rispetto agli animali.
I microfilamenti controllano anche la
migrazione delle vescicole di esocitosi.
Il loro ruolo è particolarmente importante
nelle cellule a crescita apicale (tipgrowth), per esempio i peli radicali e i
tubetti pollinici.
Nella cellula animale in interfase il citoscheletro microtubulare si
irradia da una coppia di centrioli situata vicino al nucleo (A). Nelle
cellule delle piante in interfase i microtubuli si irradiano dal nucleo
in più punti e si dispongono a ridosso del plasmalemma, formando
citoscheletro corticale (B).
I microtubuli nella cellula animale si formano a partire da un centro
organizzatore dei microtubuli (MTOC) che è associato ai centrioli.
Nelle cellule delle piante si ritiene che esistano numerosi MTOC,
associati all involucro nucleare e forse al plasmalemma
B
A
Citoscheletro
corticale
10
04/04/17
Nelle varie fasi del ciclo cellulare della cellula delle piante sono
attivi diversi sistemi di citoscheletro microtubulare. In
interfase è presente un citoscheletro corticale di microtubuli
associati al plasmalemma, il cui orientamento influenza quello
delle microfibrille di cellulosa che vengono deposte nella
parete in espansione. All inizio della mitosi (fase G2) questo
sistema scompare ed è sostituito da una banda compatta di
microtubuli (banda preprofasica) la cui posizione individua il
piano nel quale si formerà il setto di divisione (detto anche
piastra cellulare) tra le due cellule figlie.
Nel corso della mitosi compare il fuso mitotico anastrale
(nelle piante non sono presenti centrioli). Come negli animali,
questo complesso citoscheletrico controlla la ripartizione dei
cromatidi fratelli nei due nuclei. In telofase compare un
quarto sistema di microtubuli, il fragmoplasto, che si forma
prima in posizione centrale fra i due nuclei e poi si espande
verso la periferia. Il fragmoplasto controlla la formazione del
setto di divisione (nuova parete cellulare).
Citoscheletro
corticale
Banda
preprofasica
Setto di
divisione
La citodieresi avviene per strozzamento centrale
ad opera di un anello corticale di acto-miosina
nelle cellule animali, mentre coinvolge il
fragmoplasto nelle cellule delle piante
Setto di
divisione
Una volta
completato il
setto, il
fragmoplasto
scompare e
ricompare il
citoscheletro
corticale, che
persiste per
tutta l interfase.
11
 Cellula vegetale")



![LEZ 0 struttura piante e cellula pprima [modalità compatibilità]](http://s1.studylibit.com/store/data/001457124_1-1e8be3d6a5644dc1f8a3a96b155dceca-300x300.png)





