")
Gianmario Gerardi
Funzioni vitali della cellula:
La riproduzione
Le due funzioni vitali della cellula: metabolismo e riproduzione
L’unità funzionale minima di un essere vivente è una cellula viva. Tuttavia, ogni essere
vivente, appare costituito in ultima analisi da niente altro che grandi e complessi insiemi
di reazioni chimiche regolate, che consentono il realizzarsi di due fenomeni chimico-fisici
di rilievo.
Il primo fenomeno consiste nella capacità di ottenere e trasferire energia,
rendendola disponibile a processi non spontanei, che altrimenti non
avverrebbero. Tali processi possono così produrre nuove molecole complesse e ricche di
energia interna e possono garantire il mantenimento di un ambiente chimico-fisico
stabile, separato da quello circostante, che possa perdurare nel tempo, e continuare nella
realizzazione dei suoi intenti. Il complesso insieme di reazioni coinvolte all’interno di
questo fenomeno va sotto il nome di metabolismo cellulare.
Il secondo fenomeno è quello della riproduzione. Esso è in realtà un effetto di
quest’ultima tendenza, appena descritta, a conservare nel tempo tutti i processi utili.
La riproduzione consiste nella replicazione esatta del proprio sistema chimico e
organico complessivo, in un nuovo individuo uguale al precedente, prima che
esso degeneri; ovvero, prima che processi inesorabili e solo in parte arginabili, come
l’ossidazione effettuata dall’ossigeno gassoso a carico delle strutture organiche, abbiano il
sopravvento.
Approfondimenti e Richiami
La freccia del tempo. L’evoluzione chimica dell’Universo, dal punto di vista termodinamico, avviene
complessivamente in un solo senso, plausibilmente originato dagli eventi del Big Bang. La tendenza generale
della materia e dell’energia dell’universo, è di modificarsi spontaneamente, e attraverso passaggi casuali, verso
una diminuzione di entalpia e un aumento di entropia. Detto in termini non specialistici, potremmo dire che
l’Universo si va raffreddando e degradando; ed è plausibile che ciò appaia a chiunque ovvio e naturale, perché
chiunque prima o
poi nella vita
impara cosa
siano un fenomeno
irrimediabilmente
irreversibile o
l’invecchiamento e la morte.
Semplificando, possiamo dire che l’entalpia è data dall’energia cinetica delle particelle e dall’energia potenziale
racchiusa nelle interazioni di legame tra un atomo e l’altro o tra una molecola e l’altra, mentre l’entropia è il
grado di disordine con cui queste particelle sono organizzate all’interno di un sistema. Ad esempio, in un
cristallo di ghiaccio c’è molto più ordine che in una nube di vapor acqueo, ma in quest’ultima, l’energia cinetica
media delle particelle è molto più elevata. Se nell’Universo l’entalpia e l’ordine vanno complessivamente
diminuendo, significa che le trasformazioni chimiche spontanee evolvono verso un maggiore disordine d’insieme
del sistema e verso una diminuzione dell’energia custodita all’interno delle sostanze. L’energia totale
dell’Universo resta costante, perché nulla può andare perduto, ma col passare del tempo l’energia contenuta
nelle molecole si degrada, passando a livelli di energia interna inferiori e a un maggiore grado di disordine.
Questo inesorabile scadimento dell’energia posseduta dalle molecole, rappresenta per l’Universo un vero e
proprio orologio, chiamato anche freccia del tempo.
La vita. Uno degli elementi di maggior distinzione dei sistemi viventi rispetto ai sistemi inanimati è senza
dubbio la loro capacità di riprodursi. Le risposte ultime al perché i processi biologici abbiano questa
2
Gianmario Gerardi
irrinunciabile esigenza a protrarre nel tempo le loro attività chimiche sono tutt’altro che chiare. Tuttavia,
essendo la vita un fenomeno termodinamico caratterizzato dall’assorbimento di energia, anziché dalla sua
liberazione, la riproduzione, in un certo senso, può essere vista come nient’altro che un mezzo, in grado di
evitare che l’energia accumulata se ne vada perduta, trascinata dall’inesorabile degradarsi e raffreddarsi di
tutto l’Universo. La vita, peraltro, non accumula solo energia, ma anche organizzazione e ordine e anche
l’organizzazione e l’ordine, se non vengono periodicamente verificati e rinnovati, finiscono per deteriorarsi e
collassare, esattamente come l’organizzazione di un’azienda o l’ordine di una stanza.
La vita ha una tendenza specifica a mantenere nel tempo i suoi processi, modificandoli, rinnovandoli,
adattandoli costantemente all’ambiente e creando sistemi adibiti alla riparazione di danni ed errori. Si pensi, ad
esempio, ai sistemi di riparazione delle membrane lipidiche o del DNA e a tutti i sistemi antiossidanti di cui le
cellule sono dotate, ma, soprattutto, si pensi appunto al complesso fenomeno della riproduzione.
La riproduzione di qualunque individuo, porta inoltre in sé un effetto secondario non da poco e cioè quello della
moltiplicazione. Un organismo che, molto prima di degenerare, ne genera almeno uno uguale, ha prodotto in
realtà due individui. Nel caso delle cellule dei tessuti animali, di molti eucarioti unicellulari e di tutti i procarioti,
la riproduzione avviene proprio per moltiplicazione e l’individuo originale non muore, ma, dividendosi, ne
genera due uguali e rinnovati. Un effetto, come si è detto, non da poco, perché la crescita numerica dovuta alla
moltiplicazione degli esseri unicellulari, molto probabilmente, ha aperto la strada a nuove strategie utili ai fini
prima descritti, come la cooperazione cellulare e l’organizzazione biologica di tipo coloniale, fino alla
costituzione dei tessuti complessi negli organismi superiori.
In termini termodinamici, la replicazione e la moltiplicazione comportano un aumento complessivo di
entalpia, soprattutto di energia potenziale di legame, e una diminuzione di entropia, realizzando
un’organizzazione del sistema specifica e ordinata.
Al contrario, il bilancio complessivo di quanto accade spontaneamente nell’Universo corrisponde ad una
diminuzione generale di entalpia e ad un aumento di disordine (aumento di entropia). Pertanto, affinché i
processi biologici abbiano luogo, è indispensabile che nei dintorni di un sistema vivente, avvengano variazioni di
entalpia e di disordine più ampie rispetto a quelle previste dal bilancio dell’Universo. In questo modo, il difetto
dovuto all’aumento di entalpia e alla diminuzione di entropia realizzate nel sistema vivente, viene colmato in
modo che il bilancio globale possa restare costante.
La riproduzione cellulare
L’invecchiamento e il ciclo cellulare
Ossidazione, invecchiamento ed errore. Gli effetti degradativi più rapidi e violenti a
carico degli esseri viventi sono quelli prodotti dall’attività ossidante dell’ossigeno, che
è presente in grande quantità nell’atmosfera terrestre. Data la sua elettronegatività (3,5
nella scala di Pauling), il suo potere ossidante è particolarmente vigoroso sulle molecole
biologiche, che, generalmente, sono molecole ridotte e quindi ricche di elettroni. Quasi
tutti gli esseri viventi sfruttano il potere ossidante dell’ossigeno per ricavarne energia: si
è tuttavia abbastanza certi che non sia sempre stato così e, anzi, è abbastanza probabile
che inizialmente la comparsa dell’ossigeno nell’aria in quantità significative, abbia creato
non pochi problemi.
Resta il fatto che tutti gli organismi, chi più chi meno, hanno una forte tendenza ad
invecchiare, a causa, soprattutto, dell’azione ossidante dell’ossigeno che, per quanto
venga arginata il più possibile, risulta prima o poi fatale.
Se anche non vi fosse questo grosso problema dell’ossigeno, che, del resto, come si
diceva è un problema insorto successivamente, l’organizzazione realizzata dai sistemi
biologici sarebbe comunque soggetta all’accumulo di errori e di danni di altro genere,
dovuti al livello di complessità del sistema.
3
Gianmario Gerardi
Approfondimenti e Richiami
Per la verità, secondo i fisici, la maggior parte di questi errori sono l’effetto del bombardamento quotidiano di
particelle provenienti dal cosmo, chiamate muoni o raggi cosmici a cui la superficie terrestre è sottoposta.
Ciò depone a favore di una qualità organizzativa del sistema vivente decisamente migliore di quanto possa
apparire ai biologi. Resta il fatto che una quota di questi errori è inevitabile e alcuni di essi possono risultare
anche fatali.
Quando questi errori riguardano direttamente il materiale genetico dell’organismo,
possono generare in esso cambiamenti irreversibili chiamati mutazioni. Esistono
meccanismi biologici di riparazione in grado di prevenire mutazioni genetiche, ma,
quando questi falliscono o si dimostrano inefficaci, la mutazione permane e si perpetua
lungo le generazioni successive (eccetto il caso in cui la mutazione non consenta
all’individuo di riprodursi).
È fondamentale ricordare che le mutazioni, talvolta, si manifestano come caratteristiche
organiche di un individuo (caratteri fenotipici) e che, perciò, sono alla base della
variabilità tra gli esseri viventi. Le mutazioni, quindi, hanno contribuito profondamente
a dare origine alla enorme diversità esistente tra gli organismi favorendo il realizzarsi
della selezione naturale di alcuni individui rispetto ad altri.
A causa dell’inevitabile degradazione ossidativa delle macromolecole biologiche e
dell’accumularsi di questi errori, le strutture di tutti gli organismi viventi vengono
costantemente rinnovate e ciascun organismo, sia che si tratti di una singola cellula o di
un organismo superiore, è in grado di produrre una copia rinnovata di sé, “un giovane
figlio.”
Il ciclo vitale di ogni organismo, quindi, prevede una fase di replicazione di se stesso,
al termine della quale l’organismo originale può anche morire. Normalmente la prima
fase di vita del nuovo individuo replicato è una fase di crescita e sviluppo, alla quale
segue quindi la riproduzione e ad un certo punto la morte.
Il ciclo cellulare. Ogni singola cellula eucariote suddivide la sua vita in 2 fasi
abbastanza distinte che prendono il nome di interfase e di divisione o mitosi (M).
La divisione è la fase della riproduzione in cui la cellula, dividendosi in due, darà origine a
due cellule uguali a quella originale, ma, in un certo senso, ringiovanite nelle loro
strutture. L’interfase, invece, è il periodo che intercorre tra una divisione e la successiva
ed è a sua volta divisa in tre sottofasi successive chiamate G1, S e G2 (figura 1).
G1 e G2 sono due fasi di crescita della cellula: mentre in G1 vengono svolte le normali
attività metaboliche specifiche, in G2 la cellula si prepara alla divisione. La fase S, situata
temporalmente tra la fase G1 e la fase G2, è un lungo periodo di sintesi molto attiva di
DNA.
Prima che la cellula si divida, infatti, tutto il DNA contenuto nel nucleo deve essere
riprodotto in una copia esattamente uguale. Le due copie uguali verranno destinate alle
due nuove cellule che avranno origine dalla divisione. I tempi di ciascuna fase del ciclo
vitale di una cellula sono molto diversi in base al ruolo che questa cellula svolge.
Nelle cellule germinative dello strato basale dell’epidermide, ad esempio, le cellule sono
in attiva divisione, con mitosi continue e con una fase metabolica G1 non estesa
(pochissimi giorni). Gli epatociti del fegato, invece, hanno un numero di divisioni inferiori
e una vita metabolica prolungata fino a più di 10 giorni, tale da consentire loro di
svolgere le funzioni epatiche per cui sono preposti.
4
Gianmario Gerardi
G2
M
Cromosomi
spiralizzati
(eucromatina)
S
G1
Figura 1 - Il ciclo cellulare. La vita di una cellula eucariote è suddivisa in 2 fasi chiamate interfase (in
azzurro) e mitosi o divisione (in giallo). L’interfase è costituita da tre sottofasi successive chiamate G1, S e G2.
Nella fase G1 la cellula cresce e svolge le sue normali attività metaboliche. La fase S consiste in un periodo nel
quale avviene una sintesi attiva di DNA, la replicazione del corredo cromosomico in vista della divisione. Nella
fase G2 la cellula si prepara in tutto e per tutto alla divisione. Lungo il periodo di divisione, la fase M, si
susseguono cinque sottofasi distinte: profase, metafase, anafase, telofase e citodieresi, ovvero la divisione vera
e propria, durante le quali il DNA è fortemente compattato a formare i cromosomi che appaiono distinguibili.
(Tobin, Dusheck – Bios2 Genetica ed Evoluzione; Mondadori)
Le cellule entrano in fase S solo quando destinate ad andare incontro alla divisione. I
neuroni, ad esempio, non si dividono mai e restano in una fase cosiddetta G0 per tutta la
loro vita. La divisione cellulare è un processo riproduttivo basato sulla moltiplicazione,
dove l’individuo originale non muore ma si moltiplica; pertanto la sua attivazione è
regolata da molti fattori, interni ed esterni, in grado di indicare quale estensione debba
raggiungere la popolazione di quel tipo di cellule. I tumori sono malattie in cui la quota di
divisione cellulare non è più regolata e il tessuto specifico comincia ad accrescersi senza
limiti per moltiplicazione cellulare. Normalmente uno dei fattori in grado di rallentare
sensibilmente i processi di divisione e il numero medio di cellule di un tessuto che
entrano in fase S e G2 è il contatto tra le cellule stesse.
L’organizzazione del DNA
Il codice delle informazioni. Il DNA, l’acido desossiribonucleico, dal punto di vista
funzionale è un vero e proprio archivio in codice, contenente l’informazione completa,
necessaria alla costruzione e regolazione di una cellula o anche di un intero organismo
pluricellulare, come potrebbe essere un rettile, un mammifero o una pianta. Ogni cellula
di uno stesso organismo contiene tutta l’informazione che riguarda l’intero organismo,
5
Gianmario Gerardi
compresa, in linea teorica, anche la sua completa ricostruzione da zero; l’unica differenza
sta nelle porzioni di DNA che, a seconda del ruolo svolto dalla singola cellula, vengono
mantenute attive oppure in stato di archivio inattivo. L’insieme di tutta questa
informazione viene chiamata patrimonio genetico.
L’archivio del DNA viene costantemente consultato durante tutta la vita della cellula
tramite il processo della trascrizione e viene copiato e completamente riprodotto prima
della divisione cellulare tramite il processo della replicazione.
È bene ricordare che il DNA non è altro che una serie di lunghi filamenti a doppia elica
complementare, ciascuna chimicamente costituita dalla polimerizzazione di 4 molecole
chiamate nucleotidi (adenina, guanina, citosina e timina) poste una dopo l’altra in
alternanza non regolare e con ripetizioni, di cui qui non parleremo. La sequenza specifica,
con cui si susseguono i 4 tipi di nucleotidi, genera dei tratti distinti di DNA chiamati geni,
costituiti ognuno dalla successione di centinaia o migliaia di questi nucleotidi.
Come mostrato in figura 2 ad ogni gene corrisponde funzionalmente la sequenza, e
quindi la struttura e la funzione, di una proteina specifica o, in seguito a
rimaneggiamenti dell’espressione genica, di alcune varianti della stessa proteina. Il
DNA risulta essere pertanto l’archivio della sequenza di tutte le proteine di un organismo,
più alcuni tratti con funzione regolativa del DNA stesso.
Figura 2 – Principio di “gene Æ proteina specifica”. Il DNA di ogni cromosoma è una sequenza di geni,
costituiti da sequenze di nucleotidi. Ad ogni gene corrisponde la sequenza di una sola proteina o di alcune sue
varianti specifiche. La proteina può svolgere la sua funzione specifica per cui è preposta solo se ha raggiunto la
sua conformazione tridimensionale corretta. (La proteina mostrata in figura esiste realmente, ma nel suo
complesso questa figura è del tutto immaginaria).
Il numero di questi filamenti, all’interno del nucleo delle cellule, è piccolo ma
estremamente preciso e specifico per ogni organismo. Nell’uomo, in qualunque sua
6
Gianmario Gerardi
cellula, i filamenti di DNA sono 46, costituiti in realtà da 23 coppie di filamenti, in cui
ciascuna coppia è formata da due filamenti separati e autonomi, la cui sequenza di geni,
però, è esattamente la stessa; mentre la sequenza di nucleotidi di uno stesso gene
presenta delle variazioni. Ciascun filamento di DNA costituisce un cromosoma.
La fase di trascrizione, citata sopra come fase di consultazione dell’archivio, consiste
nella sintesi chimica di una copia esatta di un gene, sottoforma però di RNA a singola
elica (una singola elica di acido ribonucleico nella quale ogni timina è sostituita da un
nucleotide simile, detto uracile). Affinché questo tratto di RNA si trasformi in una
proteina è necessario che avvenga una fase di traduzione, basata appunto su un codice,
il codice genetico.
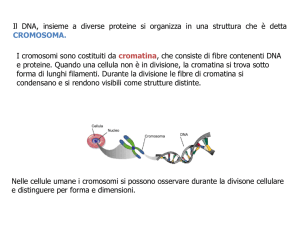
Il cromosoma e la replicazione del patrimonio genetico. Ognuno dei filamenti di
DNA presenti nel nucleo è in grado di organizzare un livello strutturale di
superavvolgimento per spiralizzazione e compattazione, tale da ridurne la lunghezza
complessiva di milioni di volte e assumendo la forma di un bastoncello, strozzato più o
meno al centro. Come mostrato in figura 3, i gradi di spiralizzazione sono
complessivamente 4. Il primo avviene per avvolgimento attorno a ottameri di proteine
chiamate istoni, il secondo è una struttura a spirale vera e propria del filamento
risultante, il terzo è un compattamento della spirale a ripetizione di “anse ad S” e il
quarto è un raggomitolamento disordinato. In questo stadio strutturale il DNA prende il
nome specifico di cromosoma ed esso si rende visibile al microscopio ottico durante il
periodo di divisione della cellula. Nelle fasi di divisione delle cellule dell’uomo, il DNA
compare nel nucleo organizzato e distinguibile in 46 cromosomi.
Nel periodo cellulare di interfase, invece, il DNA di ogni cromosoma è indistinguibile, in
quanto molti dei suoi tratti sono despiralizzati ed estesi, affinché possa essere possibile la
loro consultazione ovvero la loro trascrizione e, nella fase S, si possa anche svolgere la
replicazione. In questa fase, l’intera massa di DNA, prende il nome di cromatina. Più
specificamente il DNA nel suo stato parzialmente svolto e accessibile viene definito
organizzato in eucromatina; il DNA in forma di “fibra” condensata in cui gli istoni sono
completamente aggregati e inaccessibili, viene invece definito eterocromatina.
eucromatina
eterocromatina
Cromosoma condensato
Figura 3 – Gradi di spiralizzazione del DNA. Tra la forma meno addensata e l’addensamento massimo di un
cromosoma vi è un rapporto dimensionale di almeno 1:1000. (Alberts B. et al - Molecular Biology of the Cell,
ed. it. Zanichelli, 1994)
7
Gianmario Gerardi
Come accennato poco fa, nella sottofase S, quella cosiddetta di sintesi del DNA che
occupa più o meno 1/3 del ciclo cellulare, la cromatina inizia anche la replicazione di tutto
il DNA presente, ovvero di tutto il patrimonio genetico posseduto. La cellula trascorre
quindi un terzo della sua vita ad eseguire la copia integrale di tutto il suo archivio
d’informazioni al fine di tramandarlo alle cellule figlie.
La replicazione del DNA, per sommi capi, si svolge attraverso la separazione delle due
eliche di un filamento e la costruzione di due nuove eliche, complementari alle due
originali. Al termine ogni filamento avrà dato origine a due filamenti uguali, ciascuno
composto da un’elica originaria e un’elica complementare di nuova sintesi (figura 4).
B
A
C
D
E
Figura 4 – Replicazione del DNA. Durante la fase S del ciclo cellulare il DNA (A) viene completamente
duplicato. La duplicazione avviene grazie alla separazione delle eliche complementari in vari punti di ogni
filamento (B) e alla sintesi di due nuove eliche complementari per ciascuna di quelle preesistenti (C). Al termine
ogni filamento di DNA sarà presente in 2 copie uguali formate da un’elica preesistente e da un elica
complementare di nuova sintesi (D). Nei procarioti (E) il filamento di DNA è circolare e vi è un’unica origine di
replicazione che si espande via via lungo tutto il cromosoma fino allo sdoppiamento completo.
(Curtis, Barnes – Invito alla Biologia; Zanichelli)
Negli organismi procarioti, privi di nucleo, il cromosoma è decisamente più breve rispetto
agli eucarioti e generalmente ha forma circolare chiusa, cioè il filamento di doppia
elica è richiuso ad anello. Anche in questo caso esso può variare il suo livello di
superavvolgimento, anche se in termini molto più semplici. Nei batteri la massa attiva di
DNA viene talvolta chiamata nucleoide.
8
Gianmario Gerardi
Il gene, la ploidia, i cromosomi omologhi e gli alleli. Nell’uomo, e in moltissimi altri
organismi, nel nucleo delle cellule ogni tipo di cromosoma, con la sua specifica sequenza
di geni, è presente in doppia copia; vale a dire che anche ogni gene è presente in doppia
copia. Questo corredo multiplo viene descritto con il termine di ploidia e indicato con la
lettera n minuscolo: organismi in cui ogni cromosoma e ogni gene è presente in unica
copia sono detti aploidi (n), mentre quando sono presenti più copie vengono chiamati
poliploidi (nn). L’uomo, i mammiferi e moltissimi altri animali superiori sono organismi
diploidi (2n), con due copie per ciascun gene o cromosoma.
I cromosomi che presentano la stessa sequenza di geni sono detti cromosomi
omologhi, le cellule umane contengono quindi 23 coppie di cromosomi omologhi. Su
ciascun cromosoma omologo sono presenti, nella stessa esatta posizione, gli stessi geni, i
quali, però non è detto che abbiano la stessa esatta sequenza di nucleotidi; anzi,
generalmente, non è così e presentano invece piccole variazioni.
L’attività genica di organismi diploidi come i mammiferi, in termini di funzione proteica
corrispondente, è quindi determinata dall’azione di due fattori, cioè di due geni
corrispondenti alla stessa proteina, ma con sequenze leggermente diverse. Questi due
fattori sono rappresentati dai geni omologhi e sono detti alleli.
Approfondimenti e Richiami
Genotipo e fenotipo. Il fatto che i geni omologhi, cioè gli alleli, possano anche avere sequenze diverse
comporta che un gene possa corrispondere ad una proteina più efficiente rispetto a quella corrispondente
all’altro allele. Spesso ciò fa sì che gli effetti di un gene mascherino addirittura quelli dell’altro allele. In questo
caso si dice che un allele è dominante mentre l’altro è recessivo; per uno stesso gene possono quindi esistere
due copie di alleli uguali e dominanti (omozigoti) oppure due copie diverse in cui uno è dominante e l’altro
recessivo (eterozigoti) o, ancora, due copie uguali ma recessive rispetto a geni uguali ma dominanti, presenti
nella stessa popolazione di individui della stessa specie (omozigoti recessivi). La manifestazione dell’attività di
una proteina, chiamata carattere somatico, è quindi la risultante dell’attività di due alleli. Tale risultante visibile,
viene definita fenotipo, mentre la composizione specifica del gene nei suoi due alleli è detta genotipo e non è
sempre manifesta.
Il cromosoma artificiale. Articoli comparsi su due pagine monotematiche in Corriere
della sera – 07 ott. 2007. L’articolo presenta, senza in verità spiegare granché, i risultati
dell’ultima ricerca di Craig Venter (che nel 2000 ha consegnato la mappatura finale del
genoma umano) il quale per la prima volta avrebbe dato alla luce un cromosoma
completo, anche se minimo, del tutto artificiale. Più precisamente si tratterebbe di una
versione minimale del cromosoma di Mycoplasma genitalium. Secondo Venter e altri
commentatori il traguardo è significativo e dimostra quanto si sia vicini alla realizzazione
della vita artificiale. Il prossimo passo sarebbe l’inserimento del cromosoma in un
organismo privato del suo DNA (vedi Allegato 1).
Sequenza del film “BLADE RUNNER”. A conclusione della discussione sull’articolo del
Corriere della sera è pertinente inoltrarsi nella provocazione con la visione di una o due
sequenze del film di Ridley Scott Blade Runner. Il film di fantascienza, ottimo in termini
cinematografici, narra di un poliziotto speciale incaricato di dare la caccia a persone
“replicanti”, ottenute in laboratorio con tecniche di ingegneria genetica, impiegate come
soldati in guerre interstellari e successivamente sfuggite al controllo umano. Il film,
attraverso la rappresentazione della vita artificiale, invita alla riflessione sui temi della
vita intesa in senso esclusivamente meccanicistico, ponendo interrogativi sul concetto di
coscienza. Nelle due sequenze il replicante più forte ed evoluto decide di eliminare il suo
creatore, umano, e al termine di un duello finale con il poliziotto (interpretato da Herrison
Ford) recita sotto la pioggia la famosa frase: “Io ne ho viste di cose, che voi umani…
ecc.”
9
Gianmario Gerardi
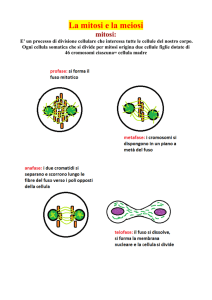
La riproduzione di una cellula: la divisione
La Mitosi. Come accennato parlando del ciclo vitale di una cellula, le cellule giungono ad
una fase di attiva preparazione alla divisione, che terminerà con la formazione di due
cellule uguali a quella di partenza. Questa fase viene complessivamente definita mitosi
(fase M del ciclo) e può occupare a grandi linee anche meno di 1/30 dell’intera durata di
vita di una cellula.
Essa si compone di 5 fasi abbastanza distinte, poste però come una serie ininterrotta di
eventi.
La prima fase viene definita profase (figura 5) ed è uno stadio nel quale un microscopio
ottico consente di cominciare a distinguere un cromosoma dall’altro, anche se il
compattamento non si è ancora del tutto concluso. I cromosomi non appaiono però come
dei bastoncelli, ma come coppie di bastoncelli paralleli, unite in prossimità della loro
strozzatura, chiamata centromero. La sintesi di DNA operata dalla cellula, durante la
sua fase S, ha infatti prodotto una copia identica per ogni cromosoma e queste due copie
si presentano come un unico cromosoma della forma semplificata e ben nota di X, dove
le braccia di un lato della X e quelle dell’altro lato, corrispondono alla copia esatta dello
stesso cromosoma originale e prendono il nome di cromatidi fratelli. Un cromosoma
profasico è quindi una struttura composta da 2 cromatidi fratelli uguali.
Inoltre è evidente che l’organizzazione citoscheletrica della cellula sta cambiando
profondamente. Da ciascuno dei due centrioli, infatti, che lentamente stanno migrando
ai poli opposti della cellula, iniziano ad allungarsi dei microtubuli, che si orienteranno tutti
ad ombrello in direzione del piano dell’equatore della cellula, quelli di un centriolo opposti
a quelli dell’altro, andando a formare una struttura detta anche “a pallone da rugby”
chiamata fuso mitotico (figura 6). I due centrioli attivi prendono il nome di
centrosomi. La doppia membrana nucleare inizia a rompersi e dissolversi.
A
B
C
Figura 5 – Inizio della mitosi. Dall’interfase (A), le cellule entrano improvvisamente in una sequenza di
notevoli mutamenti. Nella profase (B) i cromosomi cominciano ad essere
distinguibili mentre la membrana
nucleare si sfalda e va scomparendo. Durante l’anafase i cromosomi terminano il loro compattamento e
migrano verso il piano equatoriale della cellula, dove vengono connessi ai microtubuli del fuso mitotico (C).
(Ricostruzione grafica: Curtis, Barnes – Invito alla Biologia; Zanichelli)
10
Gianmario Gerardi
Figura 6 – Fuso mitotico e centrosoma. Durante la profase le due coppie di centrioli presenti nella cellula si
spostano e migrano ognuna verso i poli opposti della cellula. Durante la migrazione, intorno a ogni coppia di
centrioli si rende visibile un’area di attività chiamata centrosoma, dalla quale cominciano ad accrescersi lunghe
fibre di microtubuli in direzione del piano equatoriale della cellula con una forma raggiata ad ombrello, fino a
formare una struttura complessiva a fuso.
(Ricostruzione grafica da materiale web Dip. Biologia Università di Pavia)
Alla profase segue la prometafase (figura 7): la forma dei cromosomi, composti dai due
cromatidi fratelli, si fa ancora più compatta ed evidente; la membrana nucleare è
scomparsa, il fuso mitotico si è ulteriormente esteso nel volume cellulare e i centromeri
di ogni cromosoma iniziano a legarsi alle estremità dei tubuli del fuso mitotico poste in
prossimità del piano dell’equatore della cellula.
Quindi sopraggiunge la metafase: l’attribuzione di questo stadio della divisione cellulare
viene fatta quando i cromosomi sono migrati tutti presso il piano equatoriale della cellula,
agganciati dai microtubuli dei centrosomi ai cinetocori, strutture specifiche di
ancoraggio dei centromeri cromosomici (figura 8). Ad ogni cromosoma sono agganciate,
presso i cinetocori, una fibra proveniente da un polo della cellula e una fibra proveniente
dal centrosoma del polo opposto. I cromatidi fratelli di ciascun cromosoma si orientano
coricati e paralleli al piano equatoriale.
Nella fase successiva, che prende il nome di anafase (figura 9), i cromatidi sono stati
separati dai loro rispettivi fratelli, tramite l’attività di contrazione dei microtubuli delle
fibre del fuso mitotico e da una sorta di attività di risalita sulla fibra del fuso, compiuta
dal cinetocoro stesso. In questa fase i cromatidi sono cromosomi a tutti gli effetti. Verso i
poli opposti della cellula migrano dunque copie uguali dell’intero patrimonio genetico
della cellula. Non tutte le fibre del fuso sono agganciate ai cromosomi; alcune sembrano
saldate a quelle opposte e allungandosi forzano la cellula a distendersi e ingrandirsi.
11
Gianmario Gerardi
Le fasi conclusive della divisione sono la telofase e la citodieresi vera e propria (figura
9). La telofase può essere interpretata come il corrispettivo della profase ma in direzione
costruttiva anziché demolitiva. I cromosomi, sempre più raccolti nelle due aree
circoscritte dei poli della cellula originale, iniziano leggermente a despiralizzarsi e intorno
ad ognuno di essi cominciano a comparire tratti di membrana nucleare.
Durante la citodieresi avviene la vera e definitiva divisione della cellula in due cellule
figlie; nella membrana esterna della cellula compare un solco, un setto di divisione
all’altezza dell’equatore che continuerà a stringersi fino alla divisione completa della
cellula; anche al piano dell’equatore, compaiono tratti di membrana a formare un setto di
separazione (figura 10).
A
B
C
Figura 7 – Seconda parte della mitosi. A metafase ultimata i cromosomi sono allineati sul piano
dell’equatore della cellula con le zone centromeriche rivolte alle fibre del fuso (A). Durante l’anafase i cromatidi
fratelli di ciascun cromosoma vengono separati e iniziano la migrazione verso i poli della cellula per attività
contrattile delle fibre del fuso e dei cinetocori (B). La telofase è l’inizio della divisione vera e propria: la cellula si
strozza all’equatore e attorno ai cromosomi separati ai due estremi della cellula si iniziano a formare due nuovi
nuclei; le fibre del fuso vanno scomparendo (C).
(Ricostruzione grafica: Curtis, Barnes – Invito alla Biologia; Zanichelli)
12
Gianmario Gerardi
Figura 8 – Separazione dei cromatidi fratelli in anafase. A metafase ultimata i cromosomi sono allineati
sul piano dell’equatore della cellula con le zone centromeriche aderenti al fuso mitotico. I cinetocori, infatti sono
attaccati alle fibre del fuso (A-B). Durante l’anafase (C), i cromatidi fratelli di ciascun cromosoma vengono
separati e iniziano la migrazione verso i poli della cellula. Il movimento è prodotto dall’attività contrattile delle
fibre del fuso e dei cinetocori (A).
(Ricostruzione grafica da materiale web assortito)
Figura 10 – Citodieresi. Citodieresi di una cellula animale con formazione del solco di scissione osservata al
microscopio elettronico a scansione.
(Campbell, Reece, Taylor, Simon – Immagini della Biologia (A) La cellula; Zanichelli)
13
Gianmario Gerardi
Altri modelli di divisione cellulare. Nelle cellule vegetali il processo di citodieresi è
diverso a causa della presenza della parete cellulare esterna che è rigida. Nella cellula
vegetale non compaiono i centrosomi e la divisione avviene dopo che al centro si è
formata una piastra di separazione, costituita da un deposito di polisaccaridi rilasciati
da vescicole provenienti dall’apparato di Golgi e da un accumulo di pectina. La piastra di
separazione matura diventerà la lamella mediana frapposta fra le due nuove cellule, le
quali inizieranno autonomamente a costruire la membrana fosfolipidica e a depositare lo
strato primario della parete. A quest’ultimo, col tempo, viene successivamente
sovrapposto il deposito secondario più spesso, inserito tra la parete primaria a la
membrana lipidica (figura 11).
Nei batteri, invece, la divisione avviene per scissione binaria, un processo che
prevede che la cellula, prima di dividersi, si sia allungata pari a circa due volte la
lunghezza iniziale del microrganismo. Nel frattempo, il DNA cromosomico circolare si è
completamente duplicato e le due nuove copie hanno ancorato un breve tratto alla
membrana dei due poli opposti di divisione. Dividendosi, la cellula, separa perfettamente
le due copie uguali di cromosoma (già mostrato in figura 4 E).
lamella
mediana
Figura 11 – Citodieresi nella cellula vegetale. La divisione avviene in seguito alla deposizione in zona
mediana della piastra di separazione, costituita da polisaccaridi e da un accumulo di pectina. La piastra di
separazione matura diventa la lamella mediana frapposta fra le due nuove cellule, nelle quali inizia la
costruzione della membrana fosfolipidica e il deposito dello strato primario della parete. A quest’ultimo viene
successivamente sovrapposto il deposito secondario più spesso, inserito tra la parete primaria a la membrana
lipidica.
(Tobin, Dusheck – Bios2 Genetica ed Evoluzione; Mondadori)
Contenuti sull’esperienza di laboratorio
Cellule mitotiche in apici radicali di cipolla. È possibile, con una dotazione minima di
strumenti di laboratorio, osservare il complesso fenomeno della mitosi nelle sue fasi
fondamentali. Ciò è reso possibile da un facile allestimento appropriato di cellule che si
trovano negli apici radicali di cipolla, a livello dei quali vi è un’intensa attività di
proliferazione cellulare.
14
Gianmario Gerardi
La cipolla è un bulbo, cioè un fusto molto corto e trasformato in un’unica gemma che
cresce sottoterra, sul quale le foglie sono trasformate in tuniche carnose.
La radice si sviluppa senza che prevalga una radice principale, che viene sostituita da una
quantità di radici che spuntano dalla base del fusto, avvolte da un tessuto chiamato cuffia
avente il compito di proteggere i meristemi apicali esposti a ruvidi contatti, man mano
che la radice si allunga nel terreno.
L'allungamento della radice non è dovuto ad un reale spostamento delle sue cellule, ma
alla produzione di nuove cellule che continuamente si aggiungono in posizione apicale.
Il tessuto che svolge il compito di produrre nuove cellule è il tessuto meristematico, che
si trova all'interno della cuffia, da essa strettamente avvolto e protetto.
Le cellule del tessuto meristematico sono cellule piccole ed appiattite, in cui il nucleo è
ancora ben evidente e facilmente colorabile; sono cellule "giovani", non ancora
differenziate che mantengono la capacità di dividersi per tutta la vita della cellula e che
quindi non hanno ancora assunto le caratteristiche morfologiche che le permetteranno di
svolgere un determinato compito. Se queste cellule hanno conservato la capacità di
dividersi, per dare origine a cellule identiche, non è però detto che i processi di divisione
si susseguano incessantemente, essi infatti subiscono l'influenza delle condizioni
ambientali (temperatura, disponibilità di alimenti) e stagionali (ore di luce). I meristemi
apicali sono attivi soprattutto in primavera mentre in autunno e in inverno sospendono la
loro attività per riprenderla la primavera successiva.
Tali cellule, per queste caratteristiche, differiscono quindi dalle cellule vegetali "adulte"
che in molti casi sono ormai incapaci di dividersi e sono caratterizzate dalla presenza al
loro interno di un grande vacuolo, contenente acqua e sostanze di riserva, che occupa
anche più del 90% del volume cellulare e che schiaccia il nucleo lungo la parete cellulare
rendendolo quindi difficile da osservare.
Nonostante, come detto in precedenza, la cellula vegetale svolga la sua divisione in modo
piuttosto diverso dalle cellule animali, in realtà le cellule degli apici radicali sono costituite
da una parete meno ispessita che può anche essere eliminata dalla procedura
sperimentale e, quindi, possono fornire un’immagine delle fasi mitotiche paragonabile a
quella di cellule animali. Inoltre, l’osservazione delle cellule ordinate a “mattone” dalla
presenza delle pareti, consente anche di elaborare concetti riguardanti il ciclo cellulare,
che però qui non sono stati fatti. Per la procedura si veda il protocollo (Allegato 2).
La riproduzione sessuata degli organismi
Il sesso. Dal punto di vista biologico, il sesso è un differenziamento a livello di organi di
uno stesso individuo oppure a livello di individui appartenenti alla stessa specie, a seguito
del quale questi organi o individui differenziati assumono un’identità cosiddetta di
“genere” che può essere di tipo maschile o femminile. Il differenziamento di genere
consente di realizzare il processo di riproduzione tramite il concorso non di un solo
individuo ma di due (riproduzione sessuata). Il sesso è un potente mezzo di
miglioramento dei caratteri di cui si è portatori, che agisce offrendo la possibilità di
aumentare sensibilmente la velocità e la severità dei processi di selezione naturale. Per
fare un parallelo sportivo, sarebbe come se, per migliorare le doti complessive di una
squadra di calcio, si provassero a caso diverse combinazioni di giocatori più o meno
bravi, arrivando rapidamente alla selezione della compagine vincente. Se ogni giocatore
venisse studiato e selezionato singolarmente, il processo sarebbe molto più lento.
La riproduzione sessuata è basata in sostanza su un energico rimescolamento di caratteri
che avviene, prima rimescolando i propri e poi effettuando una fusione di caratteri con
quelli del partner di sesso opposto, il quale si è anch’esso preoccupato di rimescolare i
suoi. Il prodotto di queste mescolanze è un figlio che non assomiglia ai due genitori e che
si espone alla forza selettiva della natura con un “formato” nuovo, la cui efficacia è del
tutto ignota e casuale.
Si tratta quindi di un sistema che può dare origine a figli “deboli” ma che può
casualmente dare origine anche a figli più “forti” o abili. La natura opererà la sua
impietosa selezione premiando i figli più adatti all’ambiente e consentendo a quel tipo di
15
Gianmario Gerardi
individui (cioè a quella “specie”) di fare un certo salto di qualità nella collezione dei suoi
caratteri “utili” alla sopravvivenza e alla riproduzione. Si tenga presente, tra l’altro, che il
figlio è il frutto di due individui e non di uno solo e quindi non proviene da un solo
processo di selezione ma da due, il che raddoppia le sue probabilità di essere il più
adatto.
Approfondimenti e Richiami
Il modo in cui Darwin rilesse ogni manifestazione di variabilità dei caratteri degli esseri viventi, sia tra i diversi
ordini e le diverse specie di organismi che all’interno di una stessa specie o di una famiglia di animali, fu di
grande impatto per tutta la comunità scientifica e per l’opinione pubblica. Le sue ipotesi molto razionali
risultarono subito molto convincenti, come se esse risiedessero già in forma quiescente fra le idee di molti
ricercatori e pensatori. Darwin ebbe senza dubbio il merito di proporre una grande integrazione di frammenti di
ipotesi e sospetti, dandogli voce con estrema lucidità e razionalità; senza farsi minimamente influenzare
dall’effetto che queste idee avrebbero potuto produrre sulla società nel suo insieme.
In effetti la società rimase tutt’altro che indifferente ad ogni suo livello e presero piede anche molte nuove
piccole teorie, diramate da quella generale sull’origine delle specie. Alcune di esse, che vedevano anche nei
processi sociali di distinzione di classe e di emarginazione una forma di selezione naturale, coltivavano l’idea
che l’uomo potesse attivamente prendervi parte e che l’umanità potesse accelerare il miglioramento della sua
specie attraverso la discriminazione cosciente di intere popolazioni. Gli effetti furono, come si sa nel caso ad
esempio del nazismo, anche di una drammaticità inaudita.
Ancora oggi, del resto, Alex Gibney, amministratore delegato della Enron, la famosa grande società energetica
americana, crollata dopo un gigantesco collasso finanziario, ha dichiarato di essere animato da intenzioni
razionali del tutto scientifiche di carattere biologico. L’amministratore della Enron ha dichiarato davanti ai
giudici e all’opinione pubblica che il suo comportamento dissoluto, con cui ha frodato milioni di cittadini
americani e ha provocato gravi incendi nello stato del Messico tramite black-out programmati di energia, era
giustificato dalle grandi novità scientifiche contenute nei lavori di Richard Dawkins sull’evoluzionismo;
soprattutto la sua famosa opera Il gene egoista. È certo, però, che né Darwin né Dawkins avessero qualcosa a
che vedere con questo tipo di conclusioni.
Riporto, anche se di non facilissima lettura, un brano tratto da L’origine dell’uomo di Charles Darwin sulla
selezione sessuale.
“Al contrario ci sono molte altre strutture e istinti che sono stati certamente sviluppati dalla selezione
sessuale, come le armi per l’attacco e i mezzi di difesa di cui si servono i maschi per lottare contro altri maschi
e per eliminare i loro rivali; il loro coraggio e la loro combattività; i loro ornamenti di ogni genere; i loro
organi per produrre della musica vocale o strumentale; e le loro ghiandole che emettono degli odori; la
maggior parte di queste ultime strutture servono solo ad attirare o eccitare le femmine; è evidente che questi
caratteri sono il risultato della selezione sessuale e non della selezione corrente, poiché maschi non armati, non
ornati e non attraenti riuscirebbero altrettanto bene nella lotta per l’esistenza e nella produzione di numerosi
discendenti se non esistessero altri maschi più dotati di loro… C. Darwin – L’origine dell’uomo…, 1871.
I gameti. Affinché dalla fusione dei patrimoni genetici di due individui nasca un nuovo
individuo, dotato di un unico e corretto corredo genetico, è indispensabile che il DNA di
ogni individuo possa essere dimezzato prima della fusione. In effetti, negli organismi
diploidi (2n) ciascun cromosoma è presente in due copie, che abbiamo detto chiamarsi
omologhe, non perfettamente uguali ma con identica sequenza di geni. Per questa
ragione, la modalità utilizzata per il dimezzamento del patrimonio genetico negli esseri
viventi con riproduzione sessuata consiste nella divisione di una cellula genitore in due
cellule aploidi (n), nelle quali è presente una sola copia per ciascuno dei cromosomi del
patrimonio. Queste cellule, destinate a fondersi con cellule aploidi di uguale natura,
provenienti da un altro individuo vengono chiamate gameti.
Negli esseri viventi in cui il sesso ha prodotto anche una profonda differenziazione
morfologica, cioè ha prodotto individui distinti di sesso maschile e di sesso femminile,
queste cellule sono chiamate gameti maschili e gameti femminili e, in base alla loro
forma, hanno acquisito nomi propri. Alcuni esempi sono lo spermatozoo e l’ovulo, i
16
Gianmario Gerardi
gameti rispettivamente maschile e femminile presenti nei vertebrati e in molti altri
animali, oppure il polline, gamete maschile dei fiori delle piante. Per maggior precisione,
anche in assenza di differenziazione sessuale, alcuni organismi sono in grado di produrre,
anche da uno stesso individuo gameti di sesso femminile e gameti di sesso maschile
(ermafroditismo).
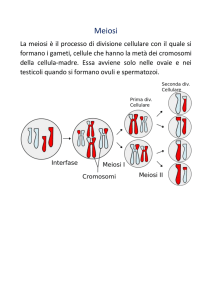
La gametogenesi e la meiosi. La formazione dei gameti spetta a cellule specializzate
dette della linea germinale. Nel loro processo di differenziamento è compresa una fase
in cui ogni cellula germinale andrà incontro a due divisioni successive. Tale processo è
chiamato meiosi, al termine del quale ogni cellula germinale avrà dato 4 gameti aploidi
diversi. Il processo meiotico, infatti, oltre a produrre cellule aploidi con corredo dimezzato
esegue dei processi destinati a differenziare ciascun gamete dall’altro, in modo che ve ne
possano mai essere di uguali.
Le due divisioni successive delle cellule della linea germinale dividono il processo unico
della meiosi in una meiosi I e in una meiosi II; fondamentalmente, due processi di
mitosi in successione. Il punto focale della meiosi è rappresentato dagli eventi della
meiosi I, durante la quale avvengono i fenomeni di rimescolamento genico di cui si
accennava prima.
Fasi della Meiosi e raffronti con la Mitosi
Le fasi della meiosi I. Le differenti fasi della mitosi prima descritte prestano il loro
nome anche alle fasi meiotiche, ciò che tuttavia accade nelle analoghe fasi della meiosi
può essere anche molto diverso. Nella profase della meiosi I (figura 13), infatti, i
cromosomi, composti come si diceva da due cromatidi fratelli, oltre ad essere nella fase
terminale della loro spiralizzazione compiono un processo aggiuntivo. Essi sono in grado
di riconoscere e di affiancare i rispettivi cromosomi omologhi presenti nella stessa cellula.
Questo appaiamento forma perciò strutture composte da quattro cromatidi paralleli, due
dei quali sono fratelli, e quindi uguali, e due dei quali sono invece omologhi ovvero solo
con la stessa sequenza genica. Queste strutture sono chiamate tetradi ed in esse si
svolge un processo chiamato crossing-over.
Il crossing-over è un fenomeno centrale nella variabilità genetica prodotta dalla
formazione dei gameti. Durante questo processo, interi tratti di cromosoma possono
essere scambiati da un cromatidio ad un altro tra quelli componenti la tetrade, purchè il
tratto scambiato si trovi alla stessa altezza (regioni omologhe) e che la sequenza dei
geni venga rigorosamente conservata (figura 12).
Ciò che viene scambiato, quindi, sono geni che possono essere diversi solo in alcuni punti
della loro sequenza di nucleotidi, ad esempio alleli dominanti con i loro alleli omologhi ma
recessivi; oppure anche geni entrambi a carattere dominante ma con sequenze
nucleotidiche non identiche.
La meiosi I, a questo punto, incontra le fasi di metafase I e anafase I (figura 13).
Meccanicamente esse si svolgono come nella mitosi; la differenza sostanziale sta nel fatto
che, nella metafase I della meiosi, i cromosomi migrano al piano dell’equatore nella
forma di tetradi. Nella successiva anafase I, la migrazione ai poli non produce la
separazione dei cromatidi fratelli come in mitosi, ma solo la separazione dei cromosomi
omologhi delle tetradi, ancora composti dai due cromatidi fratelli.
La meiosi I si conclude con una telofase e una citodieresi uguali a quelle della divisione
mitotica con la differenza che non avviene la ricostruzione completa dei nuclei in quanto
le cellule devono andare incontro ad una nuova divisione.
17
Cromosomi
omologhi
Gianmario Gerardi
Centromero
Cromatidi
fratelli
Centromeri
Scambio di
tratti di
cromosoma
Cromatidi fratelli
Figura 12 - Crossing-over. I cromosomi, composti da due cromatidi fratelli, sono in grado di riconoscere e
affiancare i rispettivi cromosomi omologhi presenti nella stessa cellula. L’appaiamento forma le tetradi in cui si
svolge il processo del crossing-over. Intere regioni omologhe di cromosoma possono essere scambiate da un
cromatidio ad un altro conservando rigorosamente la sequenza dei geni. In alto a destra una fase di crossingover al microscopio elettronico.
(Ricostruzione grafica da materiale web assortito)
Approfondimenti e Richiami
Il riassortimento indipendente dei geni. Questa migrazione opposta dei cromosomi omologhi è una fonte
secondaria di variabilità genetica, in quanto non esistono meccanismi in grado di discriminare quale dei due
cromosomi omologhi debba raggiungere un polo della cellula piuttosto che l’altro. Pertanto, se prendiamo in
considerazione solo geni situati su cromosomi diversi, essi effettueranno una segregazione ai poli della cellula
che fra loro sarà di tipo indipendente. L’effetto è che, geni dominanti o recessivi che siano, situati su cromosomi
diversi, subiranno un riassortimento indipendente e casuale. In più, il fenomeno del crossing-over fa in modo
che tale riassortimento casuale possa avvenire anche per geni che risiedevano sullo stesso cromosoma, grazie
allo scambio di tratti di cromosoma all’interno delle tetradi. Geni presenti sullo stesso cromosoma, ridurranno la
loro probabilità di segregare indipendentemente dagli altri geni dello stesso cromosoma man mano saranno più
vicini.
Le fasi della meiosi II. La meiosi II è un altro processo di mitosi a tutti gli effetti, che
avviene a carico di ciascuna delle due cellule prodotte dalla divisione della meiosi I. Ciò
che cambia non sono gli eventi, ma solo la composizione cromosomica delle cellule
originarie e di quelle figlie.
La profase II della meiosi (figura 14) non prevede un riarrangiamento strutturale del
DNA, il quale si conserva nello stato di compattamento proveniente dalla meiosi I. Ciò
che accade con regolarità, analogamente alla profase mitotica, è la formazione del fuso
mitotico.
18
Gianmario Gerardi
A
profase I
B
metafase I
C
anafase I
D
telofase I
E
interfase II
Figura 13 – Meiosi I. La prima divisione del processo meiotico è chiamata Meiosi I. Si tratta di una sostanziale
mitosi, ma durante la profase ogni cromosoma si affianca al proprio omologo formando i chiasmi (A). Nella
metafase i cromosomi sono regolarmente disposti sul piano equatoriale della cellula, ma ogni coppia di
cromosomi omologhi è associata alle fibre del fuso lungo la stessa linea (B). Nella anafase I, infatti, le fibre del
fuso non separano i cromatidi fratelli, come nella mitosi, ma separano ogni cromosoma omologo. Nella telofase
e nella citodieresi (D-E) si abbozzano due nuclei contenenti una singola collezione di cromosomi omologhi.
(Ricostruzione grafica: Curtis, Barnes – Invito alla Biologia; Zanichelli)
Nella metafase II della meiosi, avviene la regolare separazione e segregazione dei
cromatidi fratelli. Non va però dimenticato che l’insieme dei cromatidi fratelli, che ora si
stanno separando e quindi sono cromosomi a tutti gli effetti, compongono un corredo
aploide (n), perché nessuno di essi possiede più la sua parte omologa all’interno della
stessa cellula. Ogni cromosoma omologo è stato precedentemente confinato in una
cellula differente durante la divisione della meiosi I (figura 14).
Alla metafase II succedono regolarmente la telofase II e la citodieresi II le quali
ricostituiscono correttamente i due nuovi nuclei ed effettuano la seconda divisione vera e
propria, con la formazione finale di un totale di quattro cellule.
Queste quattro cellule sono i quattro gameti, i quali, successivamente, concluderanno se
necessario il loro differenziamento morfologico che li renderà adeguati al loro ruolo.
Ogni gamete conterrà perciò un corredo aploide (n) costituito da una sola copia per
ciascun gene del patrimonio complessivo dell’organismo.
19
Gianmario Gerardi
A
profase II
B
metafase II
C
anafase II
D
telofase II
E
interfase II
Figura 14 – Meiosi II. La seconda mitosi del processo meiotico è chiamata anche Meiosi II. Rispetto alla
divisione avvenuta durante la meiosi I, la meiosi II è una mitosi vera e propria durante la quale avviene la
separazione dei cromatidi fratelli. Nelle due cellule originatesi dalla meiosi I, viene organizzato un nuovo fuso
mitotico e i cromosomi, presenti in una sola copia omologa si ridistribuiscono in modo che ciascuno possa
legare il proprio cinetocoro ad una delle fibre del fuso (A, profase II). Durante la metafase II il fuso mitotico è
maturo e i cromosomi si trovano allineati sul piano equatoriale della cellula (B) e nell’anafase II e telofase II i
due cromatidi di ciascun cromosoma migrano in direzione opposta verso i poli della cellula. Nell’interfase II le
quattro cellule finali riorganizzano i propri nuclei. Il corredo cromosomico di ciascuna è aploide (n).
(Ricostruzione grafica: Curtis, Barnes – Invito alla Biologia; Zanichelli)
La gametogenesi (figura 15). Nell’uomo, e in generale nei vertebrati maschi, la meiosi
delle cellule della linea germinale si svolge nei testicoli e termina con la formazione degli
spermatozoi. Le cellule dei tubuli seminiferi delle gonadi maschili si dividono per mitosi
come spermatogoni, fino al differenziamento di spermatociti primari, i quali iniziano il
processo meiotico. Al termine della meiosi I le cellule prendono il nome di spermatociti
secondari, mentre al termine della meiosi II le cellule, con il loro corredo gametico
apolide (n) definitivo, vengono chiamate spermatidi. Gli spermatidi, successivamente, si
differenziano in spermatozoi, sviluppando il flagello e assumendo la forma specifica. Il
processo inizia negli individui dopo un periodo di maturazione chiamato pubertà e
continua all’incirca per tutta la vita.
Nelle femmine, invece, la meiosi I prende inizio già negli organi genitali del feto durante
la vita intrauterina (nella donna, circa al terzo mese). Nelle ovaie, all’interno di strutture
preposte chiamate follicoli, le cellule che iniziano la meiosi sono gli oociti primari, i
quali, però, termineranno il processo di meiosi I solo all’epoca dell’ovulazione, durante
l’età fertile della donna, quindi anche molti anni dopo. La meiosi I non produce però due
cellule uguali, ma un oocita secondario grande e una cellula molto più piccola chiamata
20
Gianmario Gerardi
corpuscolo polare. La meiosi II, invece, si svolge nella breve fase successiva alla
fusione dell’ovulo con uno spermatozoo e, anch’essa, effettua una divisione asimmetrica,
portando ad un grande ovulo vero e proprio e ad un altro piccolo corpuscolo polare. Esso,
insieme ai due corpuscoli polari prodotti dalla meiosi II del primo corpuscolo, finirà per
degenerare. Se la fecondazione dell’ovulo non ha luogo, perciò, la meiosi II non si
conclude e il gamete femminile non raggiungerà mai la maturazione finale.
Lo zigote. La fusione del gamete aploide di un individuo maschio con il gamete prodotto
da un individuo femmina produce una nuova cellula diploide, nella quale ciascun
cromosoma ritrova la sua parte omologa all’interno della stessa cellula. Ciascun gene
sarà nuovamente rappresentato da due alleli omologhi, i quali avranno però un
assortimento differente rispetto agli alleli degli altri geni e rispetto a quello dei genitori da
cui sono provenuti i gameti. Questa nuova cellula, che darà origine ad un nuovo individuo
completo, è chiamata zigote; da essa si svilupperà l’embrione.
Figura 15 – Gametogenesi dei vertebrati. Gli spermatociti primari nel maschio e gli ovociti primari nella
femmina iniziano il processo meiotico. Al termine della meiosi I, gli spermatociti hanno generato 2 spermatociti
secondari, mentre l’ovocito si è diviso in un ovocito secondario e in un piccolo corpuscolo polare. Le cellule
passano nella fase di meiosi II: i due spermatociti secondari si dividono in 4 spermatidi, mentre l’ovocita
secondario e il suo corpuscolo polare si dividono in un ovulo maturo e 3 nuovi corpuscoli polari. Al termine della
meiosi II le cellule hanno un corredo genetico apolide (n).
(Ricostruzione grafica da materiale web assortito)
21
Gianmario Gerardi
GLOSSARIO
Acido desossiribonucleico: (o DNA) macromolecola biologica costituita da due catene polimeri di nucleotidi,
appaiate longitudinalmente attraverso interazioni di tipo ponte-H tra le basi azotate costituenti i nucleotidi.
Acido ribonucleico: (o RNA) macromolecola biologica del tutto simile a quella dell’acido desossiribonucleico,
differente da quest’ultimo per la presenza di un ossidrile posto sull’anello dello zucchero ribosio di ciascun
nucleotide e per il fatto che la base azotata Timina, è sostituito dalla base Uracile.
Adattamento: in gergo biologico evolutivo, è l’insieme di modificazioni del fenotipo che intercorrono lungo le
generazioni di una stessa specie di organismi, in grado di aumentare le probabilità di sopravvivenza e di
riproduzione degli individui grazie ad una migliore interazione con l’ambiente circostante.
Allele: versione alternativa di uno stesso gene specifico, presente all’interno di uno stesso genoma. Il numero
di alleli per ciascun gene è indicato dal grado di ploidia della specie di appartenza: specie aploidi (n) possiedono
un unico allele, specie diploidi (2n) ne possiedono due e così via. Gli alleli di uno stesso gene sono caratterizzati
da interazioni reciproche di dominanza o recessività.
Anafase: fase del processo di divisione di una cellula nella quale i cromosomi che sono stati separati alla fine
della metafase (cromatidi fratelli nella mitosi o cromosomi omologhi nella prima divisione meiotica) agganciati
alla stessa fibra del fuso mitotico migrano rispettivamente verso i poli opposti della cellula.
Centrioli: organuli cellulari costituiti da una coppia di cilindri formati da nove triplette di brevi microtubuli
paralleli. La loro attivazione avviene nella profase della divisione cellulare, durante la quale essi migrano ai poli
della cellula e formano attorno a sé una regione di assemblaggio delle strutture microtubulari che prende il
nome di centrosoma.
Centromero: regione cromosomica più o meno centrale, priva di sequenze geniche codificanti, visibile al
microscopio ottico nelle fasi di spiralizzazione del cromosoma come una strozzatura e deputata, insieme ai
cinetocori, all’ancoraggio del cromosoma alle fibre del fuso mitotico durante le fasi di divisione cellulare.
Centrosoma: insieme dei centrioli e della regione attiva circostante di assemblaggio delle strutture
microtubulari, visibile al microscopio ottico a partire dalla profase e fino alla tarda anafase della divisione
cellulare. Dal centrosoma si orgina tutto il fuso mitotico.
Cinetocoro: struttura proteica discoidale annessa al centromero del cromosoma in grado di legarsi alle fibre
del fuso mitotico.
Citodieresi: divisione vera e propria della cellula al termine del processo più complesso della mitosi o delle
divisioni meiotiche.
Codice genetico: codice di traduzione della sequenza di nucleotidi di un gene nella sequenza di aminoacidi
costituenti la proteina codificata da quel gene. Ogni aminoacido è codificato da un numero variabile (da uno a
sei) possibili combinazioni di triplette di nucleotidi chiamate codoni.
Corpuscolo polare: cellula derivante dalle divisioni meiotiche di un oocito (ovocito) primario, molto più piccola
della cellula uovo e destinata a degenerare. Durante la meiosi, a partire da un oocito primario si formeranno tre
corpuscoli polari ed una sola cellula uovo matura.
Cromatidi fratelli: elementi costitutivi di un cromosoma profasico, costituiti dalla spiralizzazione di due
filamenti di DNA uguali provenienti dalla replicazione di un filamento originario, uniti fra loro nella zona del
centromero. I cromatidi fratelli vengono separati e destinati alle due cellule figlie durante il processo della
mitosi.
Cromatina: massa indistinta di cromosomi despiralizzati presenti nel nucleo della cellula durante il periodo di
interfase. Viene a sua volta suddivisa in eterocromatina ed eucromatina.
Cromosoma: filamento unico ed ininterrotto di DNA a doppia elica presente nel nucleo delle cellule in numero
specifico a seconda della specie; visibile in modo distinto al microscopio ottico nella sua forma compatta
spiralizzata durante le fasi di mitosi o meiosi cellulare, sottoforma di bastoncello. La spiralizzazione di un
filamento di DNA produce un accorciamento del filamento anche di sei ordini di grandezza.
Cromosomi omologhi: cromosomi costituiti da filamenti di DNA all’interno di una stessa cellula, recanti
un‘identica sequenza di geni. Si distinguono fra loro per la possibile presenza di differenze nella sequenza
nucleotidica di ciascun gene. Il numero di cromosomi omologhi per ciascun cromosoma è indicato dal numero di
ploidia: cellule diploidi (2n) hanno due cromosomi omologhi per ciascun tipo di cromosoma.
22
Gianmario Gerardi
Crossing-over: processo caratteristico della prima profase della meiosi, durante il quale i cromosomi omologhi
si appaiano, affiancandosi in modo che ogni gene risulti affiancato al suo gene omologo. Durante il processo,
porzioni omologhe di cromatidio vengono fisicamente scambiate tra un cromosoma omologo e l’altro.
Elettronegatività dell’ossigeno: capacità di attrarre su di sé elettroni nella formazione di un legame chimico
(valore 3,5 nella scala di Pauling).
Energia cinetica: energia di moto di atomi e molecole, espressa in termini fisici come metà della massa di un
corpo moltiplicata per il quadrato della sua velocità.
Energia potenziale: energia intrinseca degli atomi e delle molecole sostanzialmente confinata all’interno dei
legami. L’energia potenziale è in generale la capacità di un corpo o di un sistema di compiere un lavoro, in virtù
della sua posizione rispetto a un campo di forze.
Entalpia: funzione di stato esprimente l’energia interna di un sistema sommata al lavoro meccanico prodotto in
condizioni di pressione costante e corrispondente al calore scambiato. Nel caso di molecole è possibile
affermare che l’entalpia è l’insieme dell’energia potenziale e dell’energia cinetica di quella molecola.
Entropia: grado di disordine di un sistema, relativo a stadi temporalmente diversi di una trasformazione del
sistema o relativo al livello di disordine circostante.
Ermafroditismo: caratteristica costitutiva di alcuni esseri viventi rappresentata dalla compresenza sullo stesso
individuo di un organo genitale femminile e di un organo genitale maschile.
Eterocromatina: cromatina costituita da DNA cromosomico nel suo massimo livello di condensazione.
Eucromatina: cromatina poco condensata comprendente tratti di DNA attivamente trascritto. Il filamento di
DNA cromosomico è avvolto intorno ad ottametri di proteine chiamate istoni e la struttura risultante si trova a
sua volta raccolta in una spirale più o meno compatta.
Fase G0: fase del ciclo cellulare nella quale la cellula non è destinata alla riproduzione e alla divisione. In questa
fase, la cellula svolge le sue funzioni specifiche in attesa di segnali che inducano attività proliferativa.
Fase G1: fase del ciclo cellulare successiva alla divisione. Cellule provenienti da un evento mitotico entrano in
questa fase di sintesi di proteine e di strutture che renderanno adulta la nuova cellula, e con un metabolismo
attivo rispetto alle funzioni specifiche della cellula.
Fase G2: fase del ciclo cellulare successiva alle fase S di replicazione del DNA, durante la quale la cellula, oltre
a compiere ancora parte delle sue funzioni metaboliche, è impegnata nella sintesi attiva di proteine e strutture
necessarie al processo imminente di mitosi.
Fase S: fase del ciclo cellulare successiva alla fase G1, o alla fase G0 nel caso si tratti di cellule provenienti da
tale fase di stasi replicativa. In questa fase avviene un’attiva sintesi di DNA necessaria alla replicazione
dell’intero patrimonio genetico.
Fenotipo: manifestazione effettiva e netta dell’attività genica in un organismo vivente. Il fenotipo corrisponde
all’insieme dell’attività genica non inibita, ovvero l’attività di un gene dominante o l’attività di un gene che, nello
stesso organismo, è presente solo nella sua forma recessiva. Il fenotipo può essere inteso relativamente a un
solo tipo di gene ma anche a più geni o all’intera attività genica di un organismo.
Fuso mitotico: struttura a forma di fuso costituita dai microtubuli prodotti dall’attività del centrosoma che si
dipartono da ciascun polo della cellula. Il fuso mitotico si forma dopo la scomparsa della membrana nucleare e
occupa gran parte della cellula metafasica.
Gamete: cellula sessuale aploide prodotta dal processo della meiosi, destinata alla fecondazione tramite la
fusione con un gamete del sesso opposto.
Gametogenesi: processo a carico delle cellule della linea germinale di un individuo necessario alla formazione
dei gameti e composto in buona parte dalle fasi costituenti la meiosi.
Genotipo: corredo genetico dell’organismo, indipendentemente da ciò che l’attività del gene di fatto
manifesterà. Il genotipo è la dotazione effettiva di geni indipendentemente dal fatto che essi siano attivi, siano
dominanti o recessivi. Il genotipo può essere inteso relativamente a un solo tipo di gene ma anche a più geni o
all’intero corredo genico di un organismo.
Interfase: fase del ciclo cellulare costituente la vita di una cellula che intercorre tra una divisione e l’altra. Essa
può essere composta dalle fasi G0, G1, S e G2.
Lamella mediana: sottile lamina di separazione posta tra le pareti di cellule vegetali adiacenti, costituita da
polisaccaridi e pectina. L’elemento è comune alle due cellule adiacenti e proviene dalla cellula madre che ha
generato per mitosi le due cellule.
23
Gianmario Gerardi
Linea germinale: classe di cellule destinate alla produzione dei gameti sessuali attraverso fenomeni di
differenziamento e attraverso il processo della meiosi.
Metafase: fase del processo cellulare di mitosi e meiosi durante il quale i cromosomi destinati alla suddivisione
tra le due cellule figlie, ovvero coppie di cromatidi fratelli nella mitosi o coppie di cromosomi omologhi nella
prima divisione meiotica, appaiono al microscopio ottico allineati sul piano equatoriale della cellula, e agganciati
alle rispettive fibre del fuso mitotico.
Muoni o raggi cosmici: particelle di origine subatomica provenienti dallo spazio, in larga parte originate dal
vento solare e dotate di carica negativa.
Mutazione: variazione stabile e sostanzialmente irreversibile nella costituzione di un gene, può risultare del
tutto irrilevante ai fini della vita dell’organismo che l’ha subita oppure profondamente significativa e
determinante.
Nucleotide: monomero degli acidi nucleici costituito da un anello di ribosio, da una base azotata e da un
gruppo fosfato.
Oocito: cellula della linea germinale femminile che per meiosi darà origine alla cellula uovo e a tre corpuscoli
polari che degenerano.
Patrimonio genetico: insieme dell’intero corredo genico di un organismo vivente.
Piastra di separazione: deposizione equatoriale mediana effettuata da una cellula vegetale in divisione,
costituita da polisaccaridi e pectina sulla quale verrà successivamente depositata la parete vera e propria delle
due nuove cellule figlie. La piastra di separazione si modificherà leggermente diventando la lamella mediana.
Ploidia: indica il numero delle serie di cromosomi omologhi all’interno dello stesso organismo. Il numero di
cromosomi omologhi per ciascun tipo di cromosoma è espresso da un numero n. Cellule con un’unica serie di
cromosomi omologhi avranno ploidia 1 e saranno chiamate cellule aploidi (1n). Analogamente, cellule diploidi
presentano due serie di cromosomi omologhi (2n), cellule triploidi tre serie (3n) e così via.
Profase: fase del processo cellulare di mitosi e meiosi durante il quale i cromosomi cominciano a rendersi
visibili in modo abbastanza distinto al microscopio ottico; inizia lo sfaldamento della membrana nucleare e la
migrazione dei centrioli ai poli della cellula.
Riproduzione sessuata: riproduzione di organismi che prevede il concorso di due organismi della stessa
specie appartenenti a sessi opposti.
Selezione naturale: fenomeno rappresentato dall’affermazione degli individui meglio adattati all’ambiente
rispetto a quelli meno compatibili. La selezione naturale privilegia individui dotati di un corredo genetico
globalmente in grado di conferire maggiori probabilità di sopravvivenza e di riproduzione.
Sequenza genica: successione più o meno continua e specifica di geni lungo il filamento di DNA di un
cromosoma. All’interno di ogni specie, l’ordine della sequenza di geni lungo ogni cromosoma è rigorosamente
conservata.
Sequenza nucleotidica: successione continua e specifica di nucleotidi all’interno di un tratto di DNA
corrispondente a un gene, a una parte di esso o a regioni di DNA con funzione non codificante. All’interno della
stessa specie le sequenze nucleotidiche dei cromosomi sono complessivamente molto conservate pur
presentando piccole variazioni. Tali variazioni possono conferire a uguali geni attività geniche diverse, in modo
più o meno marcato. Per questo, all’interno di una popolazione, vi sono geni che risultano dominanti rispetto ad
altri. Questi ultimi sono detti recessivi.
Specie: raggruppamento di organismi viventi tra i quali l’incrocio riproduttivo risulta costantemente fecondo.
Tale caratteristica distintiva rappresenta un fondamentale elemento tassonomico classificativo.
Spermatidio: cellula aploide della linea germinale maschile. A partire da uno spermatocita si ottengono, al
termine della seconda divisione meiotica, quattro spermatidi che andranno incontro alla parte finale del
differenziamento in spermatozoi.
Spermatocita: cellula della linea germinale maschile, originate per mitosi da spermatogoni. Lo spermatocita
primario va incontro alla prima divisione meiotica e lo spermatocita secondario alla seconda divisione meiotica.
Spermatogonio: cellula della linea germinale maschile diploide. Gli spermatogoni sono presenti per tutto il
periodo riproduttivo e si dividono per mitosi.
Telofase: fase di divisione di una cellula nella quale i cromosomi hanno compiuto definitivamente la migrazione
verso i poli opposti della cellula, è iniziata la formazione di due nuovi nuclei e lungo il piano equatoriale sta
formandosi una strozzatura che diventerà il solco di separazione.
24
Gianmario Gerardi
Tetradi: strutture costituite dall’appaiamento di due cromosomi omologhi, ciascuno costituito dai suoi due
cromatidi fratelli. Le tetradi si formano durante la profase della prima divisione meiotica e permettono che
avvenga il crossing-over.
Traduzione: complesso processo durante il quale l’informazione portata dalla sequenza nucleotidica di un RNA
messaggero viene convertita nella sequenza aminoacidica di una proteina.
Trascrizione: processo attraverso il quale viene sintetizzato RNA messaggero a partire dalla lettura di una
sequenza codificante di DNA.
Uracile: base azotata specifica degli acidi ribonucleici, presente in sostituzione alla timina degli acidi
desossiribonucleici.
Zigote: cellula originata dalla fusione di due gameti di sesso opposto nella quale il corredo cromosomico è stato
riportato al suo grado corretto di ploidia unendo il patrimonio genetico di un gamete con quello dell’altro.
25
")