15/03/2012
A. Crisafulli
F. Podda
INTRODUZIONE CUORE, APPARATO CIRCOLATORIO
Formato dal cuore e dai vasi sanguigni che possono essere suddivisi in:
• Vasi di distribuzione: arterie
• Vasi di scambio: capillari
• Vasi di raccolta: vene
La funzione di questo sistema è quella di trasportare sangue e garantire la sopravvivenza e la
funzionalità ottimale dei tessuti.
La circolazione è suddivisa in due sistemi collegati in serie tra loro: il circolo polmonare a
bassa pressione (15 mmHg) e il circolo sistemico ad alta pressione (100mmHg)*
Il piccolo circolo: atrio destro → ventricolo destro → arterie
polmonari polmoni (scambio gassoso, ossigenazione) → vene polmonari
Il grande circolo: atrio sinistro → ventricolo sinistro → aorta →
organismo (scambio gassoso tissutale) → vene cave → atrio destro
L’energia utile per vincere le resistenze emodinamiche, viene fornita
dal cuore, che permette al sangue di scorrere nel torrente circolatorio.
Tuttavia il cuore non è il solo elemento a fornire quest’energia,
intervengono anche i meccanismi delle pompe muscolari e della
respirazione; quest’ultimo meccanismo permette il ritorno venoso del
sangue dalla periferia agendo sulle variazioni di pressione. Durante
l’inspirazione la pressione interna della gabbia toracica diminuisce
richiamando sangue dalla periferia
*si tratta di valori medi
Le valvole cardiache e la loro tenuta sono fondamentali per il
corretto funzionamento di tutto il sistema; devono garantire
l’unidirezionalità del flusso ematico e la loro conformazione anatomica
permette di svolgere al meglio questo compito. Se la tenuta valvolare viene meno si avranno delle
patologie più o meno gravi (es: soffio cardiaco)
Il cuore è formato da uno scheletro fibroso sul quale prendono inserzione le quattro valvole e
il tessuto muscolare (si differenzia dal t. muscolare scheletrico perché le cellule non formano un
sincizio anatomico, pur mantenendo il sincizio “funzionale” importante per la propagazione
dell’impulso elettrico attraverso le aree di bassa resistenza elettrica quali sono i dischi intercalari).
I miocardiociti sono ricchissimi di mitocondri, il metabolismo cardiaco è totalmente aerobico.
Il cuore, a sua volta, è irrorato dalle due arterie, coronaria destra e coronaria sinistra. La
variabilità anatomica è molto ampia, ma generalmente il lume della sinistra è maggiore e si sfiocca
quasi subito nel ramo circonflesso e nel ramo discendente.
Le arterie coronarie sono vasi terminali, hanno pochissime anastomosi e questo si ripercuote
sull’ossigenazione cardiaca in caso di occlusione; l’infarto del miocardio è dato dalla morte per
necrosi ipossica del tessuto.
Il cuore ha una frequenza contrattile intrinseca di 90-100 bpm (frequenza stabile dei
trapiantati cardiaci) e l’innervazione autonoma si limita alla modulazione di questo ritmo.
Ortosimpatico e Parasimpatico lavorano “in opposizione”.
• Parasimpatico (recettori muscarinici) → nervi vaghi → atri → riduce frequenza
• Simpatico (recettori β per Adrenalina e Noradrenalina) → atri e ventricoli → aumenta forza
e frequenza.
(notare che il parasimpatico agisce solo su atri e frequenza e non sulla forza.)
In condizioni di riposo il cuore è sottoposto prevalentemente all’innervazione vagale e quindi
agli effetti del parasimpatico, mentre in caso di stress o esercizio fisico, si ha prevalenza
dell’ortosimpatico.
Il cuore ha tre tipi di tessuto
-Tessuto muscolare propriamente detto
-Tessuto nodale
-Tessuto di conduzione
Il tessuto nodale e quello di conduzione sono particolari specializzazioni del tessuto
muscolare.
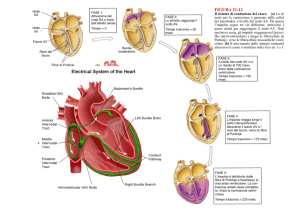
Il Nodo del seno e il Nodo atrio-ventricolare fanno parte del tessuto nodale, questo si
continua poi con il Fascio di His che invece fa parte del tessuto di conduzione.
Il Fascio di His si divide, a cavallo del setto interventricolare, in una branca destra e una
branca sinistra che a sua volta si dividerà ancora in un fascio anteriore e uno posteriore; le fibre
terminali di queste branche son dette Fibre del Purkinje. Tutto questo sistema di conduzione serve a
trasmettere l’impulso generato dalle cellule pacemaker a tutto il tessuto muscolare contrattile e
modularne l’ ”ordine” di contrazione.
Nodo seno-atriale → muscolatura atri → Nodo atrio-ventricolare → Fascio His → fibre
Purkinje → base ventricolo
17-03-2012
Crisafulli
Claudia Maggiore
RIEPILOGO LEZIONE PRECEDENTE:
La scorsa lezione abbiamo parlato dell’apparato circolatorio,ricordando i fondamentali
anatomici e andando ad esaminare la direzione del flusso sanguigno nel grande e nel piccolo
circolo. Abbiamo pure parlato della conformazione di atri e ventricoli e delle valvole cardiache, la
cui funzione è quella di imprimere al sangue una certa direzione, unica, evitando reflussi. Abbiamo
anche visto che il cuore stesso è irrorato dal sistema sanguigno grazie alle arterie coronarie che
assicurano il trofismo del cuore. Questo inoltre è innervato dal S.N.A., soprattutto dal simpatico,
mentre l’innervazione parasimpatica è limitata per lo più agli atri. Il cuore è formato
principalmente da 3 tipi di tessuto. Per la gran parte si tratta di tessuto muscolare che si comporta,
da un punto di vista sia meccanico che biochimico, similmente al muscolo scheletrico (i ponti actina
e miosina si formano allo stesso modo), e poi da una serie di tessuti specializzati: il tessuto nodale e
il tessuto di conduzione che servono a fare in modo che si generi un impulso spontaneo nel cuore
che da il via al battito cardiaco. Questo impulso viene poi trasmesso nella periferia cardiaca
attraverso un sistema di conduzione che garantisce la sincronia nel battito, di modo che prima si
contraggano gli atri e poi i ventricoli. E’ importante che anche all’interno del ventricolo l’attività sia
sincrona, infatti si deve prima contrarre il setto e i muscoli papillari (muscoli che tengono in sede,
attraverso dei tendini, le valvole mitralica e tricuspide) e poi il resto della muscolatura.
Le valvole atrio-ventricolari non sono libere ma i bordi sono legati a un sistema tendineo (le
corde tendinee) che a sua volta è legato ai muscoli papillari (piccoli muscoli che originano dal
miocardio). Queste valvole shanno questa struttura perché altrimenti rischierebbero, con la
contrazione atriale, di essere trascinate negli atri, cosa che peraltro succede in alcune persone
(prolasso valvolare mitralico, patologia molto diffusa nella popolazione). Quindi prima che si
contragga tutto il cuore devono contrarsi i muscoli papillari in modo che mettano in tensione le
corde tendinee evitando che le valvole atrio-ventricolari vadano si insinuino negli atri.
Abbiamo visto quali sono le caratteristiche del miocardio: è un tessuto muscolare a tutti gli
effetti, l’unica grande differenza con il tessuto muscolare scheletrico è che più che essere un
sincizio anatomico è un sincizio funzionale.
Abbiamo parlato anche del tessuto nodale e in particolar modo del nodo seno atriale che è la
struttura dove normalmente parte l’impulso che poi si dirige agli atri, poi al nodo atrio-ventricolare
e da lì a tutto il cuore grazie al sistema di conduzione.
------
------
------
------
Le quattro proprietà fondamentali del tessuto cardiaco sono:
l’eccitabilità, la ritmicità, la conducibilità e la contrattilità.
1) ECCITABILITÀ (batmotropismo):
in termini generali si definisce eccitabilità di una cellula o di un tessuto la capacità della
cellula stessa di rispondere a uno stimolo con un cambiamento di stato. Lo stimolo adeguato per
ottenere una risposta in un tessuto cardiaco è la modificazione del potenziale elettrico a riposo della
cellula, che normalmente è di circa – 90mv. La risposta del cuore a uno stimolo efficace è la
contrazione, cioè il potenziale d’azione è la contrazione, come accade similmente nel muscolo
scheletrico.
Perché le cellule cardiache sono eccitabili?
Le cellule cardiache sono eccitabili perché dotate di un potenziale di riposo, cioè perché la loro
membrana cellulare è polarizzata principalmente grazie all’azione della pompa sodio-potassio. Le
cellule cardiache infatti tendono ad estrudere il potassio fuori dalla cellula (infatti c’è una grande
differenza di concentrazione fra i due compartimenti) e contemporaneamente, per compensare le
cariche, a portare dentro potassio. Per ogni sodio che esce, entra dentro un potassio. Tutto ciò
avviene ovviamente con spesa energetica(ATP). Ma il sodio e il potassio non sono gli unici ioni a
determinare il potenziale di membrana delle cellule cardiache, vi partecipa anche il cloro. Il cloro si
distribuisce passivamente ai due lati della membrana in base a un gradiente elettrochimico, cioè
sulla base dell’elettronegatività che trova all’interno. L’elettronegatività prevalente interna di queste
cellule è dovuta peraltro alla presenza di proteine. Quindi a causa del fatto che si estrudono cariche
positive e che troviamo anche proteine all’interno della cellula si crea un ambiente elettronegativo
rispetto all’esterno. Altri ioni che partecipano alla genesi di questo potenziale di membrana sono:
-Ca2+ (la percentuale di calcio libero all’interno della cellula è bassa perché lo ione viene
soprattutto requisito dal reticolo sarcoplasmatico)
-Mg2+
Nella realtà delle cose la genesi del potenziale di membrana di queste cellule è abbastanza
complessa. Comunque è importante sapere che la principale corrente è generata dalla pompa sodiopotassio che estrude fuori sodio (che non può rientrare dentro perché i cancelli al sodio sono chiusi
e rientra solo se si crea il potenziale d’azione), e il potassio che invece viene portato dentro. Tutto
ciò genera questo potenziale elettrico.
Come si fa a calcolare il potenziale elettrico non solo di questa cellula cardiaca ma in
generale?
Si usa l’equazione di Nernst che serve a valutare qual è il potenziale transmembrana di una
cellula dove gli ioni sono distribuiti in maniera disuguale a cavallo delle due membrane.
Ek=(RT/ZF)ln([K]e/[K]i)
In questo caso è l’equazione di Nernst è applicata al potassio, ma noi possiamo usarla per
qualsiasi altro ione.
Tenendo presente che i valori dell’equazione (RT/ZF) sono costanti, questa equazione si
riscrive in una forma più semplificata:
Ek=60log([K]e/[K]i).
Se noi dunque applichiamo quest’equazione alle concentrazioni di potassio troviamo un
valore di circa-94mv. Andando a misurare il potenziale di membrana di un miocita con degli
elettrodi (uno messo dentro la cellula e uno messo fuori) troviamo dei valori che oscillano dai -85 ai
– 95mv; ciò vuol dire che il potenziale del potassio fornisce più o meno un valore di quella che è la
misura reale. Questo perché il potenziale di una membrana semipermeabile come quella del
miocita, dipende essenzialmente dal potenziale dello ione di cui, in quel momento, la cellula è più
permeabile.
Ad esempio se ci sono 100 ioni ma la permeabilità ce l'ha uno solo mentre gli altri sono
scarsamente permeabili, il valore del potenziale di membrana lo darà quell’unico ione che è
permeabile. In questo caso per il miocita è il potassio. Infatti se andiamo a prendere le
concentrazioni, ad esempio, di sodio troviamo un valore di + 60mv, che non è il valore reale della
membrana. C’è uno sfalsamento fra questi due risultati perché il sodio non è permeabile, infatti se
lo fosse dovremmo trovare +60mv nella misurazione reale. Il sodio non passa attraverso la
membrana e la concentrazione a cavallo della membrana è mantenuta per il fatto che i canali al
sodio sono chiusi e quindi lo ione non può passare.
Se noi applichiamo l’equazione di Nernst al cloro troviamo che il valore è -80 mV, valore
simile a quello di membrana, il cloro è in parte permeabile e in linea di massima tende a disporsi a
cavallo della membrana sulla base del potenziale di membrana stesso. Infatti ricordiamo che
all’interno della cellula ci sono più cariche negative,per la presenza di proteine che non possono
uscire,quindi il cloro tende ad uscire fuori per motivi di gradiente elettrico, perché all’interno trova
più cariche negative e quindi tende a uscire.
Il potenziale di riposo dei miociti dipende in gran parte dal potassio che è lo ione più
permeabile nella membrana del miocita. Ciò è evidente anche per il fatto che se varia la
concentrazione di potassio ci sono conseguenze rilevanti. Un aumento ad esempio del potassio
extracellulare e/o una diminuzione del potassio intracellulare causano una depolarizzazione dei
miociti,cioè i miociti invece di avere -90mv come potenziale di membrana avranno -80/-70 mV di
potenziale; -70 tra l’altro è il valore soglia che scatena il potenziale d’azione. Quindi se c’è un
aumento di potassio extracellulare tale da depolarizzare le mie cellule fino a – 70 mv, la cellula si
depolarizza spontaneamente causando ad esempio problemi aritmici.
Spesso problemi renali creano un aumento di potassio extracellulare nel nostro corpo che non
viene escreto in maniera ottimale dal rene e ciò può determinare aritmie. Infatti una delle più grosse
difficoltà nelle patologie renali è quella di controllare costantemente i livelli di potassemia. Una
diminuzione di potassio extracellulare e/o un aumento intracellulare causano invece
iperpolarizzazione dei miociti. Questa è una condizione che si verifica più raramente (si può
verificare per disidratazione, terapie diuretiche, ridotta assunzione di potassio..).
Così come per il muscolo scheletrico, anche per i miociti esiste un valore di potenziale soglia al di
sopra del quale la cellula reagisce con un potenziale d’azione, e questo valore è di circa -70mv.
Il potenziale d’azione che si verifica è abbastanza simile a quello della cellula muscolare,
tuttavia dobbiamo considerare che la ripolarizzazione è molto più lunga rispetto alle cellule
muscolari scheletriche e ai neuroni.
Infatti per un neurone la durata è circa 1 – 2 millisecondi, per un muscolo scheletrico dai 3 ai
10 ms dipende dal muscolo. Per il muscolo cardiaco invece il periodo di recupero è estremamente
lungo: 200-300 ms e dipende dalla frequenza cardiaca,se ad esempio sono tachicardico dura meno,è
modulabile.
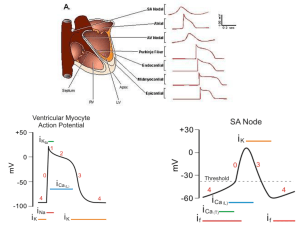
Il potenziale d’azione della cellula muscolare cardiaca come in tutte le cellule eccitabili
dipende dai movimenti ionici.
Nell'esempio abbiamo una cellula cardiaca a riposo con -90mv, se arriva uno stimolo efficace
che depolarizza la cellula questa risponde con un potenziale d’azione quindi con un’ascesa rapida
verso valori elettropositivi, fino a circa +20mV (fase zero), ciò è dovuto ad un ingresso massivo di
sodio all’interno della cellula per apertura dei canali per il sodio che per gradiente entra all’interno
della cellula. Il potenziale a questo punto non dipende principalmente dal potassio ma si avvicina al
potenziale del sodio che ora è lo ione più permeabile. Questi canali per il sodio rimangono aperti
per un tempo estremamente ridotto e in seguito inizia il ruolo di altri ioni.
Un piccolo ruolo all’inizio è dovuto al cloro che tende a uscire fuori dalla cellula creando una
ripolarizzazione rapida (fase 1), ma soprattutto c’è una lunga fase di ripolarizzazione lenta (fase 2 )
dove si aprono dei cancelli al calcio, il quale fuoriesce all’esterno, quindi la cellula diventa più
elettronegativa all’interno e contemporaneamente anche una piccola quota di potassio viene estrusa.
C’è poi una fase rapida di ripolarizzazione (fase 3), dovuta alla corrente di potassio che diventa
sempre più imponente fino a tornare alla fase 4, diastole, quando tutti i cancelli si chiudono e la
cellula è nuovamente a riposo. Il tutto ha una durata di circa 200 ms. E’ importante ricordare tutto
questo anche perché molti farmaci aritmici di utilizzo comune vanno proprio a influire su queste
correnti ioniche.
Ma nella cellula nodale le cose sono diverse: infatti la cellula nodale si occupa di dare il ritmo
al cuore, è la cellula dove nasce l’impulso per la depolarizzazione di tutte le altre cellule cardiache.
Quando siamo in diastole (cioè il cuore non si contrae), queste cellule hanno un potenziale d’azione
che non è stabile ma ha un valore che via via diventa più elettronegativo, cioè la cellula si
depolarizza pian piano da sola. Parte da valori di potenziale di membrana un po’ più bassi intorno ai
-60 mV e raggiunge il potenziale soglia intorno ai – 40 mV. C’è una corrente di ioni positivi (calcio
e sodio) in entrata che determina una depolarizzazione lenta di queste cellule. Queste cellule sono
infatti dotate di cancelli che fanno entrare poco sodio e quindi progressivamente depolarizzano la
cellula, inoltre c’è anche una corrente di calcio che normalmente non c’è a riposo;dunque la cellula
si avvicina al potenziale soglia e poi al potenziale d’azione con ingresso di sodio e di calcio in
maniera più massiva.
La velocità con cui questo meccanismo avviene dipende dall’innervazione parasimpatica e
ortosimpatica.
La frequenza cardiaca dipende dalla pendenza di questa corrente:cioè più velocemente la
cellula raggiunge il potenziale soglia più avremo una frequenza cardiaca maggiore.
Normalmente le cellule nodali hanno una frequenza intrinseca di circa 100 battiti al
minuto,poi il sistema nervoso autonomo modula questa frequenza e la fa aumentare o diminuire.
Finora abbiamo visto due tipi di cellule la cellula nodale e la cellula ventricolare;nella realtà
esistono varie altre cellule con comportamenti misti. Alcune come il nodo atrio-ventricolare si
contraggono autonomamente ma hanno una frequenza più bassa rispetto al nodo seno atriale. Ci
sono dunque due nodi in grado di contrarsi spontaneamente, uno che va a 100 l’altro a 50 (battiti al
minuto). Quello che batte a 50 non si manifesterà mai perché la frequenza di quello che batte a 100
lo sorpasserà di continuo: al nodo atrio-ventricolare arriverà l’impulso dal nodo seno atriale prima
che lui riesca a depolarizzarsi. Normalmente noi abbiamo un ritmo cardiaco che è quello del nodo
del seno, però abbiamo anche la possibilità di avere un ritmo che origini dal nodo atrio-ventricolare
caso mai il nodo del seno venga distrutto. Poi c’è anche il fascio di His e le cellule del Purkinje che
sono pure in grado di depolarizzarsi spontaneamente, anche loro possono diventare dei pacemakers,
questo però avviene solo se non ce ne sono altri più veloci.
Abbiamo dunque visto che esistono altri gruppi cellulari che hanno una spontanea
generazione d'impulso, ma con potenziali d’azione diversi e molti di questi in grado di fare da
pacemakers vicariante o se il “vero” pacemakers è danneggiato o se lo è la via di conduzione
(blocco atrioventricolare).
Come per il muscolo scheletrico anche per quello cardiaco esiste un periodo refrattario.
Il periodo refrattario è il periodo durante il quale lo stimolo applicato ad una cellula non produce
nessuna risposta. Il periodo refrattario della cellula miocardica si distingue in assoluto e relativo.
Il periodo refrattario assoluto si verifica qualora qualsiasi stimolo si applichi alla cellula, questa
non risponda con alcuna modificazione, poi c’è un periodo di 20 -30ms dove lo stimolo può
determinare una risposta che però è inferiore a quella che avremmo ottenuto se la cellula fosse stata
a riposo (periodo refrattario relativo):
cioè, se io applico degli stimoli in questo periodo (che peraltro devono essere più intensi degli
stimoli normali), non ottengo un potenziale d’azione classico ma un potenziale inferiore, sia per
durata sia per intensità.
Esiste poi, subito dopo il periodo refrattario relativo, un periodo chiamato di eccitabilità
supernormale, in cui degli stimoli anche sotto soglia possono scatenare il potenziale d’azione. Il
fatto che il periodo refrattario sia molto più lungo nel cuore rispetto alle altre cellule eccitabili, ha
un significato funzionale.
Nel cuore il picco di tensione (cioè attività meccanica) sviluppato dal miocita coincide con il
periodo refrattario, ne deriva che il miocardio non è tetanizzabile. (muscolo scheletrico: se si
mandano più stimoli vicini questi si sommano creando una tensione di intensità superiore rispetto a
quella di un singolo stimolo; rappresentabile come una linea piatta ).
Nel cuore questo fenomeno non è possibile perché il secondo stimolo lo posso avere solo alla
fine del periodo refrattario, che è molto lungo,e siccome il picco di tensione era avvenuto molto
prima non è possibile sommare gli impulsi. Il cuore quindi non può quindi irrigidirsi come fa il
muscolo. Dunque non è possibile ottenere una contrazione prolungata e duratura come si fa nel
muscolo col tetano. Questo ovviamente ha dei seri motivi fisiologici, infatti un cuore irrigidito è
incompatibile con la vita.
2)RITMICITÀ (CRONOTOPISMO): Proprietà del cuore di eccitarsi e quindi di contrarsi
ritmicamente e di mantenere ritmo costante(altrimenti ho una aritmia).
Normalmente il ritmo comincia dal nodo seno atriale, cioè in quel gruppo di cellule specializzate
che sono poste allo sbocco della vena cava superiore. Tuttavia, come già detto, il nodo seno atriale
non è l’unica struttura in grado di determinare ritmo cardiaco ma vi sono altre strutture come il
nodo atrio-ventricolare e il fascio di His.
Normalmente però queste cellule non intervengono perché la frequenza intrinseca del nodo
seno atriale è superiore a quella di queste strutture, per cui queste vengono sopraffatte dalla
frequenza del nodo seno atriale.
Nel nodo, come già detto, il potenziale d’azione dipende dall’ingresso nella cellula di sodio e
calcio che abbassano il valore del potenziale di membrana. La frequenza cardiaca dipende dalla
pendenza, velocità con cui questo fenomeno avviene. Modulando questo fenomeno si modula la
frequenza cardiaca. Noi normalmente abbiamo una frequenza intrinseca di 100 battiti al minuto su
cui poi agisce il nervo vago che, a riposo, porta la frequenza a circa 60/80 battiti al minuto. Il nodo
atrio-ventricolare invece ha una frequenza di scarica intorno ai 50 al minuto ed è meno modulabile
da parte del vago, rispetto al nodo del seno. Il fascio di His e la rete del Purkinje, infine, hanno una
frequenza di 20 -40 battiti al minuto. Questi dispositivi sono una serie di sistemi di sicurezza che
garantiscono (nei limiti) un buon funzionamento cardiaco. Frequenze cardiache troppo basse però
non sono adatte alla vita perché non permettono una resistenza del cuore nemmeno per i più
semplici moti (camminare, salire le scale). Quindi mentre il vago abbassa la frequenza cardiaca il
simpatico invece fa il contrario, tende ad aumentare la frequenza cardiaca ed aumentare la
pendenza con cui si depolarizzano queste cellule. Il fenomeno in realtà è più complesso e le cellule
adoperano meccanismi ancora in gran parte sconosciuti. Quello che si sa però è che le azioni del
parasimpatico sono mediate da un aumento della conduttanza al potassio, che esce dalla cellula,
rendendo l’interno ancora più elettronegativo e quindi iperpolarizzando la cellula. Le azioni del
simpatico sono invece mediate soprattutto da un aumento della conduttanza al calcio e quindi la
depolarizzazione che è dovuta anche a un ingresso di calcio viene accelerata. In questo modo il
S.N.A. modula la frequenza di scarica di queste cellule che poi danno il battito cardiaco.
3)CONDUCIBILITÀ (dromotopismo)
La conducibilità è quella proprietà dei tessuti cardiaci di condurre l’eccitamento nato nelle
cellule pacemakers. La propagazione dell’eccitamento avviene a velocità diverse nei vari tessuti,
che l’impulso incontra, dal nodo del seno fino alle cellule ventricolari, e il motivo per cui avviene
questo ritardo nell’eccitazione è un motivo funzionale.
La prima zona che si depolarizza normalmente è il nodo seno atriale che ha una velocità
abbastanza bassa di conduzione, poi l’impulso passa alla muscolatura atriale che è abbastanza
veloce a propagare l’impulso e a contrarsi e arriva alla giunzione atrioventricolare. Infine l’impulso
arriva al nodo atrio-ventricolare e qua subisce un rallentamento abbastanza importante per poi
passare al fascio di His. Qui la conduzione dell’impulso accelera nuovamente per portarsi al sistema
di Purkinje e alla muscolatura ventricolare. Il punto centrale che si deve ricordare è che al livello del
nodo atrio-ventricolare l’impulso subisce un deciso rallentamento. Il motivo di questo rallentamento
è che prima di far contrarre i ventricoli bisogna dare il tempo agli atri di svuotarsi nei ventricoli
stessi. Infatti se atri e ventricoli si contraessero insieme, dato che i ventricoli sono più potenti degli
atri e hanno quindi una pressione superiore al loro interno, il sangue raccolto negli atri non potrebbe
recarsi nei ventricoli. Perciò prima che si metta in moto la muscolatura ventricolare si devono
svuotare gli atri e quindi prima che arrivi l’impulso bisognerà dare agli atri il tempo di svuotarsi.
Una volta che gli atri si sono svuotati può partire l’impulso ventricolare. Per dare questo
tempo c’è bisogno che l’impulso venga rallentato e a questo ci pensa il nodo atrio-ventricolare.
Altra cosa importante è che prima che si contragga tutta la muscolatura ventricolare devono
contrarsi i muscoli papillari, quei muscoli che tengono le corde tendinee in modo tale che quando il
muscolo cardiaco si contrae in toto, le corde tendinee siano in grado di tenere ferme le valvole
cardiache.
La prima parte del cuore che si depolarizza è l’endocardio e in particolare quello del setto
interventricolare, in seguito il resto dell’endocardio e prima ancora dell’endocardio i muscoli
papillari. Alla fine, l’intera muscolatura viene interessata da quest’onda depolarizzante e si ha la
contrazione. Questo è quello che succede normalmente.
4)CONTRATTILITÀ (inotropismo)
Proprietà delle cellule di contrarsi e di generare una forza all’interno della cavità
cardiaca,forza che poi spinge il sangue nel torrente circolatorio. L’inotropismo è quindi la capacità
di generare una forza. In fisiologia, negli esperimenti che si fanno in vitro in muscoli cardiaci
isolati, viene definita come la massima velocità di contrazione di una fibra muscolare. Negli
esperimenti si opera così: si prende la fibra e si caricano dei pesi progressivi e si va a vedere la
velocità di accorciamento sulla base del peso applicato. Più peso applico più la velocità tende ad
essere bassa. Questo tipo di esperimento, fatto già negli anni ‘50 e ’60, non si fa più e oggi per
valutare la contrattilità del cuore si usano altri parametri come la frazione di eiezione.
In medicina è più utile conoscere la derivata temporale della pressione (dP/dt massima)
all’interno della cavità cardiaca che è un parametro che ancora oggi si usa in clinica, però per
ricavarlo bisogna fare un cateterismo cardiaco. Questo parametro è una registrazione della pressione
all’interno del ventricolo di sinistra in relazione alla contrazione cardiaca. Si va a vedere la massima
velocità con cui viene sviluppata la pressione all’interno del cuore e si nota che questa pressione
cambia, questo parametro cambia, in relazione allo stato contrattile del miocardio.
In B abbiamo un soggetto iperdinamico per somministrazione di adrenalina che è una delle
sostanze che aumenta la contrattilità cardiaca, cioè aumenta la forza con cui il cuore si contrae, per
cui quando si va a fare la misurazione in questo soggetto si trova un dP/dtmax alta.
In A abbiamo un soggetto normale e in c un ipodinamico, cioè un soggetto che ha ad esempio
uno scompenso cardiaco. Questa pratica ancora si utilizza anche se ora tende ad essere sostituita da
indici indiretti di contrattilità come immagini cardiografiche, frazione di eiezione ecc.
Quello che è importante sapere, comunque, è che la contrattilità può variare, cioè la forza con cui il
mio cuore si contrae può variare in ambiti fisiologici e non necessariamente patologici. Le cause
delle variazioni della forza di contrazioni cardiache sono essenzialmente queste:
1)Incremento dell’attività simpatica,cioè una scarica di adrenalina che oltre ad aumentare la
frequenza cardiaca fa anche in modo che il cuore abbia una contrazione più forte e più efficace.
2)Aumento del precarico ventricolare. Il muscolo cardiaco come il muscolo scheletrico ha
una lunghezza ideale alla quale contraendosi ha forza maggiore, cioè se il muscolo si accorcia la
forza generata è inferiore rispetto alla lunghezza ideale. Normalmente la lunghezza ideale per le
fibre cardiache non è la lunghezza normale ma corrisponde alla lunghezza delle fibre cardiache
stirate. Infatti in questo modo le camere cardiache vengono riempite un po’ di più dal sangue e
quindi la contrazione che si avrà nel battito successivo sarà più forte. Il precarico ventricolare altro
non è che il volume del cuore alla fine della diastole quando è riempito al massimo. Più aumenta il
volume più la contrazione successiva sarà intensa e il cuore si svuoterà di più. Stessa cosa avviene
per l’aumento della frequenza cardiaca: se si tachicardizza il cuore, la forza con cui poi si contrae
aumenta.
3)Aumento del postcarico, è la resistenza che incontra il cuore quando si svuota nei sistemi
vasali. Questi vasi hanno una certa resistenza che è modulabile soprattutto dal diametro delle
arteriole di piccolo calibro che possono aprirsi o chiudersi. Se le arteriole si chiudono aumentano la
resistenza di eiezione e aumentano il postcarico ventricolare. Se si aprono, invece, diminuisce la
resistenza e diminuisce il postcarico. Se il cuore sente l’incremento del postcarico contrae le sue
fibre in maniera più efficace, con una forza maggiore.
Ci sono alcuni farmaci che tendono a ridurre la contrattilità cardiaca come ad esempio i βbloccanti (ancora molto usati), che vengono utilizzati soprattutto nella terapia dell’ipertensione e
dell'aritmia.
Caso a parte è l’acidosi, che è invece una delle maggiori cause di riduzione della contrattilità
cardiaca. Si va in acidosi quando il pH dell’organismo scende al di sotto dei normali livelli
fisiologici e può dare vari problemi, ma una delle conseguenze più drammatiche dell’acidosi sono i
problemi a livello dell’apparato cardiocircolatorio, proprio della capacità contrattile del cuore.
Nel caso della stimolazione simpatica non solo la forza è superiore ma il picco viene raggiunto
anche un po’ prima, è tutto più veloce ed efficiente. In tutti e due i grafici è visibile l’effetto del
precarico.
Nel secondo riquadro abbiamo due esperimenti in cui c’è un soggetto di controllo a cui viene
aumentato artificialmente il precarico cardiaco, cioè il volume del cuore prima che si contragga. Più
aumenta il precarico, aumentando il volume di riempimento ventricolare, più si ottiene anche un
aumento della gittata sistolica (fino ad un valore limite imposto dalla rottura della fibra).
Se poi a quest’aumento del volume telediastolico (volume presente all’interno del ventricolo
al termine della diastole) che già di per se stesso aumenta la forza di contrazione, si aggiunge anche
una stimolazione simpatica l’effetto è ancora superiore. Quindi la contrattilità cardiaca è modulabile
principalmente dal simpatico, dal volume di sangue che rientra e anche dalla resistenza che il cuore
incontra per svuotarsi.
La contrazione del muscolo avviene ad una lunghezza ideale, dove sviluppa una tensione
attiva (una tensione dovuta alla contrazione). Anche il cuore ha una lunghezza massima in cui
sviluppa una tensione attiva massima, cioè la massima capacità di contrazione, che non corrisponde
a quella di riposo ma ad una riserva di contrazione attiva, per cui anche nel cuore è possibile
costruire una curva tensione- lunghezza con una tensione ideale. Ovviamente più stiriamo più
avremo un ritorno elastico fino a quando non si romperanno le fibre.
Alessandro De Angelis
Fisiologia 22-03-12
Legenda: […..] = commenti
ECG
Nonostante le prime registrazioni elettrocardiografiche siano state fatte molto tempo fa,
rimane ancora uno degli esami fondamentali per capire se ci sono problemi cardiaci e ci dà
tantissime informazioni: informazioni sul ritmo cardiaco (es. ci consentono di vedere se ci sono
aritmie), informazioni di tipo funzionale (es. ci permettono di valutare eventuali ispessimenti del
cuore, insofferenze ischemiche); questo perché ci sono delle modificazioni particolari nell’ecg
quando si manifestano determinate patologie.
L’ecg, per tutti questi motivi e siccome è molto semplice da eseguire, continua ad essere un
esame fondamentale in cardiologia.
TRIANGOLO Di EINTHOVEN
Tutto si basa dal punto di vista teorico sul fatto che il cuore genera dei potenziali elettrici
(potenziali d’azione), e questi potenziali possono essere rilevati ponendo degli elettrodi in zone
particolari del corpo. In questo tipo di registrazione si assume che il cuore sia al centro del torace e
che occupi un’area simil triangolare chiamata: triangolo di einthoven (nome di colui che per primo
ha proposto questa teoria).
Il cuore è al centro di questo triangolo e può essere esplorato cambiando punti di vista: dai
vari apici e dai vari lati di questo triangolo; altro non si fa che registrare i potenziali cardiaci
“cambiando il punto di vista” dell’elettrodo che esplora. Gli elettrodi (indicati in figura come -/+) in
passato erano costituiti da placche metalliche mentre adesso si usano elettrodi adesivi, il significato
è lo stesso, altro non fanno che fungere da rilevatori di potenziali elettrici. Il risultato normalmente è
un’alternanza ritmica di potenziali negativi e positivi che hanno un andamento particolare.
ELETTROCARDIOGRAFIA
L’elettrocardiografia si occupa della registrazione, analisi e interpretazione dei fenomeni
elettrici,
la registrazione viene effettuata per mezzo di elettrodi posti in punti di repere specifici del
torace e degli arti. In teoria io posso mettere elettrodi ovunque anche in testa e rilevo ugualmente i
battiti cardiaci, però in realtà vanno messi in determinati punti perché:
- sono i punti in cui i potenziali elettrici sono più elevati, per cui registro i potenziali elettrici
più alti e sono meno influenzati da eventuali disturbi di altro tipo.
- perché nel tempo si sono stratificati determinati punti standard e vengono usati sempre
quelli.
Il concetto è che gli elettrodi teoricamente si possono mettere ovunque anche se poi
l’interpretazione va uniformata a quella che è l’elettrocardiografia standard.
I potenziali elettrici che vengono rilevati vengono poi scritti su carta millimetrata che scorre
ad una velocità costante, che normalmente è di 25mm al secondo. Adesso non vengono neanche più
utilizzati sistemi del genere che sfruttano lo scorrimento della carta a velocità costante, viene
acquisito tutto da sistemi computerizzati e poi stampato da una comune stampante, però sempre su
carta millimetrata.
Nell’elettrocardiografia standard, 1mm di questa carta corrisponde a 4 ms; la velocità di
scorrimento è 25 mm/s, l’ampiezza dei potenziali è tarata in modo tale per cui ad 1mV di
registrazione sulla carta ci sia una depressione positiva o negativa pari a 10 mm. Si possono
apportare delle modifiche a questi tipi di tarature, come per esempio far scorrere la carta a tratti più
velocemente, per avere delle registrazioni più precise in base alla durata di determinati eventi, ma a
questo punto non siamo più nell’elettrocardiografia standard.
DERIVAZIONI ELETTROCARDIOGRAFICHE
In questo tipo di registrazioni esistono più tipi di derivazioni elettrocardiografiche che
possiamo registrare, cioè più misure o modi di vedere il cuore.
• uno è quello delle DERIVAZIONI BIPOLARI che vengono chiamate derivazione 1°, 2° 3°
[indicate nella tabella colme derivazioni standard degli arti]
• un’ altra serie di derivazioni, più numerose, vengono chiamate DERIVAZIONI
UNIPOLARI nelle quali non esiste più un polo positivo e un polo negativo ma solo un elettrodo
esplorante. Queste derivazioni vengono chiamate derivazione AVR, AVL, AVF [indicate nella
tabella come derivazioni aumentate degli arti].
• infine abbiamo un’altra serie di registrazioni [dal libro risultano essere sempre
DERIVAZIONI UNIPOLARI] con gli elettrodi non posti più su arti ma sul torace e vengono
chiamate derivazione V1, V2, V3, V4, V5, V6. (indicate nella tabella come derivazioni
precordiali)
Il risultato è che noi abbiamo 12 tracce, e ognuna di queste tracce rappresenta il potenziale
elettrico del cuore visto da 12 punti diversi. Ci sono anche in questo caso altri tipi di derivazioni che
non sono quelle “standard” ma che rappresentano più l’eccezione che la regola quando si fa un’
elettrocardiografia. Quindi un ECG è fatto costituito da: una carta che scorre a 25mm al secondo,
c’è un’ampiezza di potenziale dove 1 mv è uguale a 10 mm e 12 derivazioni, cioè dodici punti di
vista. Questi sono i punti in cui si mettono gli elettrodi [ figura 1, 3] :
-nella derivazione 1 si mette un elettrodo nel braccio destro negativo, e uno nel braccio
sinistro e praticamente in questa derivazione l’elettrodo esplorante guarda il cuore come se fosse
posizionato in questa linea, e questa è per convenzione la derivazione che guarda il cuore a 0°.
-nella derivazione 2 l’elettrodo negativo è posto sempre nel braccio destro mentre l’altro è
posto nella gamba sinistra , quindi questa derivazione guarda il cuore con un angolo di 60°.
-la derivazione 3 ha l’elettrodo di riferimento [negativo] nel braccio di sinistra, e l’elettrodo
di registrazione nella gamba di sinistra, e guarda il cuore da 120°.
Poi abbiamo le derivazioni aumentate degli arti che hanno costanti gli elettrodi di riferimento
ma cambia la posizione dell’elettrodo di registrazione.
-AVR (elettrodo di registrazione nel braccio destro) è come se guardassi il cuore con un
angolo di -150°.
-AVL (elettrodo di registrazione nel braccio sinistro) l’elettrodo esplorante guarda il cuore
con un angolo che convenzionalmente è di -30°.
-AVF (elettrodo di registrazione nella gamba sinistra) guarda il cuore con un’angolazione di
90°.
Poi ci sono le derivazioni precordiali che si mettono sul torace.
-V1 abbiamo l’elettrodo nel quarto spazio intercostale sulla linea margino-sternale di destra.
-V2 nel quarto spazio intercostale a sinistra dello sterno.
-V3,V4,V5,V6 quinto spazio intercostale a sinistra dello sterno, prima V3 e poi V4,V5,V6
verso sinistra a distanza normalmente di qualche cm.
[ per sicurezza verificate anche voi gli angoli delle varie derivazioni..!! ]
RELAZIONE TRA EVENTI ELETTRICI DEL CUORE ED ECG
Questa è la relazione che c’è tra quello che noi registriamo e quello che sta succedendo nel
cuore, abbiamo visto precedentemente la sequenza di attivazione del cuore e che le cellule del nodo
seno atriale sono responsabili del normale ritmo cardiaco.
Quando si accende il nodo seno atriale (siamo nel primo disegno della figura) non si vede
niente perché le cellule del nodo seno atriale sono talmente poche e hanno potenziali talmente bassi
che non vengono rilevati. Si comincia a vedere qualcosa quando vengono attivati tutti gli atri, tutta
la muscolatura atriale, e voi vedete in una derivazione standard (che potrebbe essere per esempio la
derivazione 1, cioè quella che ci consente di vedere il cuore a 0°) una depressione positiva, che
viene chiamata onda P, che altro non è che la depolarizzazione degli atri. L’impulso poi, passa al
nodo atrio-ventricolare dove viene rallentato. A livello del nodo atrio-ventricolare vale lo stesso
discorso del nodo seno-atriale: noi non percepiamo niente, perché è un gruppo cellulare piccolino,
cioè registriamo una linea piatta. Dopo, l’impulso, passa ai ventricoli e normalmente nella
derivazione 1° è un impulso negativo che rappresenta le depolarizzazione del setto e dei primi
muscoli papillari. Dopo che il potenziale arriva a tutto il resto del ventricolo registreremo una
grande depressione positiva che rappresenta praticamente l’attivazione di tutta la muscolatura
ventricolare. (Alla fine, non è costante però può esserci una piccola onda data dalla
depolarizzazione della parte alta dei ventricoli). La linea ritorna sulla linea di base e inizia la
ripolarizzazione del miocardio, cioè le cellule riacquistano gli ioni che avevano perso con il primo
potenziale d’azione e si registra nella ripolarizzazione (che corrisponde alla diastole) un’onda
positiva e abbastanza duratura che si chiama onda T (successiva al tratto chiamato QRS).
LPQ RST
La prima uscita si chiama onda P, poi l’onda piccolina onda Q, l’ onda positiva grande onda
R, l’onda negativa finale onda S e infine l’ onda T.
L’onda T è seguita da un’altra piccola ondina positiva, chiamata onda U, che non siamo
ancora in grado di capire bene perché sia presente; non ha nessun significato né patologico né
fisiologico ed è presente solo in alcuni individui.
Questo è quello che normalmente vediamo con un ECG, abbiamo informazioni su
depolarizzazione atriale, depolarizzazione ventricolare e ripolarizzazione ventricolare. Mancano
informazioni sulla ripolarizzazione atriale perché la ripolarizzazione atriale normalmente avviene
quando c’è ripolarizzazione ventricolare, e quest’ultima da un punto di vista della massa delle
cellule che si depolarizzano è preponderante rispetto a quella degli atri, che quindi viene mascherata
e noi con questo tipo di registrazione sul torace non siamo in grado di registrare. Si può invece
registrare per esempio con un elettrocardiogramma endocavitario cioè mandando un elettrodo
dentro gli atri, non è una cosa che si fa frequentemente.
INFORMAZIONI CHE SI POSSONO RICAVARE DA ECG
Si va a vedere quanto durano e quanti sono alti, cioè quanti mV sono tutti questi vari segmenti
e che morfologia hanno, tutte cose il cui approfondimento richiede molto tempo; per ora dobbiamo
ricordare i concetti principali.
- La durata dell’ onda P mi dice quanto tempo impiegano gli atri a depolarizzarsi,
normalmente dura 80 ms ovvero 2 quadratini di carta millimetrata (1 quadratino = 40ms) e se ci
impiega di più abbiamo una patologia.
- L’intervallo P-R è il tempo che passa tra l’inizio dell’onda P, cioè la depolarizzazione
atriale, e l’inizio della depolarizzazione ventricolare. Ci indica quanto tempo impiega l’impulso a
passare dagli atri ai ventricoli. Normalmente questo intervallo (sono tutte misure a riposo) è al
massimo intorno ai 200 ms, 4-5 quadratini. Questo intervallo può diventare più lungo in situazioni
patologiche e indica che il sistema di conduzione ha qualche problema, cioè l’impulso ci impiega
troppo a passare dagli atri ai ventricoli e il problema può essere a vari livelli.
- L’ampiezza dell’onda P è un’altra delle cose importanti, infatti non dovrebbe mai superare
i 2 mV cioè i 2 quadratini in altezza [ 2 ?? ], perché se li supera vuol dire che gli atri sono più grandi
del normale.
- La durata totale del complesso QRS è normalmente al di sotto dei 120 ms cioè 3
quadratini, se dura di più vuol dire che ci sono problemi nella trasmissione dell’impulso all’interno
dei ventricoli, o perché qualche punto della trasmissione del impulso nervoso è interrotto e quindi
viene rallentato, oppure perché la massa ventricolare è più grande più spessa del normale per cui
l’impulso ci mette di più a viaggiare.
- Altra cosa che si può andare a vedere è quanto è ampia l’onda R, anche se qua i riferimenti
sono meno precisi per cui bisogna fare altri calcoli, che però noi per il momento non dobbiamo
conoscere.
- L’intervallo Q-T che va quindi dall’inizio della depolarizzazione ventricolare alla fine della
ripolarizzazione ventricolare. Praticamente ci dice quanto tempo impiega il ventricolo a
depolarizzarsi e ripolarizzarsi, quindi la durata di tutto il potenziale d’azione che coinvolge tutte le
cellule dei ventricoli (non gli atri); è una misura importante e non dovrebbe essere superiore ai
420/440 ms, non più di 11 quadratini. Se aumenta è una condizione pericolosa per lo scatenamento
di aritmie (esiste una malattia che si chiama “sindrome di T lungo” che predispone a morte
improvvisa per aritmie fatali)
- L’intervallo S-T non ha durata standard. Infatti per quanto riguarda il segmento S-T non è
tanto la durata ad essere importante quanto che stia sulla stessa linea della isoelettrica, cioè non
deve essere né sotto né sopra (forse il limite è non più di 1 mv sotto o sopra tale linea). Posizioni del
segmento S-T diverse sono indice di patologie cardiache (per esempio insufficienza delle arterie
coronarie).
Quindi possiamo dire che sono molte le informazioni che possiamo ricavare, facendo delle
semplici misure su un tracciato elettrocardiografico.
A COSA SONO DOVUTI I POTRENZIALI CHE REGISTRIAMO SU CARTA
MILLIMATRATA?
Sono dovuti all’insieme dei potenziali d’azione delle varie cellule cardiache.
Supponiamo di avere una fibra cardiaca e di mettere degli elettrodi: uno fuori dalla cellula
elettrodo A, uno dentro elettrodo B e un altro sempre fuori elettrodo C a distanza però
dall’elettrodo A.
-SITUAZIONE A Se noi abbiamo la cellula in questa situazione, cioè le cariche positive
all’esterno e le cariche negative all’interno, noi registriamo una isoelettrica, una linea piatta.
-SITUAZIONE B Supponiamo adesso che ad un’estremità di questa fibra ci sia un potenziale
d’azione che si propaga nella direzione indicata dalla freccia nella figura. Le polarità sono invertite,
l’interno diventerà positivo e l’esterno negativo, per cui si crea un fronte d’onda, si crea un dipolo
elettrico. L’elettrodo A e l’elettrodo B registreranno un'onda positiva (che corrisponde al potenziale
d’azione), invece gli elettrodi che si guardano all’esterno (cioè l’elettrodo B e l’elettrodo C)
registreranno un’onda che sarà simile ma che poi avrà un evoluzione un po’ più problematica.
-SITUAZIONE C Supponiamo che continui questo potenziale d’azione e che arrivi a
depolarizzare tutta la fibra, il potenziale all’interno della cellula non è più positivo ma negativo
perché le cariche si sono invertite. All’esterno avremo negativo-negativo, cioè non ci sarà più quella
differenza di potenziale tra elettrodo A ed elettrodo C e quello che registriamo è una linea
isoelettrica.
-SITUAZIONE D Inizia la fase di ripolarizzazione cellulare e tutto torna indietro al contrario,
la cellula riprende la sua polarità normale. Durante questa depolarizzazione quello che registrano gli
elettrodi esterni è un’onda negativa, l’inverso dell’onda positiva che avevamo visto prima perché in
questo caso il potenziale sarà positivo-negativo e non più come era prima.
-SITUAZIONE E Alla fine quando la fibra si sarà completamente ripolarizzata noi
registreremo nuovamente una linea isoelettrica cioè uno zero.
Questo era quello che succede in una fibra, mentre ora vedremo quello che succede in uno
schema di tutto il cuore
Il VETTORE ELETTRICO MEDIO CARDIACO
Il cuore è formato da molte fibre per cui quello che noi andiamo a registrare è il risultato di
una serie di vettori elettrici, che avranno diverse direzioni. In una piccola porzione di cuore ci
saranno una serie di fibre che hanno un vettore orientato in un senso; in un’altra porzione, invece,
fibre con un vettore orientato in un’altra direzione; e così via. Quindi quello che noi andiamo
registrare con l’ECG sarà il vettore risultante cioè la somma [vettoriale] di tutti i vettori. Il concetto
da ricordare è che l’ ECG non è altro che la registrazione del vettore elettrico medio cardiaco in
un determinato istante e che l’evoluzione temporale di questo vettore da l’ ECG (cioè quel
tracciato che noi vediamo sulla carta millimetrata).
Evoluzione spazio-temporale del vettore elettrico medio cardiaco in relazione al ECG
Nella realtà ciò che succede è l’evoluzione nello spazio e nel tempo del vettore elettrico
cardiaco medio in relazione all'ECG.
- osservando il tracciato PQRST e il cuore, possiamo notare che all’inizio della
depolarizzazione degli atri il vettore è orientato in questo senso (il vettore medio, cioè la somma di
tutti i vettori che in quel momento “stanno avvenendo” negli atri).
- l’impulso passa al sistema di conduzione, al nodo atrioventricolare, e non succede niente
infatti siamo sulla isoelettrica.
- il vettore arriva al sistema di conduzione e al setto. Quindi ricominciamo a vedere dei
potenziali che in questo caso sono negativi perché il vettore si è spostato e non è più come era
prima, è orientato invece verso l’alto e un po’ anche posteriormente (teniamo presente che stiamo
osservando il cuore dalla derivazione 1°, quindi a 0°)
- avremo poi il complesso QRS (nell'ECG) , cioè il complesso di depolarizzazione di tutto il
ventricolo, con un grosso vettore (rappresentato da quest’onda R) orientato più o meno com’è
orientato il vettore di depolarizzazione atriale, cioè verso il basso e verso la punta del cuore. Il
vettore cambia poi direzione e si orienta verso gli atri, fino a ritornare nuovamente sulla linea
isoelettrica (il vettore ritorna a 0)
Quindi riepilogando quello che noi registriamo è l’evoluzione temporale di un vettore
elettrico. [nella figura il vettore dovrebbe essere una freccia bianca che però si vede poco]
Sempre nella stessa immagine oltre alla traccia elettrocardiografica possiamo notare “dei
segni strani azzurri” che rappresentano un altro tipo di registrazione chiamata vettocardiografia,
cioè la registrazione del vettore cardiaco nelle tre dimensioni con dei sistemi abbastanza complessi
(per adesso ci accontenteremo di conoscere l’ elettrocardiografia).
IL RISULTATO FINALE
Come risultato finale noi avremo 12 tracce, le 12 derivazioni di cui avevamo parlato prima
(per adesso abbiamo visto nel dettaglio soltanto la derivazione 1°). Ognuna delle 12 derivazioni ha
un tracciato (un’evoluzione) particolare, diversi tra loro proprio perché guardano il cuore da diversi
punti di vista (per esempio la dervazione V1 è molto diversa dalla derivazione V6 proprio perché
una guarda il cuore dal davanti e un po’ da destra, l’altra dal lato e a sinistra, infatti i vettori sono
completamente diversi).
Dall'analisi di tutte queste 12 tracce siamo in grado di dare giudizi diagnostici anche
abbastanza precisi su molti tipi di patologie cardiache.
[ Domanda: per quale motivo nel ECG depolarizzazione e ripolarizzazione hanno lo stesso
senso, cioè sono tutte e due positive?
Risposta: Perché normalmente le depolarizzazione va dall'endocardio all'epicardio, dagli strati
più interni del cuore agli strati più esterni, mentre la ripolarizazzione segue il percorso inverso,
dall'epicardio all’endocardio. Nel cuore in toto quindi, le zone che per prime si sono depolarizzate
si ripolarizzano per ultime. Questo avviene perché le cellule endocardiche “soffrono di più” per la
pressione all’interno della cavità cardiaca, per cui hanno bisogno di più tempo per recuperare gli
ioni; le cellule epicardiche “soffrono di meno” per la pressione all’interno del cuore e sono più
veloci nella ripolarizzazione].
LA MECCANICA CARDIACA
Noi nella figura abbiamo schematicamente il ventricolo di sinistra simile ad un tronco di
cono con una sezione più o meno circolare. Il ventricolo di destra invece forma una sorta di tasca
che sta attorno al ventricolo di sinistra e sulla destra, con una cavità che non è proprio circolare. La
parete del ventricolo di destra è notevolmente più sottile di quella del ventricolo di sinistra. I due
ventricoli hanno modi di contrarsi leggermente diversi.
Gran parte della contrazione del ventricolo di destra si compie attraverso un accorciamento
della superficie laterale e attraverso una compressione verso il ventricolo di sinistra che “gli fa da
àncora” (cioè come se il ventricolo di sinistra fosse una parete ferma che attraverso il setto
interventricolare funge da àncora per tutto il ventricolo di destra, che così si schiaccia e si accorcia).
La contrazione del ventricolo di sinistra è diversa; molto lo fa attraverso la riduzione degli
atri, una letterale compressione del volume che c’è all’interno degli stessi. Anche il ventricolo di
sinistra ha una certa capacità di accorciarsi, di accorciare il suo diametro longitudinale, con un
movimento che va a comprimere il sangue che ha all’interno.
Questi movimenti determinano l’espulsione di una parte del volume di sangue che c’è
all’interno dei ventricoli stessi, che però non corrisponde alla quantità totale presente, ma è molto
inferiore. Quella appena descritta è la
frazione di eiezione: la quantità di sangue espulsa dal cuore rispetto alla quantità totale
presente in diastole.
Es. supponiamo che il ventricolo di sinistra abbia 100 ml di sangue al suo interno, alla fine di
una sistola normalmente di questi 100 ml ne rimangono 40ml/45ml, quindi la frazione di eiezione,
la quantità totale di sangue espulso, è intorno al 55-60% della quantità totale presente nel
ventricolo (che era arrivata dagli atri). Un buon 40% di sangue rimane all’interno del ventricolo e
non viene espulso. Tutto questo sempre in condizioni di riposo perché durante l’esercizio la % può
cambiare. La frazione di eiezione del ventricolo di sinistra è intorno al 55-60%, quella del
ventricolo di destra normalmente è un po’ più bassa, intorno al 50%, perché la capacità di
contrazione del ventricolo di destra è un po’ inferiore rispetto al ventricolo di sinistra (proprio per lo
spessore della parete).
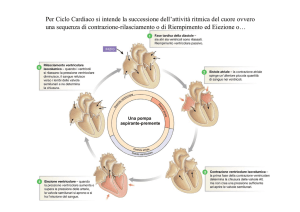
IL CICLO CARDIACO
Il ciclo cardiaco è l’insieme di movimenti, di azioni compiute dal cuore. Essenzialmente è
costituito dall’alternanza di 2 FASI, una di sistole e una di diastole, la cui durata totale dipende
dalla frequenza cardiaca e normalmente è di circa 0,8 / 0,9s (in una persona quindi con frequenza
cardiaca di 70/75 bpm) può arrivare ad 1 s (se la frequenza cardiaca è di 60 bpm). Oltre alla
suddivisione in sistole e diastole all’interno del ciclo cardiaco noi possiamo individuare almeno
altre 5 sub-periodi (alcuni arrivano ad individuarne anche 7) che hanno un significato ben preciso e
che ora vedremo. Abbiamo quindi una fase di sistole e una di diastole.
Nella SISTOLE [figura-lettera a] possiamo definire almeno altri due momenti, che sono
quelli di contrazione ventricolare isovolumetrica e quello di eiezione ventricolare.
- Contrazione ventricolare isovolumetrica. Durante questa fase i ventricoli si contraggono e
sia per la morfologia del vaso che per la trazione dei muscoli papillari, le valvole atrio-ventricolari
si chiudono e impediscono al sangue di tornare verso gli atri. Nei primi momenti della fase di sistole
però la pressione ventricolare (15/20 mmHg) non è tale da vincere la pressione (90 mmHg) che si
trova all’interno delle grande arterie (arteria polmonare e arteria aorta). Quindi nella prima fase
della sistole, il cuore si contrae senza variazioni volumetriche, cioè si contrae per generare quella
forza che gli servirà poi per aprire le valvole. C’è infatti un aumento di pressione senza che ci sia
una variazione di volume (P e V che noi possiamo misurare mandando dentro il cuore uno
strumento per misurare la pressione e uno per misurare il volume).
- Eiezione ventricolare. La pressione aumenta progressivamente fin quando non raggiunge e
poi supera quella delle grande arterie, per cui la valvola polmonare e la valvola aortica si aprono e
consentono il flusso di sangue verso il piccolo e il grande circolo. Mentre ciò accade ovviamente la
valvola mitrale e la valvola tricuspide continuano a rimanere chiuse per l’azione pressoria su i loro
lembi generata dalla contrazione ventricolare.
Finita la fase di contrazione, esaurita la spinta, il ventricolo si rilassa e inizia la fase di
DIASTOLE [figura-lettera b].
- Rilassamento ventricolare isovolumetrico. Durante questa prima fase di diastole la
pressione all’interno dei ventricoli è più bassa di quella delle arterie per cui le valvole a nido di
rondine si chiudono. La pressione all’interno dei ventricoli però è ancora troppo alta rispetto a
quella degli atri, per cui il ventricolo si rilascia ma senza variazione di volume, creando una
situazione simile alla situazione precedente [contrazione ventricolare isovolumetrica], con la
differenza che prima si stava contraendo senza produrre variazioni di volume invece ora si sta
rilassando senza produrre variazioni di volume. Quindi non c’è un flusso di sangue netto né verso le
arterie né dagli atri. La pressione continua a scendere, fino a quando la pressione che c’è all’interno
degli atri non avrà uguagliato e poi superato quella che c’è all’interno dei ventricoli [momento che
rappresenta l’inizio della fase successiva].
- Riempimento ventricolare. Quando la pressione all’interno degli atri è maggiore di quella
dei ventricoli, le due valvole atrio-ventricolari si aprono, una grande quantità di sangue
passa dagli atri ai ventricoli, gli atri si sgonfiano dentro i ventricoli per gradiente di
pressione (perché quando il ventricolo è completamente rilasciato la pressione ventricolare è
circa 0 mmHg mentre negli atri 5/6 mmHg); questa è la prima fase quindi che segue quella
del rilassamento ventricolare e viene chiamata fase di riempimento ventricolare rapido [in
figura è indicato l’atrio rilassato]. In un cuore normale, giovane, gran parte del riempimento
cardiaco in diastole avviene durante questa fase, il 70-80% di sangue che deve riempire il
cuore lo fa durante la fase di riempimento ventricolare rapido. Alla fine di questa fase di
diastole abbiamo la contrazione atriale (riempimento ventricolare lento) attraverso la
quale l’atrio immette nel ventricolo la parte rimanente di sangue che vi era negli atri, cioè
dà un’ulteriore spinta al sangue verso i ventricoli. La contrazione atriale in un cuore
normale, giovane, può anche mancare e non succede niente perché come abbiamo già detto
gran parte del riempimento il cuore lo effettua nella fase precedente, mentre può diventare
importante in stati patologici. La contrazione atriale è responsabile di non più del 20% del
riempimento cardiaco.
VARIAZIONE PRESSORIE E VOLUMETRICHE DURANTE UN CICLO CARDIACO
Nella figura sono rappresentati i 5 periodi del ciclo cardiaco visti prima: riempimento
ventricolare (lento), contrazione ventricolare isovolumetrica, eiezione ventricolare, rilassamento
isovolumetrico e riempimento ventricolare rapido.
Se noi mandiamo dentro il ventricolo un misuratore di pressione (pressione ventricolare
indicata nel grafico con una linea rossa) e un altro misuratore di pressione dentro l’atrio (pressione
atriale indicata nel grafico con un linea blu) registreremo le due curve che vediamo nel grafico.
Inoltre contemporaneamente possiamo anche registrare la pressione aortica (tramite “il bracciale”),
il volume nel ventricolo di sinistra, l’ ECG e i toni cardiaci.
-La pressione nell’aorta. Osservando il comportamento della pressione nell’aorta noi
vedremo che durante le diastole la pressione tende progressivamente a scendere. Praticamente si
perde energia nel sistema e se non ci fosse il ciclo cardiaco andrebbe a zero (la morte per
intenderci). Questo fino a quando non inizia la fase di eiezione ventricolare, in cui la pressione
all’interno dei ventricoli supera quella che c’è all’interno dell’arteria e il sangue viene eiettato in
aorta con l’andamento che vediamo nel grafico, un tratto ad ascendere piccolo e un tratto a
discendere quando il ventricolo ha esaurito la sua spinta. La curva della pressione arteriosa continua
a scendere fino ad arrivare in un punto in cui forma una sorta di piccolo dente, chiamato incisura
dicrota, che individua il momento in cui la valvola aortica si chiude e il sangue fa una sorta di
rimbalzo elastico sulle pareti. [il professore indica nel grafico i punti in cui la valvola aortica è
aperta e chiusa]. Quando la valvola aortica è chiusa non consente il passaggio del sangue dal
ventricolo verso l’aorta perché a livello aortico la pressione è più alta che dentro il ventricolo. La
durata dell’ eiezione, ponendo che la durata totale del ciclo cardiaco sia di 1s, è normalmente
intorno ai 250-300ms, quindi non più di 1/3 del ciclo cardiaco totale.
-La pressione nel ventricolo di sinistra. Noi abbiamo una prima fase in cui il ventricolo non
si contrae, abbiamo un aumento della pressione all’interno del ventricolo dovuto al fatto che l’atrio
si sgonfia, che è praticamente l’onda di pressione atriale che si trasmette dentro il ventricolo. Dopo,
inizia a contrarsi anche il ventricolo e abbiamo la contrazione isovolumetrica [indicata nel grafico
con il numero 7]. La pressione dentro il ventricolo inizia a salire, fin quando non diventa uguale a
quella che c’è nell’aorta (inizia l’eiezione ventricolare). Continuando il ciclo cardiaco il ventricolo
continua a contrarsi e spremersi, la pressione all’interno sale fino a corrispondere a quella che
troviamo in aorta. Poi il ventricolo si rilascia, finisce la contrazione, la pressione bruscamente
scende e va a 0; inizia la fase di diastole ventricolare con il riempimento ventricolare che è
rappresentato dal tratto della curva che ascende, con la pressione che progressivamente sale.
-La pressione nell'atrio di sinistra. La curva della pressione atriale è qualcosa di molto
simile a quella della pressione ventricolare, solo che manca tutta la fase eiettiva perché l’atrio non
partecipa all’eiezione del sangue nell’arteria.
-Volume nel ventricolo di sinistra. La quantità di sangue presente nel ventricolo raggiunge il
massimo subito dopo la contrazione atriale, poi inizia la contrazione ventricolare, e quando la
valvola aortica si apre, il volume bruscamente scende perché il cuore si sta svuotando. Il volume
telediastolico indicato nel grafico rappresenta la quantità di sangue presente nel cuore prima della
sua contrazione, ha una grande importanza clinica (importanti variazioni del valore normale si
hanno in quelle patologie dove il cuore tende ad essere più grosso del normale). Il cuore si svuota
fino a raggiungere il massimo dello svuotamento quando la valvola aortica si chiude per caduta
pressoria all’interno del ventricolo e inizia il riempimento ventricolare (prima c’è il rilassamento
ventricolare isovolumetrico, poi il riempimento ventricolare rapido e il riempimento ventricolare
lento con contrazione atriale).
Sotto i grafici visti fin ora è presente anche l’ ECG ed è interessante notare come l’evento
elettrico, (in particolar modo il QRS) preceda l’evento meccanico, cioè come la depolarizzazione
preceda la contrazione.
Nel grafico sopra vediamo che “nel cuore di destra” avviene qualcosa di molto simile a ciò
che avviene “nel cuore di sinistra”, però in scala ridotta. Mentre la pressione media all’interno
dell’aorta è intorno ai 100 – 120 mmHg, nell’arteria polmonare è molto più bassa 20 - 25 mmHg;
quello che però succede è più o meno la stessa cosa.
TONI CARDIACI
[in questo grafico a destra] c’è una retta con dei segni gialli che rappresentano i toni cardiaci
[visibili anche nella figura a sinistra e nel fonocardiogramma in basso].
1°tono [tà] è il suono prodotto dalla chiusura quasi sincrona delle valvole atrioventricolari.
All’interno del primo tono possiamo individuare almeno 4 picchi corrispondenti a fasi diverse della
chiusura delle due valvole atrioventricolari, però i picchi più importanti sono il primo e il secondo
(cioè i rumori più forti) prodotti dalla vibrazione delle valvole atrioventricolari durante la chiusura.
2°tono [tùm] è il suono prodotto dalla chiusura quasi sincrona della valvola aortica e
polmonare e dura un po’ di più rispetto al 1° tono. [non sono presenti tutti i picchi che ci sono nel
primo tono]
Quindi quello che si va ad ascoltare quando si fa l’auscultazione cardiaca sono questi due
rumori, che devono essere puri (cioè non devono esserci altri rumori nel mezzo perché possono
avere significato patologico), e devono avere un determinato tono e una determinata durata.
Osservando il grafico che analizza più nel dettaglio il fonocardiogramma
se noi registriamo contemporaneamente il polso carotideo (prendiamo la carotide e ci
mettiamo sopra un trasduttore di pressione) e i rumori cardiaci, troviamo un perfetto sincronismo tra
l’ascesa di questo polso e il primo tono cardiaco (chiusura delle valvole atrioventricolari) e un altro
sincronismo tra l’incisura dicrota (il rimbalzo di cui parlavamo prima) e il secondo tono cardiaco.
Quelli indicati con 3 e 4 rappresentano il 3°tono e 4°tono, sono chiamati toni aggiunti e spesso
hanno significato patologico. Il 3° tono è dovuto alle vibrazioni del ventricolo che si riempie, è un
tono presente in situazioni patologiche come cuori rigidi o ischemici ma anche nei cuori sani di
alcuni atleti. Il 4° tono invece è SEMPRE espressione di patologia
Analizzando il POLSO GIUGULARE (si usa la vena giugulare perché è la vena più
semplice da usare dal punto di vista anatomico, ma un tracciato simile lo ritroviamo più o meno in
tutte le vene) cioè analizzando le variazioni pressorie all’interno della vena giugulare troviamo dei
picchi positivi chiamati onde A, onde C, onde V e dei picchi negativi chiamati onda X e onda Y .
Onda A, è chiamata cosi perché rappresenta l’onda trasmessa dalla contrazione atriale, cioè
quando l’atrio si contrae trasmette l’onda pressoria non solo ai ventricoli ma a tutto il sistema
venoso e più la vena considerata è vicina al cuore più l’onda a si vede (come nel caso della
giugulare).
Onda C, è l’onda di contrazione ventricolare trasmessa all’albero circolatorio che può essere
definita anche come un’onda retrograda meccanica che si trasmette a tutto l’albero circolatorio.
Onda X. Nel momento in cui la valvola aortica si apre, la pressione scende bruscamente
all’interno del sistema venoso. Perché nel momento in cui il ventricolo si contrae non solo pompa
sangue nell’arteria polmonare e nell'aorta, ma genera anche un’azione di traino sulle valvole
atrioventricolari. Il setto (una delle prime parti del ventricolo a contrarsi) infatti, contraendosi, porta
letteralmente giù le valvole atrioventricolari che così “risucchieranno” il sangue dalle vene verso gli
atri. Questa appena descritta è proprio una forza che si chiama vis a tergo che il cuore esercita sulle
valvole atrioventricolari, che poi a sua volta si trasmette su tutto il sistema venoso e che quindi
favorisce il ritorno di sangue al cuore (il cui effetto è visibile nel grafico come un brusco calo di
pressione).
Onda V. La pressione pian piano risale e c’è un punto di massima pressione quando le valvole
atrioventricolari sono ancora chiuse ed è chiusa anche la valvola aortica.
Onda Y. Appena si apre la valvola atrioventricolare il sangue fluisce verso il ventricolo e
quindi la pressione si riabbassa nuovamente al livello del sistema venoso.
Quindi anche nel torrente circolatorio venoso abbiamo un ciclo continuo di picchi e
depressioni che determina un movimento di sangue verso il cuore.
Fisiologia 24 Marzo 2012
Prof. Crisafulli
Sabrin
"Ha fatto vedere diversi video ecografici. Durante la presentazione ha indicato i ventricoli, gli
atri e le valvole. Hanno fatto da introduzione alla lezione. Di seguito trovate ciò che ha detto. Per
non appesantire il discorso cercherò di trascrivere ciò che si può capire anche senza vedere il
video."
Esiste una fase in cui le valvole si aprono. E un momento in cui entrambe le valvole si
chiudono, questo accade durante la sistole, nello specifico durante la contrazione isovolumica .
"Poi ha fatto vedere i flussi dall' atrio sx al ventricolo sx e dal ventricolo sx all'aorta, visti
sotto l'effetto Doppler."
Il Doppler registra i flussi e li codifica come variazione di colore. Tuo ciò che è rosso è un
qualcosa che si avvicina alla sonda, tutto ciò che compare in blu è un qualcosa che si allontana dalla
sonda. I due flussi sono asincroni. Possiamo individuare un flusso di ingresso e un flusso di uscita.
Il flusso blu è in uscita, siamo in diastole; il flusso rosso è in entrata, vuol dire che siamo in
sistole ( mentre dice questo indica il ventricolo sx ). L'intervallo tra una fase e l'altra è dell'ordine
di 10-15 ms. Nei soggetti con una frequenza bassa, queste fasi hanno un ritmo lento e quindi più
apprezzabili. Con uno sguardo più attento osserviamo anche la contrazione degli atri, oltre che del
ventricolo.
"Poi in un altro video si è soffermato su quesi due punti":
•
sull'attaccatura della valvola tricuspide che normalmente è un po’ più bassa rispetto
alla valvola mitrale.
•
Sul trascinamento del setto interventricolare nei confronti degli atri, tira giù lo
scheletro fibroso del cuore. Durante la sistole il setto si contrae e stira gli atri, aumentandone
il volume! La contrazione ventricolare non serve solo per pompare il sangue dal ventricolo,
ma anche per favorire e migliorare il riempimento degli atri!
"In un altro video ha sottolineato il ruolo dei muscoli papillari, come apparato di ancoraggio
per le valvole atrio-ventricolari." Nel caso in cui questo meccanismo non funziona parliamo di
incontinenza ( non sono sicura che abbia usato questo termine) e avremo passaggio di sangue dal
ventricolo all'atrio e si creano flussi, è una situazione abbastanza comune e si chiama prolasso della
valvola mitrale.
Fine di rassegna video, che hanno fatto da introduzione alla lezione
-----------------------------------------------------------------------------------------------------------------------------------------------Ritorno venoso:
"Per poter pompare sangue nel torrente
circolatorio è necessario il cuore riceva
continuamente sangue dal sistema venoso.
Diversi meccanismi garantiscono il ritorno
venoso ed il pre-carico cardiaco" ( tratto dalla slide)
Il cuore viene visto come una vera e propria pompa del sistema vascolare . C'è tutto un
sistema che fa sì, che il sangue ritorni al cuore con un certo gradiente di pressione, cosicché il cuore
possa svolgere la sua funzione di pompa, non a vuoto. La pressione che il cuore imprime al sangue
tende a cadere nell'albero circolatorio. Tuttavia a livello del sistema venoso ho sempre una
pressione maggiore rispetto a quella che trovo nell'atrio di dx, perchè il sangue arriva con un
minimo di energia ( la quale risale all'energia che il sangue ha ricevuto al momento dell'eiezione);
ciò consente il ritorno venoso. Ciò è molto importante, almeno a riposo.
Il gradiente di pressione viene creato e mantenuto attraverso:
1. Vis a tergo --> ossia azione di pompa del cuore. Questo fa in modo che durante la sistole,
all'interno dell'atrio stesso si crei un calo di depressione che risucchi il sangue dal sistema venoso.
Ciò è dovuto anche al trascinamento degli atri ,dovuto alla contrazione dei ventricoli e del setto
interventricolare, fenomeno noto come" effetto risucchio".
2. Pompa muscolare --> crea una pressione sui vasi ad esempio della gamba ,dovuta alla
contrazione dei muscoli della gamba e alla disposizione delle vene tra le fibre muscolari, così il
flusso viene in tal modo facilitato. Le valvole permettono l'unidirezionalità del movimento del
sangue,le quali sono a nido di rondine. La direzione va così dalla periferia al cuore. Se questo
sistema non funziona avremo stasi, ciò accade in alcune patologie. E' fondamentale durante
l'esercizio (non necessariamente durante un'attività fisica, anche il semplice camminare è
considerato un esercizio), affinché al cuore arrivi tutto il sangue che ha pompato. Altrimenti il cuore
avrà una gittata sistolica inferiore rispetto allo stato di riposo.
3. Spremitura simpatica delle vene --> dovuta all'azione del sistema nervoso autonomo, in
particolare dell'ortosimpatico , il quale spreme letteralmente le vene. Avviene durante l'esercizio
fisico (il professore a fine corso ci terrà un seminario sul rapporto attività fisica e sistema
cardiovascolare). In generale, durante l'attività fisica, l'attività simpatica aumenta, la quale
vasocostringe, aumenta l'attività cardiaca, cose che abbiamo visto con la pressione arteriosa.
Il 60% del sangue si trova nelle vene, le quali fanno quindi da serbatoio di sangue di
emergenza, con la semplice spremitura del Sistema Nervoso simpatico, abbiamo mobilizzazione di
questo volume di sangue, ad esempio in uno stato di emorragia
4. pompa respiratoria---> durante l'inspirazione abbiamo
l'aumento della pressione nelle
l'abbassamento del diaframma, quindi l'aumento di volume della vene, causa un aumento del ritorno
gabbia toracica, accompagnata dal calo di pressione nella
venoso , aumento del volume del sangue
gabbia toracica. Questo decremento pressorio provoca un
nell'atrio, quindi aumento della pressione
richiamo di sangue dalla periferia verso il torace e quindi
atriale, quindi aumento del volume
garantisce il ritorno venoso. Nell'espirazione forzata, ho una
ventricolare telediastolico, cioè aumento
costrizione del torace, abbiamo spremitura delle vene, vena cava della quantità di sangue nel cuore alla
superiore e inferiore, ciò può dare problemi a pazienti con
fine della diastole. Se il cuore funziona
bene, risponde aumentando la gittata
scompenso cardiaco.
L'aumento della pressione nelle vene lo posso avere con il sistolica, ossia pompa tutto quel sangue
semplice aumento del volume di sangue circolante. Per esempio che gli arriva con più forza. Ciò può
essere fisiologico, ma anche patologico.
quando la VFG diminuisce (indice di insufficienza renale),
Fisiologico quando è dovuto alla pompa
l'H2O nel sistema venoso aumenta, perché il riassorbimento
dell'acqua aumenta, quindi anche il volume di sangue aumenta, muscolare durante l'esercizio fisico.
Patologico nel caso, ad esempio, di
quindi più sangue arriva al cuore. Praticamente avrò lo
discompenso cardiaco congestizio
scompenso cardiaco congestizio. Il cuore riceve più sangue di
quanto ne può pompare, il cuore si gonfia letteralmente.
.
Gittata cardiaca e emodinamica
Quando parliamo di cuore le principali grandezze prese in considerazione sia in fisiologia, che
in clinica sono:
• Frequenza cardiaca
• Gettata sistolica (ml)
• Gettata Cardiaca (l/min)
• Volume di fine diastole (pre-carico) (ml)---> VFD
• Volume di fine sistole (ml)--->VFS
• Frazione di eiezione (%)
La frequenza cardiaca è il numero di battiti al secondo.
La gittata cardiaca è la quantità di sangue espulsa dal cuore in ogni singola sistole.
La gittata cardiaca altro non è che la moltiplicazione della frequenza cardiaca per la gittata
sistolica, quindi la quantità di sangue espulsa dal cuore in un minuto.
Volume di fine diastole è la quantità di sangue presente dentro il cuore alla fine della diastole,
poco prima che il cuore si contragga, viene spesso indicata come precarico cardiaco e misurato in
ml, ha il valore di 120-130 ml in una persona normale.
Il precarico ventricolare è di 120 ml nel ventricolo di sx, questa quantità va moltiplicata per il
volume del ventricolo di dx, che ne contiene altrettanto, poi moltiplichiamo per il volume di sangue
contenuto negli atri. Si prende come riferimento il precarico del ventricolo di sx.
Volume di fine sistole è la differenza tra il volume di fine diastole e la gittata sistolica. Cioè è
la quantità di angue che rimane dopo la gittata sistolica
Frazione di eiezione è la quantità di gittata sistolica espulsa rispetto al volume di fine
diastole , si esprime in %. Se ho 100 mL nel ventricolo di sx e ne ho eiettati 60 ml. Significa che il
volume di eiezione è del 60%.
Ci sono una serie di sistemi per misurarli, alcuni vengo usati più di altri. Alcuni sono invasivi,
altri no, Alcuni sono pi semplici rispetto ad altri.
Metodi per la determinazione della gittata cardiaca
• Principio di Fick
• Termodiluizione e diluizione del colorante
• Scintigrafia miocardica
• Ecocardiografia e Doppler
• Rebreathing del CO2 e dell C2H2
• Cardiometria ad impedenza
• Pletismografia
Il principio di Fick è semplice, lo vedremo più in là, con il polmone. E' un principio molto
usato in fisiologia. Non vedremo tutti questi metodi, alcuni ve li accenno giusto per sapere che
esistono.
La termodiluizione e la diluizione del colorante sono metodi che vengono usati
essenzialmente quando si fa cateterismo cardiaco. Il cateterismo cardiaco, è un metodo invasivo
serve per analizzare il cuore con delle sonde, arteriose e venose, sia al livello degli atri , che dei
ventricoli, si fanno dei campioni e si studiano diversi parametri del cuore, come la pressione
intraventricolare, la saturazione dell' O2 del sangue nei ventricoli e degli atri. La termodiluizione
consiste nell'iniettare nel cuore del sangue pre-riscaldato o dei coloranti veri. E si va a vedere come
queste sostanze si diluiscono nel sangue generale, serve per calcolare quanti volumi di sangue
posseggo. Se conosco il volume del liquido iniettato e vedo quanto si è diluito, posso sapere in che
volume di sangue si è diluito, con dei semplici calcoli.
Scintigrafia miocardica consiste nell'iniettare nel torrente circolatorio sostanze radioattive,
marcando radioattivamente gli eritrociti, poi si misura la radioattività all'interno del cuore con un
rilevatore di radioattività, lo si pone a livello del torace. Conoscendo la quantità di sangue
radioattivo iniettato e la quantità di liquido in cui si è diluito, posso risalire alla quantità di sangue
nel cuore.
Quindi conoscendo la radioattività iniettata siamo in grado di calcola la gittata sistolica,
VFD,VFS ecc
Ecocardiografia attraverso l'Effetto Doppler ci dà la possibilità di misurare tutti quei
parametri gittata sistolica, gittata cardiaca,VFD,VFS ecc E' un esame di routine, perchè non
invasivo e non ha alcuna pericolosità. E' stato introdotto da circa 20 anni.
Per motivi di ecogenicità del cuore, quindi dello stato di salute del cuore, non si può fare su
tutti i pazienti, perché un conto è applicarlo su un atleta, un conto è usarlo su un paziente con un
cuore ricoperto da uno strato di grasso.
"Di seguito sono elencati i metodi più usati in fisiologia", meno invasivi anch'essi
Rebreathing della CO2 e del C2H 2 ( Acetilene ) iniettando queste sostanze nel torace e
sulla base delle variazioni della corrente di queste sostanze, sincrone con il torrente circolatorio
siamo in grado di estrapolare la gittata sistolica e così la gittata cardiaca. Non tanto i VFD e VFS.
Pletismografia sulla base dell'onda sfigma, cioè dell'onda pressoria che arriva in periferia e
usando un determinato algoritmo è possibile alla gittata sistolica in maniera indiretta.
Adesso vedremo il principio di Fick, il quale ci dà una stima della gittata cardiaca.
Con questo principio siamo in grado di misurare qualsiasi flusso (anche di gas), purché ci sia
una sostanza che passando attraverso un organo subisca una variazione di concentrazione. Nel
nostro caso il flusso che ci interessa è quello del plasma.
PAx e PVx sono rispettivamente le concentrazioni della sostanza che stiamo studiando nel
sangue arterioso e venoso, cioè il flusso di entrata e uscita dal circuito dell'organo considerato, in
questo caso il cuore. L'equazione di Fick si applica in 2 casi, o quando la sostanza viene
completamente eliminata dal circuito o quando si sa a priori qual è la percentuale di sostanza che
viene eliminata. Se viene eliminata in parte devo conoscere la percentuale con cui viene eliminata.
La formula è questa:
F= Qx/PAx-PVx
dove F= flusso, Qx= quantità di sostanza che scompare (o compare) dal
plasma, PAx e PVx= concentrazione della sostanza nel sangue arterioso
e venoso rispettivamente.
Per calcolare il flusso bisogna conoscere la differenza artero-venoso della sostanza presa in
considerazione, ne devo conoscere anche la quantità usata.
La sostanza usata nel prossimo esempio è l'O2, la quale si presta bene al principio di Fick.
L'O2 nel circuito, (applicato al sistema circolatorio. Partendo ovviamente dal cuore e andando in
periferia), preso in esame scompare, invece la CO2 è una sostanza che compare, applichiamo il
ragionamento inverso. Quindi Qx dipende dalla sostanza presa in considerazione. Parliamo di
consumo di O2 nel caso in cui usiamo l'ossigeno.
"Digressione": quando in periferia arriva il sangue ai tessuti, il flusso di sangue in uscita dai
tessuti conterrà meno ossigeno di quanto ne aveva prima il flusso in entrata. Perchè una certa
quantità di ossigeno è stata consumata, quindi sarà scomparsa nel passaggio artero-venoso, ciò
viene indicato con VO2. E viene chiamato per l'appunto consumo di ossigeno!
Per calcolare la differenza di pressione artero-venosa dell'ossigeno in concentrazione prendo
in considerazione i seguenti parametri.
Visto che a noi serve calcolare la gittata cardiaca (GC), riscriviamo la formula nel seguente
modo, ponendo :
GC= QX/Ca-Cv ---> otteniamo la quantità di sangue pompato in un minuto
Per calcolare la differenza di pressione artero-venosa dell'ossigeno in concentrazione prendo
in considerazione i seguenti parametri.
In media la concentrazione dell'ossigeno nel sangue arterioso è 190 ml per litro, significa che
per ogni litro di sangue mandato ai tessuti questo contiene 190 ml di ossigeno. Invece il sangue
venoso, refluo, nello stato di riposo, ha una concentrazione di ossigeno è pari a 140 mL per litro di
sangue, in gran parte legato all'Hb, i l resto è disciolto. Con questi dati posso calcolare la differenza
artero-venosa dell'ossigeno che sarà 50mL per litro di sangue passato nei tessuti.
Il consumo di ossigeno lo ottengo attraverso il metabolimetro. Ed è soggettivo da individuo a
individuo.
La concentrazione plasmatica dell'ossigeno nel sangue venoso e nel sangue arterioso li
ottengo attraverso un prelievo venoso ed arterioso.
La gittata cardiaca sarà uguale a 250 mL/min. Se lavoro con i volumi, ottenuti con
l'ecocardiogramma, posso risalire al volume di fine diastole e il volume di fine diastole, facendo le
differenze ottengo la GC.
Risalgo alla gittata sistolica dividendo per la frequenza cardiaca, misurando il polso radiale. Il
princio di Fick viene usato dagli anastesisti, per controllare di continuo la GC. Usando una canula
venosa ed una canula arteriosa.
Equazioni utili
• GC= GSxFC; GS= GC/FC; GS= VO2/AVO
2diff./FC;
• GS= VFD-VFS; GC= (VFD-VFS)xFC
• GS= VFDxEF; GC= (VFDxEF)xFC
• EF= (GS/VFD)x100
Conviene conoscerle, soprattutto per chi vorrà fare cardiologia.
Valori normali di GC e GS a riposo
• GC= 4-7 L/min; IC> 2.5 mL/m2
Dipende molto dalla taglia del soggetto, più uno è grosso, più ha bisogno di nutrite i tessuti.
Più il cuore è grande, più fortemente pomperà il sangue. 4 l di GC vanno bene per una persona che
pesa 50 Kg, 7 l invece per una persona che pesa 100 Kg. Spesso la gittata cardiaca tiene conto della
superficie corporea, parliamo allora di indice cardiaco se dividiamo la gittata cardiaca per la
superficie corporea. Una persona con normali dimensioni ha una superficie corporea pari a 1.8 m^2
• GS> 40 ml di norma---> dipende molto dalla frequenza cardiaca. Un bradicardico, come un
atleta avrà una frequenza cardiaca a riposo di 50, avrà una gittata sistolica di 100-110 mL. Non
esistono criteri massimi di gittata sistolica. Invece criteri minimi si, a meno che non parliamo di
bambini, per i quali è ammissibile che abbiano una gittata sistolica bassa.
• VFD: ♀,per donne: 56-104 mL; ♂uomini: 67-155 mL---> volume fine diastole
• VFDI: ♀ e ♂: 35-75 mL/m2---> i sta per indicizzazione, cioè viene considerata la superfice
corporea e non tiene conto del genere.
• VFS: ♀: 19-49 mL; ♂: 22-58 mL-->volume fine sistole
• VFSI: ♀ e ♂: 12-30 mL/m2
• EF> 55%--> è molto importante, rappresenta il volume di sangue espulso a fine sistole.
Esprime la quantità di gittata sistolica espulsa rispetto al volume di fine diastole, si esprime in %.
Nelle persone normali deve essere superiore al 55%, se inferiore significa che il cuore non funziona,
la forza con cui il cuore si contrae non è sufficiente a pompare il sangue che c'è nel ventricolo, sotto
i 45% è patologico e si parla di scompenso. Si riscontra molto in clinica, assieme al VFD. Parametri
misurati facilmente con l'ecocardiogramma.
La gittata cardiaca è determinata da diversi fattori:
•
Nodo seno-atriale ( pacemaker naurale del cuore), in altri termini dà la frequenza
cardiaca.
•
Meccanismo principe: aumento dell'attività simpatica, che modula la frequenza
cardiaca. O meglio dipende dall'equilibrio tra parasimpatico e simpatico. A riposo predomina
il parasimpatico, abbiamo 60/70 battiti al minuto. Senza innervazione il nodo SA ha 100
battiti/minuto
•
GS, questo dipende dal volume telediastolico del cuore che a sua votla dipende dal
ritorno venoso. se ho una diminuzione del ritorno venoso, ho una diminuzione della GS
Nel caso in cui il cuore voglia aumentare la sua GC, in caso di emorragia, quindi perdita di
volemia, quindi diminuisce il VFD. Il meccanismo di emergenza, quindi di compenso è l'aumento
della frequenza cardiaca. Per mantenere la quantità costante. Il cuore viene regolato dal sistema
nervoso.
Quando ci sdraiamo la nostra frequenza cardiaca è di 60, in piedi se misuriamo il polso la
frequenza cardiaca sale subito perchè quando sono sdraiato la forza di gravità lo distribuisce verso il
torace. Invece quando siamo in piedi, la forza di gravità lo richiama verso le gambe dal torace,
quindi il VFD diminuisce. Quindi diminuisce anche la GS.
Per mantenere costante la gittata cardiaca aumenta la frequenza, attraverso il sistema nervoso.
Come? Grazie ai barocettori aortici e carotidei percepiscono la caduta di pressione, perchè
diminuisce la GS. I barocettori saranno responsabili dell'aumento della frequenza e della forza con
cui il sangue viene spinto.
Disidratazione da mancanza di acqua presenterà una simile risposta.
•
Un'altra cosa che determinerà la GC sarà l'adrenalina. Perché? E' cronotropa+--->
aumenta la frequenza cardiaca, ma non la GS. E' inotropa +, cioè aumenta la forza con il
cuore si contrae, quindi aumenta la frazione di eiezione. Che passerà dal 55% fino addirittura
al 90%. Negli atleti può arrivare fino a quasi il doppio.
•
Agendo sulla VFD, aumentandolo, modulo la GC, agendo sul ritorno venoso.
Azione inotropa positiva ce li hanno anche gli ormoni surrenalici, in particolare i cortisolici.
La vasopressina, il glucagone. Oltre alle catecolamine circolanti.
Diapositiva riassuntiva:
Ho anche un controllo, una forma di autoregolazione omeometrica, che si basa sull'aumento
del aumentando il volume di pre-carico,quindi il cuore si gonfia, il VFD aumenta, così aumenta
anche la forza di contrazione, in maniera più vivace nel battito successivo.
Condizioni fisiologiche che mi modificano la GC sono :
•
Pasti
• Eccitazione
• Ortostatismo
• Esercizio
Durante il sonno si riduce la frequenza cardiaca e litri circolanti, si riduce anche la GS , è
come se l'organismo andasse in contro ad un'ibernazione
Dopo i pasti abbiamo una frequenza cardiaca più alta, perchè la digestione richiede un certo
impegno metabolico, aumentano il richiamo di sangue e il consumo di ossigeno 300-400ml.
Eccitazione dovuta al sistema nervoso simpatico, argomento già trattato.
ortostatismo: quando ci alziamo di punto in bianco in
piedi la GC tende a ridursi, quindi il cuore cerca di compensare Durante l'esercizio fisico il richiamo di sangue
il richiamo di sangue nelle parti lontane del corpo, con un
da parte dei muscoli è anche maggiore, arriva a
aumento della frequenza cardiaca, perchè ho una diminuizione circa l'80%. Di norma i muscoli richiamono il
20%, anche stando fermi. Del 100% del sangue
di VFD, quindi della GS. Il cuore cerca di compensare, ma
in uscita il 5% se ne va al cuore attraverso le
difficilmente arriva ai livelli di norma e di riposo di GC. Se
coronarie. Il cervello anch'esso richiama
sangue, costantemente. una buona parte se lo
sdraiato ho 5-6L di GC, quando mi alzo posso arrivare al
prende il rene, non solo per funzioni
massimo a 5.5 L di GC. A riposo la gittata cardiaca è più che metaboliche, ma per filtrare il sangue. Infine
sufficiente per garantire le esigenze nutrizionali del nostro
abbiamo il 30% del sangue condiviso tra fegato
e intestino.
organismo.
L'esercizio fisico è il meccanismo principe capace di
modificare la GC, gli atleti possono arrivare fino a 40 l.
Un'altra legge fondamentale nell'apparato circolatorio è la legge di Poiseuille, che mette in
relazione il flusso e la pressione in un circuito in generale. Nel nostro caso applicato al circuito
vascolare.
Che cosa dice questa legge?
Il flusso dentro questo circuito, che è pari alla gittata cardiaca è dato da questa formula:
F= (P1-P2)πr4/8ηl
dove F= flusso, P= pressione, r= raggio del condotto, η=viscosità del fluido, l= lunghezza del
condotto
• La quantità πr4/8ηl è la resistenza (R) che incontra
il flusso a scorrere in quel condotto--> può essere tranquillamente espresso come R
• la 1) si può anche riscrivere come F= ΔP/R
Posso riscrivere la legge anche come R= ΔP/F
• Per cui: R= Pressione aortica-pressione vena
cava (o negli atri)/GC ---> serve perchè è uno dei parametri importanti nello studio
dell'apparato cardiocircolatorio in fisiologia e in medicina
Ognuno di questi termini ha un significato. P indica la pressione ai capi del circuito, quindi in
ingresso e in uscita. R è il raggio medio del condotto irregolare. NB: Il raggio è elevato alla 4. η
indica la viscosità del fluido. L sta per lunghezza del condotto.
La pressione aortica la posso calcolare misurando la pressione nel braccio con un manicotto.
La pressione della vena cava nell'atrio dx, intanto la posso desumere rispetto alla pressione aortica,
che è attorno ai 100, oppure mando un piccolo catetere nella vena cava, che misura la pressione in
modo diretto, oppure in modo indiretto attaverso l'ecocardiogramma. La gittata cardiaca la posso
ottenere con la legge di Fick.
Ho a che fare soprattutto con costanti π/8ηl, tranne che per r4 --> questo è il raggio medio
arteriolare ed è un indice della resistenza al flusso del sistema cardiocircolatorio. Piccole variazioni
del raggio, mi determinano grandissime variazioni di resistenza, perchè il raggio è alla quarta.
Supponiamo che abbiamo una grossa perdita di liquidi, ho emorragia, ho ipovolemia, discesa
della pressione.
Aumenterà la frequenza cardiaca e avremo una scarica adrenergica che ridurrà i raggi delle
arteriole, che son vasi dotati, nella loro parete, di muscolatura liscia con recettori adrenergici.
Variano immediatamente il loro calibro, regolando la pressione arteriosa. La riduzione del diametro,
mi corrisponde ad un aumento della resistenza al flusso. L'aumento della resistenza comporta un
aumento della pressione che tende a riportarla a valori normali (se i due eventi, abbassamento in
origine della pressione e vasocostrizione, avvengono contemporaneamente allora si avrà il
mantenimento di una pressione costante). La pressione sarà uguale al flusso\resistenza. Questo è il
motivo per cui chi ha subito una perdita di liquidi è pallido e freddo, proprio a causa di questa
vasocostrizione a livello periferico e quindi soprattutto a livello della cute.
Lezione di fisiologia, 27/03/2012
Prof. Crisafulli
Roberto Serreli
Ripartiamo da questa legge che mette in relazione il flusso in un condotto con la pressione, la
differenza di pressione ai capi del condotto stesso e le resistenze. Abbiamo visto che questa legge
dice che il flusso, a cui possiamo sostituire il termine gittata cardiaca, è uguale alla differenza di
pressione ai due capi del condotto, e i due capi sono il ventricolo di sinistra e l'atrio di destra, dove
si chiude praticamente il circuito, per πr4 , dove r è il raggio medio arteriolare, cioè il raggio medio
dei condotti, in particolar modo arteriolare perché è là che si può compiere una regolazione dei
diametri. E’ molto importante il fatto che questo fattore r sia alla quarta, il che vuol dire che piccole
variazioni di raggio, in dilatazione o in riduzione del raggio stesso, danno grandi variazioni del
flusso stesso, fratto 8 per η, dove per η si intende la viscosità del sangue, che dipende
essenzialmente dal numero di globuli rossi presenti nel sangue (è data anche da altre cose, ma la più
importante è appunto l'ematocrito), e l è la lunghezza del condotto. In una persona normale, che non
abbia problemi, in condizioni stabili, l'unica cosa che può variare di questi fattori sono la differenza
di pressione e r4, perché la viscosità del sangue non cambia, a meno che uno non abbia una
emorragia improvvisa, e anche la lunghezza dei condotti non cambia. A cosa serve questa
espressione? intanto ci consente di calcolare le resistenze, questo fattore qua (πr4 / 8ηl) rappresenta
la resistenza che il sangue incontra nel circuito, quelle che vengono chiamate resistenze vascolari
periferiche, perché stiamo parlando non del circuito polmonare ma del circuito sistemico; quindi, le
resistenze sono incorporate in tutto questo fattore qui, e questa equazione può anche essere scritta
come F (flusso) = ΔP / R, o anche Gittata Cardiaca = ΔP / R. Il flusso, e quindi la gittata cardiaca è
possibile calcolarlo con vari sistemi; ΔP lo possiamo calcolare? Si, basta conoscere la pressione che
c'è nel ventricolo di sinistra, e la pressione che c'è nel ventricolo di sinistra in sistole è quella che c'è
anche nell'aorta, per cui basta che io misuri la pressione aortica, e automaticamente ho anche la
pressione del ventricolo di sinistra. Possiamo calcolare la pressione dell'atrio di destra? Si, possiamo
per esempio prendere per buona la pressione che misuriamo nella vena cava, non è difficile da
calcolare anche con sistemi indiretti, oppure possiamo approssimare che (lo sappiamo) la pressione
all'interno dell'atrio di destra è circa 3,4, massimo 5 mmHg, poca cosa rispetto ai 100-120 che ci
sono in aorta, per cui questo fattore qui è calcolabile. E a cosa ci serve? ci serve a calcolare le
resistenze. Le resistenze hanno una grande importanza perché sono uno dei fattori più importanti
nella regolazione della pressione arteriosa, cioè dalla modulazione delle resistenze si modula la
pressione arteriosa, lo vedremo nella prossima lezione. E’ anche importante perché in alcune
patologie si ha un aumento importante delle resistenze, per esempio nello scompenso cardiaco, dove
si tenta di mantenere la pressione stabile nel circuito incrementando le resistenze. Oppure è
importante in altre patologie dove le resistenze periferiche cadono in maniera drammatica, per
esempio malattie metaboliche come l'ipertiroidismo che causa una intensa vasodilatazione, e che
quindi va a influenzare questo fattore qui vasodilatando: quindi si riducono enormemente le
resistenze e questi signori hanno un calo di pressione proprio perché hanno una vasodilatazione
generalizzata. Per cui, applicando questa semplice equazione, si possono avere una serie di
informazioni importanti sullo stato del nostro apparato cardiocircolatorio. Quindi alla fin fine
nell'apparato cardiocircolatorio le cose che ci interessano sono 3:
1) il flusso ( o cioè la gittata cardiaca)
2) la pressione
3) le resistenze
tutte cose che si possono calcolare.
Questa legge che vi ho fatto appena vedere, nella sua approssimazione è valida quasi
sempre, però ci son dei limiti. Uno dei limiti più importanti è questo, che il flusso del fluido, in
questo caso del sangue, attraverso il torrente circolatorio, sia laminare: se ci sono per esempio
stenosi (restringimenti dei vasi) o dilatazioni dei vasi e i flussi diventano turbolenti, questa legge
non vale più. Altra cosa importante è che la viscosità del sangue rimanga costante, e anche questa è
una approssimazione che in alcuni casi non corrisponde alla realtà, perché soprattutto nei grossi
tronchi arteriosi c'è un effetto sui globuli rossi, un effetto dinamico per cui tendono ad accumularsi
al centro del vaso, c'è un "effetto scrematura" si chiama, cioè i globuli rossi nel sangue tendono ad
accumularsi soprattutto al centro del vaso lasciando il plasma in periferia, e questo fa si che la
viscosità del sangue non sia costante ma sia maggiore al centro del vaso rispetto alla periferia del
vaso stesso. Per cui, anche in questo caso l'applicazione della legge di Poiseuille non è perfetta.
Altra cosa molto importante è che i condotti devono avere pareti rigide, cioè non devono
deformarsi per effetto dell'onda pressoria causata dal flusso che passa attraverso il condotto stesso; e
anche questa non è la verità: nel torrente circolatorio, lo vedremo la prossima lezione, soprattutto le
arterie non sono affatto rigide, anzi sono elastiche e smorzano l'onda pressoria. Per cui ci sono delle
limitazioni a questa legge che però è applicabile, con determinati limiti: ci sarà un errore sistematico
nell'applicarla.
Abbiamo visto fino ad ora i flussi, altra cosa importante nella valutazione dell'apparato
cardiocircolatorio sono i volumi cardiaci: quanto è grande il cuore, quanto è spesso e cose del
genere. Sono tutte cose che ormai si fanno (al 99%) con questo tipo di metodica qui, cioè con
l'ecocardiografia. In alcuni casi, quando l'ecocardiografia non è possibile, per motivi di scarsa
ecogenicità (capacità di un mezzo di generare eco) del paziente, o quando bisogna avere una
risoluzione molto alta, allora si va a fare anche la risonanza magnetica nucleare (RMN), che è
una metodica molto più macchinosa e costosa. Ha una risoluzione migliore ma la si fa solo in
determinati casi, anche perché non possiamo sottoporre tutti a RMN: intanto perché è seccante, si
tratta sempre di mettere il paziente dentro a un tubo, quindi se si può non la si fa. Poi abbiamo
anche altri metodi, come la scintigrafia radioisotopica, si tratta di metodiche con radioisotopi,
quindi si danno radiazioni al paziente (se fa si evita); o l'angiografia, metodica ancora più invasiva,
si tratta di mandare cateteri dentro arterie e vene. Sono tutte cose che si fanno , ma solo in casi
molto selezionati.
L'ecografia, dicevo, è la cosa più semplice, è la cosa con cui verosimilmente avrete a che fare
in futuro, è molto semplice da fare. Vi porto un esempio di un dato di frazione di eiezione, di un
volume di fine diastole, un volume di fine sistole e via dicendo, di un cuore di una persona normale
in questo caso. Allora, questo è il nostro cuore visto dalla "finestra apicale", cioè mettiamo una
sonda all'apice del cuore. Quindi il cuore si vede così: qua c'è il ventricolo di sinistra, qua il
ventricolo di destra che qua non si vede bene, qua l'atrio di destra che invece si vede bene, e l'atrio
di sinistra è questo. Questa dicevo è la cavità del ventricolo sinistro in telediastole, cioè prima che
poi si contragga, lo vediamo perché c'è una freccetta qui che ci indica esattamente che stiamo
prendendo questo fotogramma poco prima del complesso QRS del cuore il che significa che si è
appena contratto l'atrio, il ventricolo sinistro è riempito al massimo per cui è al suo massimo
volume. Si tratta semplicemente di seguire i contorni della parete interna del cuore e considerando
(usando una serie di equazioni) come se fossero tanti strati uno sull'altro, tante sezioni di tronchi di
cono. E' un algoritmo matematico, lo fa direttamente la macchina, il medico non deve far niente se
non seguire il contorno di questa cavità. Esistono fra l'altro varie equazioni, questa si chiama tecnica
di Simpson (il metodo più accurato per la valutazione dei volumi polmonari è il metodo si Simpson
perché può essere utilizzato, a differenza di altri metodi, anche in presenza di alterazioni importanti
della geometria ventricolare ed in presenza di aneurismi. Tale metodo si basa sul concetto della
sommazione dei volumi di una serie di sezioni cilindriche sovrapposte, ed è tanto più accurato
quanto più numerose sono le sezioni sommate), EDV è l'acronimo inglese di End Diastolic Volume,
cioè volume di fine diastole, e come vedete qua questo soggetto ha 121ml che è un quadro normale.
Stesso soggetto, cioè stesso cuore, stesso giorno, stessa seduta, qua però stiamo prendendo il
cuore, i contorni interni della parete del ventricolo di sinistra dopo che il ventricolo si è
contratto, perché vedete qua la freccetta fucsia, a questo livello qui, vuol dire che è passato il QRS,
è passata anche l'onda P, quindi il ventricolo si è contratto e si sta ripolarizzando. E' il volume
minimo, ""si è spremuto"". E a questo punto tira fuori l'algoritmo, tutta una serie di parametri, che
sono ESV, che vuol dire End Sistolic Volume, cioè volume di fine sistole, e il nostro signore ha 48
ml, SV che vuol dire Stroke Volume, che vuol dire gittata sistolica, cioè il volume di sangue che il
cuore ha eiettato in quel battito, che sono 73.06 ml, che altro non è che questo (EDV) meno questo
(ESV); EF che vuol dire Ejection Fraction, cioè frazione di eiezione, è 60.2%, il che vuol dire che il
cuore in questo caso ha eiettato il 60% circa della quantità di sangue che era contenuta nel suo
interno prima della sistole; SI che vuol dire Sistolic Index (io su internet l’ho trovato come Stroke
Index, Indice Sistolico, valore che esprime il volume di sangue espulso in una contrazione da ogni
ventricolo per m2 di superficie corporea), che vuol dire indice sistolico, che altro non è che la gittata
sistolica divisa per i metri quadri di superficie corporea, quello che vi avevo detto la lezione passata,
è uno dei sistemi per indicizzare, per fare in modo di uniformare le misure su persone di taglia
diversa, per cui io divido per metri quadri la superficie corporea e trovo questo valore qua, che è
anch'esso normale. Per fare questa operazione qui bisogna fare una ecografia e ci vogliono 5 minuti,
è molto semplice. Quindi, questo è un esempio di come si può calcolare la frazione di eiezione, la
gittata cardiaca, questa ovviamente è la gittata sistolica, poi noi qua avremo anche la frequenza
cardiaca che vedete riportata qui, intorno ai 60, basta moltiplicare 60 per lo Stroke Volume (SV),
cioè per la gittata sistolica, e otteniamo la gittata cardiaca. Quindi, un sacco di informazioni in una
immagine. Si possono poi fare tante altre cose, ma lo scoprirete poi se andrete a fare cardiologia.
Un'altra tecnica che si usa abbastanza frequentemente ma soprattutto la si usa per valutare gli
stati di sofferenza miocardica dovuti a insufficienza delle coronarie, cioè quando arriva poco
ossigeno al cuore, è la scintigrafia, si tratta di mandare nel torrente circolatorio sostanze marcate
con isotopi radioattivi, queste sostanze possono essere anche i globuli rossi stessi ma più spesso
albumina marcata con sostanze radioattive, che si distribuiscono nel torrente circolatorio e
passano anche nel cuore, ovviamente. Si mette un rilevatore di radioattività sul torace e si va a
seguire quello che succede nel cuore e si possono anche indirettamente calcolare i volumi cardiaci
in varie proiezioni e risalire alle volumetrie e di conseguenza alla frazione di eiezione, in questo
caso vedete che questo soggetto ha una frazione di eiezione particolarmente bassa, 18%, che vuol
dire che ha un cuore che non funziona. Tutto viene fatto, ovviamente, con algoritmi matematici,
cioè con sistemi costruiti apposta per analizzare questo tipo di immagini. Non è una cosa che si fa
frequentemente perché è un esame invasivo.
Il cuore quando pompa sangue nel torrente circolatorio fa un lavoro, un lavoro inteso in senso
fisico ossia induce lo spostamento di una massa, in questo caso il sangue, non solo imprimendogli
una pressione ma anche una velocità. Esistono vari sistemi per calcolare che lavoro fa il cuore
durante la sua attività, esistono varie formule e io vi ho portato quella più semplice, che dice che il
lavoro cardiaco è uguale a
PmGp+1/2mv2
dove
Pm è la pressione media in aorta, praticamente è la pressione che ha il ventricolo di sinistra
(questo è il lavoro cardiaco del ventricolo di sinistra, non di tutti e due), ci dice in pratica contro
quale pressione il ventricolo deve lavorare;
Gp è la gittata sistolica espressa in ml, cioè il volume che il cuore immette nel torrente
circolatorio,
1/2 mv2, per chi si ricorda un po’ di fisica questa è una energia cinetica,
m altro non è che la gittata sistolica non più espressa in ml ma espressa in grammi (nella slide
è segnata come “massa della gittata sistolica”) e
v è la velocità media di scorrimento del sangue nell'aorta.
Questa equazione nel calcolo del lavoro cardiaco non viene usata mai, non viene applicata
almeno in medicina pratica, viene applicata magari da chi fa fisiologia sperimentale, soprattutto
sugli animali, perché per calcolare tutte queste cose (massa della gittata cardiaca, della gittata
sistolica ad esempio) devi prendere il sangue in esame, insomma tutte cose un pochino invasive, in
particolar modo la velocità media di scorrimento del sangue in aorta non è che sia così semplice da
calcolare, si può fare col Doppler (L'ecografia Doppler o più semplicemente ecoDoppler è una
tecnica non cruenta e non invasiva quindi facilmente ripetibile utilizzata in medicina per lo studio
della situazione anatomica e funzionale dei vasi sanguigni, arteriosi e venosi, e del cuore in tempo
reale. Prende il nome dal suo principio fisico di funzionamento, l'effetto Doppler) ma in fisiologia
sperimentale bisogna avere un flussimetro dentro l'aorta, una cosa che non si fa più sui pazienti. In
clinica si usa quello che è chiamato doppio prodotto, che altro non è che il prodotto della
frequenza cardiaca per la pressione arteriosa sistolica (FC x PAS), che non è il lavoro cardiaco
vero e proprio ma ci si avvicina molto.
Quindi da un punto di vista pratico voi in futuro troverete soprattutto questa notazione qui, il
doppio prodotto, che è una buona approssimazione del lavoro cardiaco. Se vogliamo sapere il
lavoro del ventricolo di destra dobbiamo sostituire a questa equazione non più la pressione media
dell'aorta ma la pressione media dell'arteria polmonare, perché il ventricolo di destra pompa sangue
nell'arteria polmonare che ha una pressione molto più bassa dell'aorta. Tutto il resto rimane uguale,
perché la gittata sistolica rimane identica nei due ventricoli, la velocità anche, mentre quello che
cambia è la pressione: mentre nell'aorta è in media 100 mmHg, nell'arteria polmonare è intorno ai
15 mmHg, quindi circa 6-7 volte inferiore, tant'è che il lavoro del ventricolo di sinistra è 6-7 volte
superiore a quello del ventricolo di destra semplicemente perché pompa contro un regime
pressorio che è 6-7 volte più grande. Altra cosa che si calcola in fisiologia sperimentale e non in
clinica, quindi non lo troverete quando farete clinica, è il rendimento del lavoro cardiaco, che poi
è giusto conoscere, e per calcolarlo bisogna sapere che lavoro fa il cuore, usando tutte le formule
che abbiamo visto prima, e qual è la spesa energetica. La spesa energetica si può calcolare
abbastanza semplicemente calcolando il consumo di ossigeno del cuore: si prendono campioni di
sangue in ingresso e in uscita dal cuore (sempre in fisiologia sperimentale), si fa la differenza
artero-venosa e si va a vedere quale è il consumo di ossigeno del cuore. Sapendo il consumo di
ossigeno e sapendo che un litro di ossigeno sviluppa circa 5kcal, si calcola quale è la spesa
energetica. Sapendo qual è il lavoro e dividendolo per la spesa energetica si ha il rendimento, che è
niente di meno di quello che abbiamo visto con lo scheletrico, è lo stesso principio.
Sperimentalmente, il rendimento del lavoro cardiaco è intorno al 18-20%, che è un valore basso,
più basso di quello del muscolo scheletrico. Non solo, facendo un po' di esperimenti si è visto che è
più conveniente aumentare il lavoro cardiaco aumentando i volumi piuttosto che la frequenza
cardiaca, mi spiego meglio: supponiamo che io abbia una gittata cardiaca a riposo di 5 litri e la
devo portare a 10, devo raddoppiarla. Per raddoppiare questa gittata cardiaca più o meno devo
raddoppiare il lavoro, e ho due strade: o aumento la frequenza cardiaca, cioè aumento il numero dei
battiti per minuto, o aumento la gittata sistolica, cioè aumento il volume che io espello per ogni
singolo battito, oppure aumento tutti e due (che è quello che più frequentemente succede). Se io
vado a misurare il rendimento in queste due situazioni, cioè aumentando la frequenza cardiaca o
aumentando la gittata sistolica si è visto che è più conveniente questo tipo di strategia qui
(aumentare la gittata sistolica). Cioè se aumento la frequenza cardiaca il rendimento tende ad
abbassarsi, se invece aumento la gittata sistolica il rendimento tende a rimanere costante,
insomma è migliore rispetto che aumentare solo la frequenza cardiaca. Il motivo per cui questo
avviene non si sa, sperimentalmente è così (a tal proposito cito invece un libro: La circolazione
coronarica: fisiopatologia clinica e farmacoterapia Di Gianni Losano,Mario Marzilli,Mariano
Ferrari. “Il rendimento del cuore varia a seconda del tipo di lavoro che viene compiuto: a parità di
prodotto FC x PAS, può essere bassa la pressione ed elevata la gettata sistolica o elevata la
pressione e bassa la gettata sistolica. Nel primo caso si parla di lavoro prevalentemente di
pressione nel secondo caso di lavoro prevalentemente di volume. Poichè nel lavoro di pressione la
maggior parte dell'energia viene utilizzata nella sistole isovolumica che termina solo quando viene
superata la pressione presente nell'aorta alla fine della diastole e poiché nella sistole isovolumica
non viene compiuto lavoro pur essendoci dispendio energetico, è facile capire come nel lavoro di
pressione il rendimento è minore rispetto al lavoro di volume”. Mario)
Questo ha un'importanza pratica, per esempio a un cardiopatico far aumentare molto la
frequenza cardiaca può far aumentare tanto il lavoro e abbassare il rendimento; mentre in una
persona normale non succede niente, in una persona che ha dei deficit cardiaci di qualsiasi tipo far
aumentare la frequenza cardiaca può essere un problema. Questo è il motivo per cui, ad esempio,
negli scompensati cardiaci funziona benissimo un farmaco che si chiama beta bloccante, che
blocca i recettori beta del sistema simpatico e impedisce che la frequenza cardiaca salga troppo,
blocca quindi gli incrementi di frequenza cardiaca. Funziona benissimo in questi soggetti perché
ogni qualvolta la frequenza cardiaca sale al di sopra di determinate soglie hanno crisi cardiache,
quindi questo concetto è bene tenerlo presente anche per fini applicativi. Altra cosa che non piace
tanto al cuore è l'aumento di lavoro dovuto ad aumenti di pressione, cioè quando ad esempio
per effetto di un'eccessiva vasocostrizione lui deve pompare sangue non più contro una pressione di
100 mmHg in aorta, ma contro una pressione di 120-130 mmHg. Aumenta il lavoro cardiaco anche
in questo caso, e si abbassa il rendimento. E' una cosa che evidentemente non piace al nostro organo
ed è anche un'altra cosa da tenere a mente, perché, stesso discorso di prima, anche in questo caso far
fare al cuore un lavoro contro pressione arteriosa non è conveniente. Questo è il motivo per cui si
danno, soprattutto a chi soffre di insufficienza coronarica o anche di scompenso cardiaco, dei
vasodilatatori per evitare che ci siano aumenti di pressione e vasocostrizioni eccessive. Quindi altro
concetto che si deve tenere a mente, è buona norma nei pazienti cardiologici evitare eccessivi
incrementi di pressione arteriosa, mentre nelle persone normali non ci sono problemi, gli aumenti
di pressione sono tollerati benissimo.
Parliamo ora dei substrati che utilizza il cuore per compiere il suo lavoro. Il cuore è un
organo prettamente unico: funziona sempre, ha al limite delle transizioni da situazioni da livelli
metabolici più bassi a livelli metabolici superiori, e in ogni caso queste transizioni le fa molto
rapidamente e con minimo dispendio aerobico;
il cuore è un muscolo aerobico, e difatti lo testimonia il fatto che è pieno di mitocondri e che
produce pochissimi metaboliti anaerobici, ne produce comunque, ma tende addirittura a mangiare
quelli che vengono prodotti dai muscoli scheletrici. Questa diapositiva ci dà dei valori medi: a
riposo, un buon 34% del metabolismo cardiaco è sostenuto dagli acidi grassi, quindi il cuore è
anche un ottimo "mangiatore" di acidi grassi; un altro 30% circa dal glucosio ma un'altra bella quota
(siamo quasi al 30%) dal lattato, cioè dall'acido lattico, che si trova naturalmente nel nostro sangue,
anche a riposo, e viene mangiato fra gli altri organi anche dal cuore (un po’ se lo prende il fegato ma
una bella quota se la prende il cuore); il cuore è anche un buon mangiatore di corpi chetonici,
soprattutto se vengono formati in periodi di digiuno. Se andiamo nell'esercizio una buona quota del
metabolismo cardiaco viene sostenuta dall'acido lattico: il cuore lo prende, lo converte in acido
piruvico e lo fa entrare nel ciclo di Krebs, semplicemente, e lo utilizza come metabolita. Questo è
un altro modo per sfatare il mito dell'acido lattico come un qualcosa di intossicante, è un substrato
energetico che infatti viene utilizzato dai muscoli ma anche dal cuore per sostenere le proprie
necessità.
In condizione di esercizio il cuore "shifta" il proprio metabolismo e si mette a "mangiare"
l'acido lattico, soprattutto quando esso è prodotto in quantità notevoli dai muscoli scheletrici. Il
consumo di ossigeno del cuore (a riposo) è circa 250-300 ml / minuto, in una persona normale.
Durante sforzo, questa quantità può anche diventare 5-6 volte più grande, cioè può raggiungere 1L,
1,5 L di consumo, il che vuol dire 5kcal / min (se consuma un litro di ossigeno).
Adesso introduciamo il sistema vasale. Qua cominciamo a vedere che esistono tanti tipi di
vasi: in particolar modo abbiamo le grosse arterie (che troviamo in uscita dal cuore), che hanno
delle pareti particolari, in particolar modo comincio a farvi notare questo strato qui, lo strato di
tessuto elastico, le grosse arterie e soprattutto l'aorta hanno un grosso strato di fibre elastiche, le
quali sono presenti in gran numero, e il motivo, che vedremo poi, è, ovviamente, funzionale. Via via
che le arterie scendono di calibro si arriva alle arteriole: la loro caratteristica principale è quella di
avere tante cellule muscolari lisce che ne avvolgono la parete, e come ho già detto questo ha un
significato nella regolazione delle resistenze al flusso; mentre infatti i grandi vasi variano di poco il
loro calibro per azione del muscoli lisci, queste arteriole lo variano tantissimo, per cui il grosso
della regolazione della resistenza al flusso avviene qui, variando il calibro delle arteriole. Poi
arriviamo via via alle arteriole più piccole, alle meta-arteriole e via dicendo fino ai capillari, che
hanno una parete composta essenzialmente da cellule endoteliali, molto importante peraltro.
Andiamo poi alle venule, che sono soprattutto composte nella loro parete da tessuto
connettivo (e anche questo ha un senso, poi lo vedremo), per arrivare poi alla fine alle grosse vene,
come le vene cave, le quali hanno tanto tessuto connettivo ma anche fibre elastiche, non tante
quante l'aorta o le grandi arterie, e hanno anche un po' di cellule muscolari: le grandi vene infatti
possono anche contrarsi. Qua (immagine sopra) vediamo un taglio perfetto di una grande vena e
una grande arteria, si vede subito quale è la differenza, la parete dell'arteria ha una importante
componente elastica, mentre la parete della vena è molto più sottile e anche più irregolare di calibro,
meno robusta. In base alla loro funzionalità, i vasi ematici li distinguiamo in:
vasi a prevalente funzione elastica (grosse arterie),
vasi a prevalente funzione di resistenza (le arteriole), quelle che, vi ripeto nuovamente,
hanno tante cellule muscolari lisce, poi abbiamo gli
sfinteri pre-capillari, i quali sono appunto degli sfinteri posti prima dei letti capillari e
regolano il flusso locale di sangue. I vasi di scambio sono i capillari, i vasi di capacità sono
invece le grosse vene; i vasi di shunt (non li abbiamo visti), sono quei vasi che normalmente sono
chiusi ma che si possono aprire in condizioni particolari, come quando ad esempio c'è l'occlusione
di un vaso e c'è bisogno di vicariare la circolazione attraverso un altro vaso. Sono vasi che si aprono
al bisogno, ce ne sono tanti in determinati organi, come ad esempio nel polmone.
Questa immagine vi fa vedere in sezione quello che vi ho raccontato. Qua abbiamo il
diametro medio dei vasi, lo spessore della parete, lo spessore dell'endotelio, la quantità di tessuto
elastico, la quantità di tessuto muscolare liscio, la quantità di tessuto fibroso, cioè di collagene,
sostanzialmente. Hanno preso l'aorta, il cui diametro è di norma sui 25 mm, e vedete che gran parte
della sua parete è fatta da tessuto elastico. Scendendo verso le arterie di calibro più piccolo, non
cambia molto se non il diametro, ma la parete è sempre piena di tessuto elastico. Le cose cambiano
quando abbiamo le arteriole dove prende il sopravvento il tessuto muscolare liscio. Nello sfintere
pre-capillare ugualmente ci sono tanti muscoli lisci (per il motivo che ho detto prima, sono vasi che
si aprono e si chiudono sulla base delle esigenze metaboliche locali, per cui hanno bisogno di molti
muscoli che aprono o chiudono il vaso al bisogno). Il capillare ha solamente endotelio, le venule
cominciano ad avere oltre all'endotelio, anche il tessuto fibroso, il tessuto connettivo, e via via vene
di piccolo e poi di grande calibro, come la vena cava che come vedete ha nuovamente una parete
elastica, hanno comunque ancora tanto tessuto fibroso e anche muscolatura liscia, che le fanno
"svuotare" in determinate circostanze (più che svuotarsi, contrarsi e far fluire il loro contenuto di
sangue).
Abbiamo visto quali sono le caratteristiche anatomiche di questi vasi, andiamo a vedere il
comportamento delle fibre elastiche e delle fibre collagene del tessuto fibroso in relazione alla
lunghezza e alla tensione che sviluppo. Nella realtà non esistono arterie elastiche pure o vasi con
solo fibre collagene, sono sempre un misto.
Le fibre elastiche hanno questo comportamento: più si stira la fibra, più aumenta la tensione,
c'è un rapporto quasi lineare tra lunghezza e tensione, ovviamente fino a un punto di rottura, cioè
fino a un punto in cui la fibra stessa tende a rompersi. La fibra collagene ha un altro tipo di
comportamento: per distensioni fino a un certo punto non si hanno aumenti di tensione, poi a un
certo punto per piccole variazioni di lunghezza della fibra la tensione aumenta enormemente. Poi in
realtà la maggior parte delle nostre arterie hanno un tipo di comportamento intermedio, sono miste
(vedi linea tratteggiata), hanno infatti sia la componente elastica che la componente collagene, che
si evidenzia soprattutto a determinati livelli di tensione; mi spiego: comanda la fibra elastica per
determinate distensioni del vaso, dopodiché è la fibra collagene che "governa" lo stato di tensione
per ulteriori distensioni della fibra stessa.
Qual è il significato funzionale della distensione delle arterie? Devono smorzare l'onda
sfigmica, cioè l'onda di gittata sistolica, l'onda pressoria che arriva per ogni singola contrazione
cardiaca. Sono poste in prima fila a smorzare l'incremento pressorio dovuto alla contrazione dei
ventricoli, in particolar modo del ventricolo sinistro, visto che è quello che pompa in maniera più
potente. Quindi in sistole tendono ad assorbire la pressione, immagazzinarla sotto forma di energia
elastica, e restituirla poi in diastole quando la valvola aortica è chiusa. Questo lo fanno tanto più
quanto più sono vicine al cuore. Se voi guardate (immagine sotto), questo è l'andamento pressorio
durante i cicli cardiaci nel ventricolo di sinistra, nell'arteria aorta e vedete quest'oscillazione
continua per ogni singolo ciclo cardiaco. Tende a fare in modo che la pressione data in sistole venga
restituita in diastole, funziona proprio come un sistema di tamponamento, come per "smorzare"
quest'onda, e questo effetto si vede mano a mano che ci si allontana dalla sorgente delle variazioni
di pressione, cioè mano a mano che ci si allontana dal cuore, fino ad arrivare a un certo punto in cui,
a livello delle arteriole più periferiche, l'onda non è più un onda sfigmica, cioè non è più una
alternanza di minimi o massimi, ma diventa un'onda continua, per effetto dei continui smorzamenti
che ha subito in tutti i tratti che ha attraversato. Vedete poi che via via che si va avanti la pressione
passa da una media di 100mmHg circa fino ad arrivare a livello dei capillari a una pressione che
scende intorno ai 30-35mmHg, per ritornare poi a livello dell'atrio di destra intorno ai 3mmHg, cioè
c'è una progressiva attenuazione della pressione che nel primo tratto è fatta soprattutto dalle pareti
elastiche delle arterie e poi via via, per attrito. Sulla destra vedete quello che succede a livello del
circolo polmonare, che poi è uguale a quello che succede nel circolo sistemico, ma in scala ridotta,
6-7 volte inferiore. Qui al lato abbiamo la stessa immagine di prima, fatta in maniera un po’ più
didattica. Quindi, le arterie elastiche hanno il compito di smorzare l'onda sfigmica, è una
caratteristica che hanno soprattutto le arterie giovani, nei bambini, nei ragazzi, e che viene via via
persa con l'invecchiamento dell'apparato cardiocircolatorio. Succede che con l'invecchiamento
l'aorta tende sempre meno ad assorbire l'onda sfigmica, quindi a smorzare l'onda pressoria e a
restituire dopo. Quindi può accadere che questa "forchetta" che vedete qui si allarghi, cioè la
pressione arteriosa sistolica, quella massima, tende a salire perché manca l'effetto di smorzamento, e
a volte quella diastolica tende a diminuire perché manca durante la diastole il ritorno elastico
dell'arteria. Per cui si dice che aumenta la pressione differenziale, che è un classico del cuore
senescente, questo perché le fibre elastiche vengono sostituite da altri tipi di fibre, e spesso vanno
anche incontro a calcificazione.
Il comportamento delle vene ve l'ho già in parte spiegato prima, è molto diverso rispetto a
quello delle arterie, non smorzano ma tendono a contenere. Questa è la relazione tra la pressione
all'interno della vena (transmurale = a cavallo della parete) e la sezione della vena stessa. Come
vedete, per piccole variazioni di pressione si hanno grandi variazioni di volume, cioè tendono non a
distendersi elasticamente ma a contenere, a fare da serbatoi, fino ad arrivare a un certo punto dove
vengono stirate le fibre collagene e a quel punto diventano dei contenitori più o meno rigidi, cioè
tendono a non contenere più ulteriori volumi di sangue. Però hanno una grandissima capacità di
contenimento e anche di svuotamento, perché le grosse vene hanno anche delle cellule muscolari
lisce nella loro parete, per cui tendono a "spremersi" in determinate circostanze: quando c'è bisogno
di un recupero volume sanguigno, ad esempio nell'emorragia, dobbiamo mobilitare masse
sanguigne importanti. In effetti quello che fanno le vene è sequestrare gran parte del volume
ematico circolante.
Come vedete, ponendofacendo 100 la quantità di sangue circolante un buon 60% (61%) è
contenuto nel sistema venoso, più della metà del sangue del nostro organismo dunque è dentro le
vene, non si sa mai che possa servire. Una certa quota ovviamente ce l'ha la circolazione polmonare
(12%), il cuore (9%), le arterie non ne hanno tantissimo (11%), arteriole e capillari (7%). Gran parte
quindi, più della metà, è contenuto dentro le vene.
Adesso andiamo a vedere che rapporto c'è tra la velocità che ha il sangue nei vasi e la
sezione del vaso stesso. Come vedete questa è l'area della sezione totale dei vasi, come se noi
facessimo in pratica un taglio su tutti quei vasi che in quel momento vengono attraversati dal
sangue, a livello di aorta, arterie, arteriole, capillari, venule e vene. L'area della sezione (che è
espressa in cm2) cresce progressivamente dall'aorta fino ai capillari, che hanno la più grande
superficie di tutti i vasi, per ovvi motivi: sono vasi di scambio, devono scambiare metaboliti,
ossigeno e anidride carbonica con i tessuti; e poi via via l'area della sezione torna a scendere nelle
vene, le quali comunque hanno un'area di sezione maggiore dell'aorta.
La velocità del sangue subisce delle brusche variazioni lungo il torrente circolatorio: è
massima nell'aorta (stiamo parlando di cm /s) anche se in questa immagine è un po’ bassa, si vedono
velocità ben maggiori di 30 cm /s, forse anche 100 (1 m /s); è importante notare che a livello dei
capillari la velocità scende quasi a zero, qui il sangue scorre molto lentamente (il motivo è lo stesso
che abbiamo espresso prima parlando della sezione, cioè favorire scambi di metaboliti, ossigeno e
anidride carbonica, aumentando il tempo di contatto del sangue con i tessuti); poi via via la velocità
torna piuttosto alta nelle vene ma non raggiunge quella delle arterie.
Vediamo il volume ematico contenuto nei vasi, il massimo è appunto nelle vene: se uniamo
il volume ematico delle vene con quello della vena cava otteniamo, come abbiamo visto prima, un
buon 60% del sangue totale contenuto nel nostro organismo. Più in basso abbiamo la velocità di
scorrimento del sangue (peraltro all'inizio è pulsatoria, ovviamente, cioè ha momenti di
accelerazione e decelerazione dovuti all'attività cardiaca, è l'onda sfigmica che viene
progressivamente smorzata e poi riprende a livello venoso); ancora più in basso vediamo la
pressione, che abbiamo già visto precedentemente.
Ora affrontiamo il discorso del flusso laminare e del flusso turbolento. Normalmente il
flusso a livello dei vasi ematici è di tipo laminare, cioè con una velocità crescente dalla periferia del
vaso verso il centro del vaso stesso, come se ci fossero tanti cilindri che scorrono uno sull'altro.
Questo accade nella normalità, a parte delle rare eccezioni perché ci possono essere
turbolenze per effetto di alte velocità, quando ad esempio una persona ha il cuore particolarmente
"prestante".
In alcune situazioni però il flusso può diventare turbolento, si creano dei vortici e il flusso
stesso non è più laminare, e questo accade quando viene superato il numero di Reynolds, che è
pari a 2000.
Abbiamo qui una equazione
N=rvδ/η
con
r= raggio del condotto che si sta attraversando(è importante ma la cosa più importante è il
secondo termine, v),
v= velocità di scorrimento del fluido (è lei che effettivamente ci dà le turbolenze nella
fisiologia ma anche in patologia; questa velocità dipende dal calibro del vaso, cioè più io ho un vaso
stretto, più la velocità per passare dentro quella strettoia deve aumentare, e questo può creare
turbolenze),
δ= densità (di solito è un parametro costante),
η=viscosità (anch'essa in linea di massima rimane stabile, a meno che uno non diventi
anemico, in quel caso la viscosità scende; in effetti gli individui anemici raggiungono più
facilmente il numero di Reynolds proprio perché viene a mancare il fattore denominatore; questo
determina la presenza spesso di turbolenze nel loro apparato cardiocircolatorio, che si manifestano
sotto forma di soffi, gli anemici "soffiano" molto più delle persone sane). Quindi quando il flusso
diventa da laminare a turbolento ci ritroviamo spesso in una condizione patologica, e le cause che
fanno aumentare il numero di Reynolds possono essere, ad esempio, un aumento del raggio del
condotto, cioè quando il vaso si dilata, anche se la dilatazione del vaso viene compensata in parte
dalla diminuzione conseguente della velocità di scorrimento; viceversa quando il vaso si costringe,
è vero che questo tenderebbe a diminuire il numero di Reynolds, ma l'aumento della velocità è
maggiore rispetto alla percentuale di restringimento del vaso. Per cui i vasi che soffiano di più non
sono quelli dilatati ma sono quelli stenotici: per capirci, se io a livello cardiaco ho una stenosi della
valvola aortica, uno dei primi segni medici che troviamo è il soffio sistolico, cioè il sangue soffia,
passa da un moto laminare a un moto turbolento, nel passaggio attraverso la valvola che si è ridotta
di calibro.
Il significato dei soffi cardiaci è quello: i soffi sono rumori generati dal flusso di sangue
turbolento attraverso vasi o valvole, in questo caso, di calibro ridotto. Altro esempio che vi
posso fare è il soffio carotideo, soffio che si sente alla base del collo, nei soggetti che hanno stenosi
carotidea (soprattutto soggetti anziani, che hanno avuto ad esempio l'ictus, hanno le carotidi che
progressivamente si restringono); noi poggiamo il fonendoscopio sul collo, in corrispondenza della
carotide: in una persona normale non soffia, in una persona che ha un restringimento lo fa. Quindi il
numero di Reynolds è importante perché ci dice quando il sangue passa da un moto laminare a un
moto turbolento, e, a parità di densità e a parità di viscosità, vediamo normalmente
nell'aumento della velocità di scorrimento la causa dei soffi.
Questo fenomeno viene sfruttato per la misurazione della pressione arteriosa. Sfrutta il fatto
che, se io causo un restringimento del vaso, in questo caso dell'arteria brachiale, in quella arteria si
generano dei flussi turbolenti, che vengono percepiti mettendo il fonendoscopio, come nel soffio.
Come si misura la pressione arteriosa: si tratta di applicare il manicotto pneumatico sul
braccio normalmente, ma si può applicare dovunque vogliamo, anche alla gamba, alla caviglia, alla
coscia, dovunque ci sia la possibilità di mettere un manicotto; il manicotto è collegato con un
sistema di pompaggio e anche con un sistema manometrico di misurazione della pressione; si gonfia
lo sfigmomanometro, in questo modo si comprime e si azzera la circolazione, si causa una
ischemia; a questo punto, la pressione del manicotto è tale da vincere la pressione arteriosa, per cui
se noi ci mettiamo ad ascoltare sotto (a valle del manicotto) non sentiamo niente perché non c'è
flusso, cioè il sangue non passa più; via via apriamo lo sfigmomanometro, facciamo uscire un po’ di
aria, si abbassa la pressione all'interno del manicotto fino a un punto tale per cui l'arteria si apre solo
un pochettino, e consente il passaggio del flusso in sistole, cioè quando la pressione arteriosa è
massima, e noi lo avvertiamo come un rumore, sentiamo "tuc" nel nostro fonendoscopio, ed è un
rumore generato da un flusso di sangue turbolento attraverso un vaso stenotico. E' quindi un flusso
di sangue molto veloce. Continueremo a far uscire aria dal nostro manicotto e continueremo a
sentire, in maniera sempre più forte, questi toni cardiaci, e arriveremo fino a un punto in cui il vaso
viene completamente aperto, cioè la pressione del manicotto non è tale da occludere in nessun modo
il vaso, cioè da causare stenosi nel vaso, e a quel punto non sentiremo più niente. La pressione
arteriosa sistolica, o pressione arteriosa massima la avvertiamo quando noi sentiamo il primo tono,
il primo rumore generato dal flusso di sangue attraverso il vaso stenotico, e la pressione diastolica
o minima c'è nel momento in cui non sentiamo più niente, cioè nel momento in cui non avvertiamo
più la turbolenza. E' una delle cose più semplici e utili da rilevare, ci dice subito qual è lo stato del
sistema cardiocircolatorio. I toni sono detti di Korotkoff (dal nome del primo che li descrisse).
(I fase: toni scoccanti, alla loro comparsa corrisponde il valore della pressione arteriosa
sistolica
II fase: toni soffianti
III fase: toni sordi
IV fase: toni affievoliti
V fase: scomparsa dei toni, qui si rileva la pressione diastolica)
Egli individuò 4 toni in questo sistema di auscultazione della frequenza cardiaca: il primo
tono, che è quello che abbiamo detto prima, quello che per primo sentiamo quando molliamo il
manicotto, poi toni che diventano più forti, poi meno palpabili e poi il quarto tono è quando non
sento. La pressione sistolica è il rimo tono di Korotkoff, la pressione diastolica è il quarto, cioè
quando non sento più niente, quando il vaso si è dilatato e il flusso non è più turbolento.
MICRO-CIRCOLAZIONE
A livello degli organi la circolazione è rappresentata dalla circolazione capillare, i quali hanno
grandissima sezione, e questo fa in modo che il flusso diventi estremamente lento e che il tempo di
contatto fra sangue e tessuti sia abbastanza dilungato e sia tale da consentire il passaggio di
metaboliti e gas nelle due direzioni. E' molto importante, come dicevo prima, che prima di questi
letti capillari ci siano degli sfinteri pre-capillari che, ricchi di tessuto muscolare liscio, regolano
l'afflusso di sangue nei vari distretti sulla base di segnali locali.
E' anche molto importante notare che i capillari sono formati da cellule endoteliali che sono
oggetto di intensa ricerca negli ultimi anni perché regolano un gran numero di funzioni
dell'organismo: l'endotelio dei vasi è l'organo più esteso dell'organismo, cioè se noi mettessimo
per terra tutto l'endotelio che abbiamo e facessimo un paragone con tutti gli altri tipi di tessuti,
risulterebbe essere il tessuto più esteso dell'organismo. Quindi è un tessuto di una importanza
straordinaria perché regola una grande quantità di eventi, dallo scambio di sostanze nutritizie tra
sangue e tessuti al tono dei vasi eccetera. E' spesso fenestrato perché deve consentire il passaggio di
sostanze, a volte anche di proteine da una parte all'altra dei vasi sanguigni.
I fattori che determinano il movimento dei fluidi attraverso i capillari sono identici a
quelli che regolano la filtrazione e il riassorbimento nel rene. C'è una pressione all'interno del
capillare (Pc) che tende a far uscire i flussi, come il plasma, tutto ciò che passa attraverso il
capillare fenestrato, e insieme a questa forza c'è la pressione osmotica dovuta alle proteine
presenti nei liquidi interstiziali (π IF); a queste due forze si oppongono la pressione idrostatica
che c'è a livello del tessuto (PIF ) (meglio del liquido interstiziale, a mio parere) e la forza
osmotica delle proteine presenti dentro il vaso (πC). Se vi ricordate la VFG del rene, è la stessa
cosa, le forze sono le stesse. Quindi la pressione netta di filtrazione è data dalla risoluzione di
questa semplice equazione (vedi nell'immagine al centro), dove sommiamo le forze che la
favoriscono e quelle che la ostacolano. E' importante notare che i capillari hanno un "lato" arterioso
che diventa via via venoso. Dalla parte del lato arterioso la pressione media a livello dei capillari è
normalmente intorno ai 35 mmHg, mentre la pressione osmotica è di 28 mmHg e tale rimane
durante tutto lo scorrimento del sangue; viceversa, la pressione capillare tende a diminuire, per
effetto dell'uscita di liquidi e per effetto anche della resistenza che il capillare stesso oppone allo
scorrere del fluido, e quindi all'estremità venosa troviamo una pressione capillare di 15 mmHg. C'è
una caduta di pressione di ben 20 mmHg, quindi questa forza che all'inizio del capillare diventa
ora di circa 10 mmHg (35, che è la pressione capillare, più 3 che è la pressione esercitata dalle
proteine dell'interstizio, meno 0 che è la pressione a livello interstiziale nell'estremità arteriosa,
meno 28 che è la pressione oncotica delle proteine a livello del plasma, totale 10 mmHg), poi
andando all'estremità venosa, si riduce la pressione capillare, tutto il resto più o meno rimane
costante, otteniamo -10 mmHg. Ciò vuol dire che dal lato arterioso il plasma esce verso il tessuto,
dal lato venoso il plasma tende a rientrare verso il vaso, per cui a livello arterioso il vaso tende un
pò a disidratarsi, a perdere liquidi, a perdere plasma, mentre a livello venoso tende a reidratarsi,
quindi a riassorbirlo. Questo movimento determina da un lato il passaggio di sostanze nutritizie dal
vaso all'interstizio per gradiente di concentrazione, dall'altro lato il passaggio di sostanze di scarto
dall'interstizio al vaso sempre per gradiente di concentrazione e favorito anche dalla riduzione della
pressione all'interno del vaso. Questo sistema si può controllare,e gran parte del controllo si attua
controllando la pressione del capillare attraverso questa equazione detta di Pappenheimer :
PC= [PA (rV/rA) +PV]/[1+ (rV/rA)]
Cosa dice questa equazione: che la pressione (PC) dipende dalla pressione arteriosa (PA), per il
rapporto tra resistenze a livello venoso (rV) e resistenze a livello arterioso (rA), più la pressione
venosa (PV), il tutto fratto per 1 più il rapporto tra resistenze venosa e arteriosa.
Alla fine il modo con cui si può regolare tutto è variando le resistenze venose e quelle
arteriose e lasciando costante la pressione arteriosa e quella venosa. Come posso variare le
resistenze? Attraverso gli sfinteri pre-capillari. Gran parte del controllo della circolazione locale
nei vari tessuti si attua modificando il rapporto rV/ rA
CELLULE ENDOTELIALI
Fungono da rivestimento, la cosa più logica, ma i globuli rossi non aderiscono (se succede si
forma un trombo, una delle cause di formazione di trombi, che portano per esempio a infarto
cardiaco, ictus, è proprio la formazione di coaguli laddove non ci dovrebbero essere, e spesso la
presenza del coagulo è dovuta al fatto che il rivestimento dei vasi viene alterato, come nella
aterosclerosi, nella quale si riduce il calibro del vaso e fa molto male agli endoteli che ci sono
dentro, che perdono la loro funzione, smettono di rivestire, o lo fanno male, e laddove si perde il
rivestimento si creano tutte le condizioni per far formare un coagulo, cioè un trombo)
Fungono da filtro di permeabilità per lo scambio di nutrienti, cioè regolano lo scambio di
nutrienti tra il capillare ed il tessuto; secernono una marea di ormoni ad azione locale, il più
famoso probabilmente è l'NO. Il monossido di azoto è il più importante vasodilatatore endogeno
che abbiamo (quando ai malati di angina si da la pastiglia sotto la lingua, essa serve ad aumentare le
concentrazioni di NO a livello della circolazione coronarica, per provocare vasodilatazione), ma
anche altri come le prostacicline, e vasocostrittori come l'endotelina (quello in tabella, ovviamente,
è un riassunto, di prodotti ce ne sono a decine);
mediano l'angiogenesi, cioè la crescita di nuovi capillari, cosa che si è rivelata fondamentale
nelle ultime ricerche sul cancro, ad esempio, dove si cerca di portare a ipossia i tessuti cancerosi
evitando che si formino nuovi vasi sanguigni al loro interno. I tessuti cancerosi infatti hanno
bisogno di molti metaboliti, e senza l'apporto dei vasi sanguigni, muoiono;
giocano un ruolo centrale nel rimodellamento vascolare, fungono da mediatori nell'azione
paracrina delle cellule vicine, cioè fanno da tramite nello scambio di informazioni tra cellule del
vaso e cellule dei tessuti;
formano la matrice extracellulare;
producono fattori di crescita in risposta a un danno, importante perché una delle cause
dell'aterosclerosi (deposito di materiale come colesterolo nella parete del vaso) è che le cellule
endoteliali, in risposta a un danno di parete, si mettano a produrre fattori di crescita che tendono a
inglobare il colesterolo, e questo contribuisce alla chiusura del vaso stesso;
secernono sostanze che regolano l'aggregazione piastrinica, e quindi la coagulazione (un
danno endoteliale favorisce la coagulazione);
sintetizzano ormoni attivi;
estraggono mediatori dal sangue e li trasferiscono;
secernono citochine, e quindi entrano in gioco anche nella risposta immunitaria;
influenzano la proliferazione delle cellule muscolari lisce durante l'aterosclerosi, in
risposta a un danno di parete.
29-03-12
prof. Crisafulli
Lorenzo Steri
CONTROLLO NERVOSO DEL CIRCOLO
La circolazione come tute le funzioni dell’organismo è controllata, nel senso che il sistema
circolatorio risponde a perturbazioni di stato, dell’omeostasi, con determinate risposte governate a
loro volta da una serie di fattori il più importante dei quali è sicuramente il SN. Come gran parte di
quello che succede nell’organismo il SN e quello ormonale regolano la risposta di vari organi.
Esempio classico del controllo sul sistema circolatorio è la risposta all’esercizio, dove si assiste
all’aumento dei battiti cardiaci.
Il cuore è controllato in gran parte delle sue funzioni dal SNA, simpatico e parasimpatico
(frequenza cardiaca, contrattilità, inotropismo, cronotropismo, ecc.). Il parasimpatico, attraverso i
nervi vaghi e con mediatore chimico acetilcolina, innerva gli atri (soprattutto), il nodo SA, la
muscolatura atriale, il nodo AV, ecc. ma non innerva i ventricoli. Viceversa il sistema simpatico
innerva sia atri che ventricoli, ma in maniera diffusa, non solo con un innervazione diretta (con
nervi) ma anche attraverso altri sistemi quale l’adrenalina, che viene rilasciata nel sangue dalle
ghiandole surrenali e può innervare il cuore, tramite quindi un mediatore umorale (l’adrenalina non
è un mediatore nervoso in senso stretto).
I nervi che arrivano al cuore derivano, per quanto riguarda il sistema ortosimpatico, dai
segmenti toracici da T1 a T5, arrivano dalla catena gangliare paravertebrale, formando una lunga
catena di gangli simpatici da cui poi si dipartono tre nervi principali che sono i nervi cardiaco
superiore, medio e inferiore. Essi seguono i vasi sanguigni, l’aorta, e arrivano al cuore.
Per quanto riguarda invece il parasimpatico l’innervazione è costituita dal nervo vago.
L’innervazione non è fondamentale per il funzionamento cardiaco. L’esempio fondamentale di tale
condizione è il trapianto, quando il cuore è trapiantato infatti viene denervato. C’è tuttavia la
possibilità che si riformi in parte l’innervazione, ma di fatto il cuore funziona anche senza.
Ovviamente il cuore denervato non è un cuore normale, nel senso che non risponde come un cuore
normale alle variazioni di stato. Per esempio un cuore denervato ha una frequenza cardiaca di base
un po’ più alta, intorno ai 100-110 battiti/min, risponde meno prontamente a un esercizio: mentre in
una persona normale sale subito il battito, il cuore denervato ci impiega più tempo e gran parte delle
variazioni si ottengono grazie all’adrenalina del corrente circolatorio, non più tramite i nervi.
AZIONI DEL PARASIMPATICO
Le azioni parasimpatiche fondamentali sono: effetto cronotropo negativo, inotropo negativo
(sugli atri) e dromotropo negativo.
Il nervo vago essenzialmente ha un effetto cronotropo negativo, cioè rallenta la frequenza
cardiaca. In condizioni basali prevale il tono parasimpatico e il nostro cuore se denervato ha una
frequenza intorno ai 100 battiti/min, quando invece siamo a riposo la frequenza cardiaca è intorno ai
70 battiti/min, il che vuol dire che abbiamo un tono parasimpatico di base.
Ha poi effetto inotropo negativo, cioè la forza di contrazione del cuore si riduce, ma solo
sugli atri (il parasimpatico non innerva i ventricoli).
C’è poi un effetto dromotropo negativo (dromotropismo: capacità di condurre l’impulso che
rallenta la conduzione dell’impulso all’interno del cuore). Si ottiene un rallentamento tramite una
scarica vagale: come sappiamo nell’ECG si osserva l’intervallo PR, dall’inizio dell’onda P
(contrazione atriale) all’inizio del complesso QRS (contrazione ventricolare), una scarica vagale
allunga proprio questo intervallo. È la testimonianza che c’è un allungamento del tempo di
conduzione all’interno dei circuiti di conduzione del cuore.
I meccanismi d’azione son diversi a dipendenza del tipo di struttura (del nodo SA, della
muscolatura atriale, del nodo AV e via dicendo). Nel nodo SA un aumento delle cariche
parasimpatiche causa un aumento della conduttanza al potassio con un'iperpolarizzazione. Quello
che succede è praticamente questo:
questa è la condizione A normale (ricorda queste cellule si depolarizzano spontaneamente
raggiungendo il potenziale soglia), se viene fornita una scarica parasimpatica di acetilcolina la
depolarizzazione avviene ugualmente, ma cambia la pendenza di questa fase di depolarizzazione
spontanea che avviene con un certo rallentamento, per cui la frequenza cardiaca rallenta. Quest’altra
è una scarica simpatica che fa il contrario, cioè aumenta la velocità del battito cardiaco. A livello del
nodo AV c’è una riduzione dell’ampiezza del potenziale d’azione che rallenta proprio la velocità
con la quale questo potenziale d’azione viene trasmesso, quindi rallenta la velocità con cui il cuore
trasmette gli impulsi al suo interno. Nelle cellule atriali c’è proprio una riduzione della durata del
potenziale d’azione. Nella figura si vede, con la linea piena, il cuore “normale” e sotto un’altra linea
tratteggiata, quello che succede quando c’è nel potenziale d’azione delle varie cellule un’azione del
parasimpatico. Quindi a livello dell’atrio si osserva una riduzione della durata del potenziale
d’azione, poi a livello della giunzione una situazione simile, poi a livello delle cellule ventricolari
non succede niente. Come già detto il parasimpatico non agisce sulle cellule ventricolari e neanche
sul fascio di His.
AZIONI DEL SIMPATICO
La branca del simpatico svolge una serie di altre azioni, opposte in linea di massima a quelle
del parasimpatico: effetto cronotropo positivo, inotropo positivo, dromotropo positivo e batmotropo
positivo.
Effetto cronotropo positivo significa che aumenta la pendenza della fase di depolarizzazione
spontanea delle cellule pacemaker e quindi la frequenza cardiaca aumenta. Queste cellule si
depolarizzano spontaneamente in maniera più veloce rispetto al normale.
Ha poi effetto inotropo positivo, aumentando la forza di contrazione del cuore. Il cuore si
spreme con più violenza, con più forza, e questo normalmente si vede. Il cuore in questo caso passa
da una frazione di eiezione (esposta nella lezione precedente) a riposo del 60% a una del 70-80 fino
a 90%.
Effetto dromotropo positivo significa che aumenta la velocità di conduzione degli impulsi
(questo è coerente con l’aumento del cronotropismo), cioè il cuore batte più velocemente e gli
impulsi devono essere condotti più velocemente per coordinare l’attività cardiaca.
Ha effetto batmotropo positivo, cioè aumenta l’eccitabilità cellulare (batmotropismo:
sensibilità o eccitabilità delle fibre muscolari cardiache allo stimolo di contrazione). Questo è un
effetto tipico del simpatico e può avere anche delle conseguenze negative, perché aumentando
l’eccitabilità cellulare può scatenare dei battiti prematuri, cioè può rendere alcune cellule pacemaker
anche se normalmente non dovrebbero esserlo. Si chiamano extrasistoli e sono problemi abbastanza
comuni. In pratica prende il comando del ritmo cardiaco un gruppo di cellule che normalmente non
lo dovrebbe fare, perché viene eccitato in maniera anormale da una scarica del sistema simpatico.
L’extrasistolìa è anche un effetto dello stress cronico, i battiti prematuri avvengono prima di quando
dovrebbero presentarsi, in quanto non nascono dal nodo SA ma da altre cellule.
Osservando l’immagine in basso della slide si apprezza lo sviluppo della forza di contrazione
durante una contrazione cardiaca, in una situazione di scarica simpatica e una condizione di
controllo normale.
C’è un aumento di capacità di generare forza in gran parte dovuta all’aumento di calcio
intracellulare che permette i ponti actina-miosina, i quali aumentano nelle cellule muscolari
cardiache e quindi aumenta anche la forza di contrazione. Il meccanismo d’azione del nodo SA è
una riduzione della conduttanza al Ca e al K che depolarizza le cellule e quindi rende più facile il
raggiungimento del potenziale soglia.
Sono riassunti i principali effetti del sistema orto- e parasimpatico (esistono altri tessuti oltre
quelli elencati come il fascio di His che sono qui omessi).
Strutture coinvolte nel controllo nervoso del sistema cardiocircolatorio.
Il SNA regola le funzioni del cuore sulla base di una marea di informazioni che arrivano a
livello di centri nervosi appartenenti al SNC che informano sullo stato del sistema
cardiocircolatorio. Ad esempio informano se c’è pressione troppo alta, pressione troppo bassa, se
manca ossigeno, se c’è troppa anidride carbonica, ecc. Il tutto è reso possibile grazie a recettori che
regolano l’apparato cardiocircolatorio i principali dei quali sono: barocettori aortici e carotidei,
recettori cardiaci, recettori polmonari, chemocettori e recettori al di fuori del sistema
cardiocircolatorio (ne esistono anche altri).
Questa serie di recettori che informano il SNC dello stato di alcune condizioni in periferia
(pressione, ecc.) informano i centri cardiaci che sono sparsi in vari distretti del SN. In particolar
modo c’è un centro spinale, le colonne intermedie-laterali del midollo spinale, centri bulbari, cioè
d’integrazione del segnale situati nel midollo allungato, dove c’è il nucleo del tratto solitario e le
parti caudali e rostrali e ventro-laterali del bulbo. Nel bulbo ci sono infatti dei centri diffusi, zone di
controllo cardiocircolatorio. Sezioni di queste regioni su animali hanno reso note le loro funzioni, in
quanto si distrugge il centro e si va a vedere cosa provoca, quindi non esperimenti sull’uomo.
Questi centri bulbari sono proprio i veri centri di raccolta da cui partono i riflessi. Arrivano
una marea di informazioni da ipotalamo, aree motorie, insula, oliva, importanti perché informano il
sistema bulbare su quello che stiamo facendo. Cioè l’area motoria dice ai centri bulbari che distretto
corporeo muoviamo in un dato momento e i centri bulbari recepiscono le informazioni e modulano
il ritmo cardiocircolatorio sulla base di queste informazioni.
La slide 9 è uno schema di quelle che sono le connessioni dei centri bulbari nel midollo
allungato con le varie informazioni che gli arrivano sia dalla periferia che dal centro (nella realtà
non è così semplice come nello schema). Questi centri ricevono informazioni da pressocettori
arteriosi e venosi, che dicono quant’è la pressione nei vari vasi della periferia, ma non solo, vi sono
anche recettori muscolari (che vedremo in una lezione a parte) che informano i centri
cardiocircolatori di cosa avviene nel distretto muscolare (se i muscoli hanno bisogno di ossigeno, se
c’è troppa anidride carbonica, se si stanno accumulando metaboliti e via dicendo). Altre afferenze ai
centri bulbari sono quelle dai chemocettori che informano su quanto ossigeno è presente nel sangue
e via dicendo. Questi centri ricevono anche informazioni dalla corteccia, in particolare dall’area
motoria e dal sistema limbico, quindi vengono informati del nostro stato emozionale, di quello che
stiamo facendo, ecc., dal cervelletto che controlla essenzialmente i movimenti, ma anche
dall’ipotalamo, grazie al quale vi sono una marea di informazioni sulla sete sulla fame sulla vita
vegetativa in generale (importante è il controllo della concentrazione di glucosio). Questi centri
sanno tutto in pratica e in base alle informazioni e dello stato dell’organismo organizzano la risposta
cardiocircolatoria (aumentano la frequenza cardiaca, la pressione, ecc.), tramite ovviamente il
sistema simpatico e parasimpatico.
RIFLESSI BAROCETTIVI
Sono i riflessi più studiati e i più semplici da studiare. I barocettori aortici e carotidei sono
piccoli organelli posti in zone strategiche, in particolar modo sull’arco aortico e sulla biforcazione
dell’arteria carotide comune quando si biforca in carotide interna ed esterna. Son lì situati per via
del ruolo dell’aorta nel nostro organismo, sanno esattamente qual è la pressione dell’organismo.
Quelli situati sulla diramazione della carotide devono proteggere l’organismo da riduzioni
pressorie, ma soprattutto devono proteggere il cervello, organo che più di tutti ha bisogno di un
flusso ematico costante e che meno di tutti è in grado di resistere all’ipossia o alla mancanza di
nutrienti. Il cervello di un adulto solitamente resiste tre quattro minuti senza sangue, dopodiché
muore. Altri organi invece resistono molto di più, per esempio il cuore può rimanere senza sangue
per molto tempo. Per un trapianto viene asportato, poi viene ibernato, nel senso che viene messo a
contatto con il ghiaccio e infine viene messo nel torace di chi lo riceve. Ci mettono ore, 6-7 ore però
resiste. Quindi questi organelli son messi qua per proteggere il cervello da qualsiasi variazione di
pressione arteriosa, in particolar modo per evitare che ci sia una riduzione del flusso a livello
dell’encefalo. Di barocettori ne esistono anche altri ma i più importanti sono sull’arco aortico e sulla
biforcazione delle carotidi.
Al centro dell’immagine abbiamo una situazione di normalità, 100, a sinistra pressione più
bassa, pressione media 70 per esempio, a livello del nodo del seno carotideo le
cellule sentono questa riduzione della pressione e riducono la loro frequenza di scarica.
Siccome sono proprio dei meccanocettori, rispondono allo stiramento e la riduzione di pressione
riduce lo stiramento e loro riducono la loro frequenza di scarica. La riduzione della frequenza di
scarica viene percepita dai centri analizzati precedentemente e contemporaneamente si ha un
aumento della frequenza cardiaca che è dovuto a un aumento delle scariche del simpatico (effetto
cronotropo positivo) e inoltre una contemporanea riduzione delle scariche del nervo vago, cioè del
parasimpatico, rispetto a una situazione normale.
Questi picchi che si osservano sono le scariche proprio di queste cellule. Nella situazione
normale c’è un abbassamento di pressione, riduzione, quindi scariche del nervo vago e aumento
delle scariche del simpatico con incremento della frequenza cardiaca. In una situazione opposta,
aumento di pressione (per qualsiasi motivo), queste cellule aumentano la loro frequenza di scarica,
vengono stirate, e l’effetto è l’opposto di quello visto prima, cioè diminuzione della frequenza
cardiaca, aumento delle scariche del parasimpatico e riduzione delle scariche del simpatico. Perché
viene aumentata la frequenza cardiaca? Allora: frequenza normale, frequenza cardiaca 70, la
pressione viene ridotta, la frequenza cardiaca diventa 90. Questo per aumentare la gittata cardiaca,
perché la pressione è uguale a (legge di Poiseuille) “gittata cardiaca x resistenze vascolari
periferiche”, a parità di resistenze se si aumenta la gittata cardiaca aumenta la pressione. Situazione
opposta: perché a fronte di un aumento della pressione arteriosa io riduco la frequenza cardiaca? Per
il motivo inverso. L’effetto è simile sulla contrattilità cardiaca, nel senso che in una ridotta
pressione arteriosa causa un aumento, sempre attraverso fibre simpatiche, della contrattilità
cardiaca, quindi il cuore si svuota più efficacemente.
Il contrario di fronte a un aumento della pressione arteriosa: c’è bisogno di aumentare la
contrattilità cardiaca se si abbassa la pressione arteriosa in quanto bisogna aumentare la gittata
sistolica. In realtà poi aumenta anche la gittata cardiaca, sempre perché la pressione è uguale a
gittata cardiaca x resistenza. In caso di pressione arteriosa elevata vale il contrario (tenete presente
che però gran parte della modulazione avviene sulla frequenza cardiaca più che sulla contrattilità).
Questo riflesso funziona non solo a livello cardiaco, ma anche a livello dei vasi, sia di resistenza
(cioè le arteriole) sia di capacitanza (cioè le vene).
Se si ha vasocostrizione aumento le resistenze periferiche quindi, rispettando la legge:
pressione = gittata cardiaca x resistenza, si ha un aumento della pressione. Si può quindi modulare
la pressione arteriosa modulando la frequenza cardiaca, la gittata sistolica e le resistenze. Giocando
su questi tre parametri il mio organismo, i miei centri cardiocircolatori, sono in grado di regolare la
pressione arteriosa. Se i vasi si chiudono, aumenta la resistenza e i vasi di capacitanza si chiudono
anch’essi.
Perché le vene si possono vasocostringere? Le vene contengono il 60% del sangue,
supponiamo ora che se ne vada il 2%. Supponiamo poi di avere 5L di sangue in totale, il cuore e le
vene ne contengono 3L. Se io porto via il 2% solo in quei 3L corrispondono a 60mL che vanno a
finire verso il cuore e aumentano la gittata sistolica per aumento del volume di fine diastole o del
precarico cardiaco. Siccome normalmente il volume della gittata sistolica è intorno ai 70-80 mL, se
si portano al cuore altri 60 mL si sta quasi raddoppiando la gittata sistolica. Basta una piccolissima
vasocostrizione per aumentare di molto il ritorno venoso al cuore e quindi indurre il cuore ad
aumentare la propria gittata sistolica. Ecco allora il significato della scarica del simpatico:
modulazione della gittata cardiaca e modulazione delle resistenze vascolari periferiche e del ritorno
venoso al cuore. in casi di pressione elevata accade tutto il contrario, cioè predomina il
parasimpatico, quindi riduzione della frequenza, del ritorno venoso, delle resistenze e della
contrattilità.
RECETTORI CARDIACI
La pressione arteriosa è il parametro più importante che viene regolato dai sistemi
cardiocircolatori. Il primo obbiettivo dell’organismo è mantenere la pressione arteriosa stabile. La
modulazione avviene anche tramite recettori cardiaci, posti dentro il cuore stesso, di cui ce ne sono
svariati (il prof ne ha riassunto qui alcuni). L’importante è sapere che nel cuore ci sono dei
volocettori e pressocettori, distribuiti sia negli atri che nei ventricoli. I volocettori registrano volumi,
quanto il cuore si gonfia, i pressocettori registrano pressioni, quelle dentro le camere cardiache.
Questi recettori segnalano lo stato di riempimento delle camere e la pressione che si genera al loro
interno. Esistono poi dentro il cuore dei recettori termici e dei recettori meno importanti, che
possono assumere interesse in condizioni patologiche. Per esempio quando si iniettano determinate
sostanze nel cuore si possono generare riflessi parasimpatici che bloccano il cuore, mediati da tali
recettori.
Aumentando il riempimento degli atri (soprattutto) si attivano, per via riflessa, questi
volocettori. Esistono anche nel ventricolo ma son molti meno e molto meno efficaci, quelli
importanti sono negli atri e questi, sempre per via riflessa, attraverso i centri bulbari, sono in grado
di indurre: vasodilatazione renale con aumento di velocità di filtrazione glomerulare e riduzione
della produzione di ormone antidiuretico. Cioè poniamo che si abbia un sovracarico di liquidi, sono
ricoverato in ospedale e il medico ha sbagliato e mi hanno dato 2 L invece che 0.5 L di liquidi, che
vanno a finire nel torrente circolatorio. Questo aumenta il precarico cardiaco, cioè aumenta il
volume di fine diastole cardiaca. Il cuore si gonfia letteralmente, l’atrio si gonfia, questo
sovraccarico di liquidi viene recepito da questi recettori, i quali informano i centri bulbari per via
riflessa e influenzano la frequenza cardiaca. Questo avviene per smaltire il più velocemente
possibile il sovraccarico di liquidi che arriva al cuore, è un meccanismo di espulsione rapida di
liquidi dal cuore alla periferia, per evitare che poi si ingorghi.
Una delle conseguenze degli ingorghi del cuore si manifesta poi come edema polmonare (una
delle condizioni più difficili per il medico e anche molto frequente). Quando si ha l’edema
polmonare il cuore viene sovraccaricato oppure non funziona bene, si ingorga il circolo polmonare,
il fluido esce dai polmoni che smettono di funzionare. Uno di questi meccanismi di difesa da questa
situazione è aumentare la frequenza cardiaca in modo tale che smaltisca la quantità di liquidi.
Contemporaneamente c’è anche una vasodilatazione del distretto renale con aumento della
velocità di filtrazione glomerulare. Un aumento del calibro dei vasi renali aumenta la pressione a
livello dei capillari che entrano nel glomerulo e questo aumenta la capacità di filtrazione
glomerulare. Questo fa in modo che il sovraccarico di liquidi venga smaltito dal rene. Se non c’è il
corretto funzionamento del rene permane il pericolo del sovraccarico di liquidi.
Contemporaneamente c’è una diminuzione della secrezione dell’ormone antidiuretico, cioè il dotto
collettore non è più permeabile all’acqua, per cui passando attraverso quel gradiente di
concentrazione nella midollare del rene, non si mette più in equilibrio con quel gradiente osmolare,
per cui le urine vengono escrete in maniera non concentrata (aumento della diuresi). L’organismo
cerca di eliminare questo sovraccarico di liquidi con la diuresi.
Poi ci sono i pressocettori che registrano a livello del cuore il sovraccarico di pressione e
hanno funzioni analoghe ai pressocettori arteriosi analizzati precedentemente. Questi a fronte di un
aumento della pressione aumentano la frequenza cardiaca e causano una vasocostrizione (come i
barocettori aortici e carotidei). Il sovraccarico di volume, oltre a influire su questi volocettori,
induce anche i miociti stessi a secernere un ormone, il peptide natriuretico atriale, anche questo
abbastanza importante. Quest’ultimo è un ormone cardiaco che induce nel rene l’eliminazione di
sodio e quindi di acqua, un altro meccanismo per informare il rene quando vi è un’alta quantità di
liquidi e che stimola la loro espulsione.
I recettori precedenti sono i più efficaci, quelli che seguono invece non agiscono in maniera
prepotente e sono perciò meno importanti.
RECETTORI POLMONARI
Ci sono dei recettori, detti polmonari, localizzati nelle arterie polmonari che rispondono
anch’essi allo stiramento. L’aumento di pressione a livello polmonare ha soprattutto un effetto
ventilatorio, ma può anche dare, quando questo aumento di pressione polmonare è intenso, una
riduzione della frequenza cardiaca con … ed è un tentativo di limitare incrementi della pressione
arteriosa polmonare. Questi sono riflessi importanti non tanto in fisiologia quanto in patologia,
esistono condizioni di pressione arteriosa cronica, dove si possono avere anche ripercussioni a
livello cardiaco.
CHEMOCETTORI
Oltre a quelli visti in aorta e carotide, esistono anche dei chemocettori, recettori che registrano
la composizione chimica del sangue. Questi sono in particolare molto interessati a sapere quanto
ossigeno abbiamo nel sangue, sono molto sensibili alla pressione di ossigeno. Sono messi vicino ai
pressocettori, negli stessi punti nei pressi dell’arco aortico e si chiamano glomi aortici e nella
diramazione della carotide comune, glomi carotidei (ne esistono anche altri ma questi sono i
principali). La risposta riflessa alla riduzione della pressione parziale di ossigeno è una bradicardia
con una riduzione della gittata cardiaca, cioè riducono la frequenza cardiaca e riducono la gittata
cardiaca e quindi il lavoro cardiaco. In sostanza la richiesta di ossigeno da parte del cuore è un
tentativo per difendere il cuore da eccessive deduzioni di pressione di ossigeno riducendo il lavoro.
Registrano se ho una riduzione della pressione di ossigeno: ad esempio salendo in montagna,
dove c’è un abbassamento della pressione dell’aria, questa riduzione viene percepita dai glomi che
dicono al cuore che deve rallentare il proprio lavoro. Un’altra risposta riflessa a riduzioni della
frequenza cardiaca sono l’aumento delle resistenze con aumento anche della pressione arteriosa e,
cosa importante, inducono una vasodilatazione a livello del circolo coronarico, le arterie del cuore si
aprono e favoriscono un maggior afflusso di sangue.
RECETTORI EXTRACIRCOLATORI
Poi esistono una marea di recettori sparsi nel nostro organismo che sono in grado di
influenzare i centri nervosi di controllo cardiocircolatorio e di indurre risposte riflesse, in particolar
modo dolorifiche. C’è gente che sviene per dolori intensi, che altro non sono che intense scariche
vagali, le quali determinano una vasodilatazione periferica e un rallentamento della frequenza
cardiaca tale da determinare una profonda caduta della pressione arteriosa con svenimento. C’è da
notare che a fronte del dolore non tutti rispondono allo stesso modo. C’è un gruppo più numeroso di
persone che risponde con una diminuzione della pressione arteriosa e un gruppo meno numeroso di
persone che risponde al contrario, con un aumento della pressione arteriosa, con una scarica
simpatica più che vagale.
Fibre tattili e meccanocettori cutanei: noi possiamo avere risposte pressorie anche
semplicemente con stimolazioni cutanee, oppure rispondiamo a un aumento di metaboliti a livello
dei muscoli ad esempio. Quando i muscoli si mettono a lavorare si accumulano acido lattico,
idrogenioni, adenosina, e c’è una risposta riflessa cardiocircolatoria, scatenata proprio da questi
riflessi. Ma ci sono anche i meccanocettori muscolari, cioè nei muscoli ci sono dei meccanocettori
che registrano lo stato di stiramento delle fibre e se io faccio ad esempio un tipo di lavoro
isometrico, dove i muscoli vanno in tensione, questi recettori registrano lo stato di contrazione
muscolare e scatenano risposte riflesse, cioè un scarica simpatica (aumento di frequenza cardiaca,
aumento di pressione, ecc.). Non solo, ma abbiamo anche molti recettori per gli organi di senso.
Stimoli visivi, olfattivi e importanti sono soprattutto quelli del senso dell’equilibrio a fronte di
variazioni proprio della porzione corporea, che possono portare ad aumento di funzioni circolatorie
come aumento di pressione, vasodilatazione e via dicendo. I piloti d’aereo che hanno intense
accelerazioni in fase di decollo o picchiata (soprattutto i piloti militari) e forti stimolazioni dei
labirinti possono dare stimolazioni intense parasimpatiche con svenimento. È un riflesso che parte
proprio dal senso dell’equilibrio.
[Il prof si è dimenticato di mettere anche Metabo- e meccanorecettori muscolari nell’elenco
dei recettori extracircolatori].
CONTROLLO NERVOSO DEI VASI
Anche nei distretti vasali c’è un controllo nervoso (ma non solo) e in linea di massima sui
nostri vasi vi sono un controllo vasodilatatore e vasocostrittore, dove il più importante è
quest’ultimo, perché normalmente i nostri vasi tenderebbero a essere vasodilatati. Nella persona
normale a riposo predomina il tono vasocostrittore, il contrario di quello che succede nel cuore,
dove predomina il tono parasimpatico (ricorda la frequenza cardiaca è più bassa che non la
frequenza intrinseca). Se noi avessimo un’interruzione dell’innervazione dei vasi avremmo una
vasodilatazione e normalmente non lo sono perché predomina un certo tono simpatico che li tiene
chiusi. Questo si può vedere in persone che soffrono molto di un forte traumatismo spinale, traumi
stradali ad esempio, con un’interruzione delle vie nervose. Quello che succede nella parte inferiore
del corpo, quella che viene colpita da questa paralisi, è un’intensa vasodilatazione perché viene a
mancare il tono simpatico di base. Quindi gran parte della modulazione del tono vascolare viene
fatta aumentando o riducendo la vasocostrizione. In sintesi, un vaso, è soggetto a una
vasodilatazione a cui si aggiunge il tono simpatico che la riduce di base e a queste due condizioni si
possono sommare stimoli provenienti dai centri di controllo ma anche da stimoli locali. I nervi
vasodilatatori infatti sono assai meno diffusi, esistono ma hanno significati particolari, sono sia
parasimpatici che simpatici. Quelli parasimpatici sono distribuiti soprattutto negli organi genitali e
alcune regioni cerebrali, hanno una funzione che non è tanto quella di preservare la pressione, ma
sono funzioni locali. Negli organi genitali sono proprio legati alla funzione sessuale, nei distretti
cerebrali sono dediti a funzioni cerebrali. Non hanno una grande importanza nell’economia generale
del sistema cardiocircolatorio, sono particolarità.
Poi esiste anche (secondo alcuni autori) un ortosimpatico colinergico, dei nervi simpatici che
però hanno acetilcolina come mediatore (è un’eccezione). Secondo alcuni sono sparsi nella
muscolatura scheletrica e determinano vasodilatazione a questo livello, ma non sono molto
importanti, anzi secondo alcuni non esistono proprio. Questi intervengono nelle zone cutanee,
soprattutto nelle reazioni emotive (tipo quando uno è spaventato → vasodilatazione). Anche questi
però non sono importanti nell’economia globale dell’organismo. Esistono poi altre particolarità, una
popolazione non ben definita di neuroni che non hanno come mediatore chimico né noradrenalina
né acetilcolina, non sono né simpatici né parasimpatici, ma che hanno come mediatore chimico
l’ossido d’azoto, NO, che sono importanti in alcune funzioni come ad esempio funzioni sessuali
legate alla vasodilatazione dei vasi del pene. Anche questi non sono importanti nell’economia
globale. Quelli fondamentali sono i vasocostrittori che modulano la vasocostrizione del nostro
organismo sulla base dei segnali che arrivano ai centri di regolazione cardiocircolatori. Anche per i
vasi, come per il cuore, esistono dei centri nervosi che sono in gran parte sovrapponibili a quelli
analizzati per ora, anche se quelli vasali sono un po’ più estesi, occupano un po’ più spazio nel
sistema nervoso. Nel midollo spinale i centri cardiaci sono a livello dei segmenti T1-T5, mentre
questi (quelli ortosimpatici) sono T1-L4. Occupano una lunghezza maggiore nel midollo spinale
proprio perché i vasi sono sparsi un po’ in tutto l’organismo e sono anch’essi situati nelle colonne
intermedie-laterali del midollo, ma anche nel tratto sacrale dei segmenti S2-S4 dei nervi del tipo
parasimpatico. Questi sono quelli che poi arrivano ai genitali, alla vescica urinaria e via dicendo.
Poi abbiamo il nucleo ambiguo da dove parte il nervo vago. Le fibre ortosimpatiche vasodilatatrici
(quelle che secondo alcuni non esistono neanche) non si sa bene da dove nascano almeno
nell’uomo.
Lo schema generale dell’innervazione simpatica dei vasi arteriosi e motori ripete quella del
cuore, cioè c’è il centro del bulbo vasomotore, dove arriva quella marea di informazioni da cui
partono informazioni per il midollo spinale per i nuclei intermedi-laterali dei vari segmenti spinali e
da qui poi partono i vari neuroni periferici che arrivano sia al cuore ma anche alle arteriole (in gran
parte) dove modulano la vasocostrizione ma anche alle vene dove modulano la veno-costrizione
cioè la possibilità di centralizzare il ….
Lo schema è uguale a quello del cuore, cioè centro e poi periferia, attraverso l’ortosimpatico e
il parasimpatico.
Nelle arterie la vasocostrizione si traduce in un aumento della pressione arteriosa, perché
viene modulato nell'equazione di Poiseuille (PA = GCπr^4/8ηl) questo fattore (πr^4), è determinato
da piccolissime variazioni del calibro delle arteriole, determinando grandi variazioni del valore
della funzione. Gittata cardiaca e 8ηl si chiamano resistenze vascolari periferiche che vengono
modulate soprattutto attraverso la modulazione del raggio medio arteriolare, cioè di quanto son
costrette o dilatate le nostre arteriole. Nelle vene invece l’azione del simpatico si esplica attraverso
una veno-costrizione e la funzione è portare volumi di sangue dalle vene verso il cuore.
C’è un esempio classico che si chiama esempio di Folkow, del ritorno venoso, fatto nel 1960
ma che vale tutt’ora. Riutilizzando i valori dell’esempio di prima con un volume di sangue ematico
di circa 5L che più o meno è quello di una persona normale, circa 3,25 L di sangue sono contenuti
nelle vene. Se riesco a causare una lieve venocostrizione, di 2,5%, riesco ad utilizzare il 2,5% di
3,25 L che sono circa 80 mL di sangue, che vanno verso il cuore. 80 mL di sangue è la gittata
sistolica, quindi raddoppio una gittata sistolica. Il cuore riceve questo volume di sangue e risponde
con un aumento del volume telediastolico, aumentando la gittata sistolica, ma come abbiamo visto
prima anche aumentando la frequenza cardiaca, cioè tentando di eliminare velocemente questo
volume di sangue e di portarlo in periferia.
DOMANDA riguarda le due slide precedenti: la parte vasodilatatrice del sistema simpatico,
non era data da diversi neurotrasmettitori, ma bensì dalla presenza di recettori diversi, che
funzionano sempre con l’adrenalina, nella muscolatura scheletrica.
RISPOSTA: allora tu avrai studiato che nella muscolatura ci sono recettori alfa e beta ed è
vero, però a quanto pare esistono anche dei recettori colinergici vasodilatatori, cioè come mediatore
chimico è proprio diverso, non solo, nei muscoli agisce oltre che la noradrenalina anche l’adrenalina
del surrene che ha anche un’azione vasodilatatrice invece che vaso costrittrice ma quello lo vediamo
la lezione prossima. Sono delle eccezioni, dei funzionamenti particolari del muscolo, dei vasi del
muscolo. Si è vero, però per alcuni esiste anche questo. Neuroni simpatici vasodilatatori con
mediatore chimico acetilcolina, non noradrenalina.
CONTROLLO ORMONALE DEL SISTEMA CARDIOCIRCOLATORIO
Nella realtà esistono tanti altri meccanismi che controllano la circolazione oltre il controllo
nervoso come quello ormonale. L’adrenalina, ormone secreto dal surrene, vasodilata, al contrario
della noradrenalina che vasocostringe. Sul cuore ha un effetto simile alla noradrenalina, cioè effetto
tachicardizzante, aumento della forza di contrazione, ma in periferia invece di vasocostringere,
vasodilata, soprattutto a livello della muscolatura scheletrica. Tant’è che un aumento di secrezione
della noradrenalina (visto nel I semestre con la regolazione ormonale) non causa un aumento di
pressione arteriosa, anzi paradossalmente può dare un'ipotensione, che aumenta sì la gittata
cardiaca, ma vasodilatando riduce le resistenze vascolari periferiche.
Abbiamo poi nella secrezione ormonale l’angiotensina II, sistema renina-angiotensina (che
abbiamo visto bene con il rene) che entra in gioco in maniera preponderante nel controllo della
tensione a lungo termine, mentre il SN l’adrenalina è un controllo a breve termine, cioè io ho una
vasodilatazione immediata, che protende a essere compensata da altri meccanismi. Oppure si può
avere una forte perdita ematica con scarica simpatica, son tutti eventi che avvengono in acuto.
L’angiotensina II, cioè il sistema renina-angiotensina, è un sistema che tende a controllare la
pressione a lungo termine, cioè ci mette tempo per agire, ma vale per un periodo più lungo. È un
vasocostrittore e ha anche un’azione renale (aumenta l’assorbimento di sodio ecc.). È talmente
importante questo sistema che gran parte dei farmaci antipertensivi oggi giorno vengono usati per
modulare tale sistema sotto vari aspetti, sia inibendo l’ACE, l’enzima di conversione
dell’angiotensina, sia anche inibendo i recettori dell’angiotensina II e prendono anche il nome di
sartani. Qualsiasi medico avrà a che fare con ACE-inibitori e sartani che sono tra i farmaci più
prescritti al mondo. La vasopressina, è un ormone d’emergenza, serve quando abbiamo bisogno di
aumentare la pressione velocemente, è uno dei più potenti vasocostrittori che esistano, però dura
poche ore (ormone ipofisario). Vasodilatatore (come visto prima) soprattutto l’adrenalina, ha
un’azione prevalentemente vasodilatatrice, in alcuni territori vasocostringe, ma l’azione
predominante è l’azione vasodilatatoria.
Altro ormone è il peptide natriuretico atriale che viene secreto dal cuore quando i miociti
vengono stirati, tipo quando vi sono sovraccarichi di volume nel cuore, ed è un’azione a lungo
termine.
CONTROLLO LOCALE DEL SISTEMA CARDIOCIRCOLATORIO
Oltre il controllo nervoso e ormonale esiste un controllo detto locale. In ogni singolo tessuto
ci può essere un controllo locale (lo vedremo meglio alla lezione prossima), ogni singolo tessuto si
autoregola il flusso ematico (come cuore, cervello, muscolo) sulla base delle condizioni locali, per
esempio se ho sul muscolo un forte accumulo di ossigeno, potassio, idrogenioni e anidride
carbonica, io in quel muscolo ho una vasodilatazione. Ad esempio io sono in esercizio, quindi uno
stimolo per il sistema simpatico, in teoria dovrei avere una vasocostrizione però nel muscolo che sta
lavorando uno di questi metaboliti vince sulla stimolazione simpatica e quindi in quel muscolo c’è
una vasodilatazione (in tutto il resto dell’organismo sono vasocostretto).
A livello locale ci sono una marea di sostanze che regolano il flusso sanguigno, sostanze come
quelle viste prima, o di scarto del ciclo di Krebs ma anche per esempio adenosina, sostanza
importantissima soprattutto nel cuore, dove vasodilata le coronarie, ma poi anche metaboliti
dell’acido arachidonico, prostaglandine, prostacicline, bradichinina, chinina e via dicendo. Fra
queste, importante, è il monossido d’azoto (NO). Esso è distribuito indifferentemente in tutti i
tessuti, viene formato dalle cellule dell’endotelio soprattutto, in risposta ad un aumento della
pressione. Aumenta la pressione e anche lo shear stress, cioè lo stress da parete, sulle cellule
appunto endoteliali che si mettono a produrre NO (principale specie chimica ma non l’unica a
essere secreta da tali cellule). Questi sono i vasodilatatori ma esistono anche vasocostrittori, in
particolar modo le endoteline, molecole prodotte dall’endotelio quando c’è una diminuzione di
pressione, ma anche una risposta miogenica. Le cellule muscolari delle pareti dei vasi, in risposta ad
un aumento di pressione, tendono a rispondere a questi aumenti di pressione con un aumento della
loro contrazione. Risposta miogenica non so se avete mai letto nei libri effetto bayliss, dove
praticamente in risposta a uno stiramento queste cellule si contraggono.
Il controllo nervoso è quindi importantissimo nell’immediato, soprattutto per conservare
l’omeostasi cardiocircolatorie in tutte quelle situazioni che cambiano la pressione arteriosa e in
particolar modo in maniera repentina. Il controllo ormonale invece ha possibilità, soprattutto
mediante l’angiotensina II e anche il peptide natriuretico atriale, di modulare a lungo termine la
pressione arteriosa. Il controllo locale infine agisce sul raggio medio arteriolare con una marea di
sostanze che vengono prodotte localmente e variano da tessuto a tessuto.
Questi nell’ultima immagine (che il prof non si aspetta si sappia per l’esame) son tutti i fattori
che controllano la pressione arteriosa (non tutti perché qualcuno manca). Agendo su ognuna di
queste cose si può modulare la pressione arteriosa, a partire dal SN per poi arrivare al controllo
locale, passando anche ad esempio dall’ematocrito. Ci avevate pensato che aumentando ad es il
numero di globuli rossi del sangue aumenta la pressione arteriosa? Avviene in quanto aumenta la
viscosità ematica ed entra in gioco quel fattore dell’equazione di Pouseille.
Il tutto però alla fin fine è basato su quest’equazione finale:
pressione arteriosa media = gittata cardiaca x resistenza vascolare periferica.
Quindi se io so quali sono i fattori che mi modulano la gittata cardiaca e quali sono i fattori
che mi modulano le resistenze vascolari periferiche, posso arrivare a capire cosa in quel momento
mi ha causato la variazione della pressione arteriosa. Questo è importante per il medico quando
arriva un paziente, che per esempio ha la pressione arteriosa alta. Può essere una qualsiasi di queste
cause. Può avere un aumento cronico ad es di adrenalina plasmatica (poco frequente) perché ha un
ferocromocitoma, cioè ha un tumore secernente adrenalina nelle ghiandole surrenali che gli causa
un aumento cronico di frequenza cardiaca e di gittata sistolica, con conseguente aumento della
pressione arteriosa. Oppure posso avere un aumento cronico delle resistenze vascolari periferiche
per aumento cronico dell’angiotensina II, perché ho ad es un tumore renale secernente renina (caso
raro) o perché ad esempio ho una stenosi dell’arteria renale (molto più frequente e possibile). In
pratica arriva poco sangue al rene, le cellule iuxtaglomerulari, in risposta, secernono renina,
attivano il sistema renina-angiotensina che in tutto l’organismo causa un aumento della pressione
arteriosa e si chiama ipertensione reno-vascolare, abbastanza frequente. Quindi tutti questi fattori
presi singolarmente sono in grado di modulare la pressione arteriosa. L’aumento di pressione
arteriosa e l’aumento di ematocrito non è un processo senza logica, c’è una condizione ben precisa
ad esempio nelle leucemie, dove causano un brusco aumento della produzione delle cellule del
sangue (globuli bianchi, globuli rossi), causano quindi un aumento dell’ematocrito e un aumento
della pressione arteriosa.
FISIOLOGIA lez 31/03/2012
Concas Maria Rosaria
Nel controllo globale del circolo, per quanto riguarda il mantenimento della pressione
arteriosa, è fondamentale sia il controllo nervoso sia il controllo ormonale che infatti mantengono
costante la pressione in determinati tessuti per garantire che sia assolta la funzione di quei
particolari tipi di tessuti, che si tratti del rene, del cuore del polmone ecc.; esiste anche la possibilità,
a livello locale, di controllare il circolo locale per soddisfare le esigenze specifiche di ogni singolo
organo.
Dobbiamo tenere ben divisi il controllo generale della circolazione dal controllo locale di
questa, perché ci possono essere dei momenti in cui si ha bisogno di un particolare afflusso di
sangue in determinati distretti, per esempio i muscoli che stanno svolgendo un particolare esercizio,
senza che questo forte afflusso di sangue ai muscoli vada a discapito della circolazione globale
dell’organismo (“..cioè non è che per correre io faccio mancare sangue al cervello..”).
La risposta emodinamica che nasce è il risultato di un “compromesso” atto sia a mantenere
una determinata pressione nell'organismo, sia a garantire un corretto afflusso di sangue nei vari
territori con varie esigenze.
Per esempio il territorio che è risparmiato da qualsiasi tipo di economia è il cervello, un certo
afflusso di sangue in questo organo deve essere sempre mantenuto, dato che mancanza di flusso o
flusso insufficiente può portare ad una ipossia proprio a livello cerebrale con dei danni che possono
diventare irreversibili.
Ora vediamo come è regolata la pressione e la circolazione nei vari distretti, e partiamo
proprio dall'organo responsabile del “movimento” del
sangue in circolo, partiamo proprio dal CUORE.
Vi ricordo brevemente che l'irrorazione arteriosa
del cuore è garantita da due arterie, l'arteria
coronaria sinistra e l'arteria coronaria destra: in
alcuni individui predomina la coronaria sinistra,
quindi buona parte del cuore è irrorata dalla coronaria
sinistra, in altri predomina la coronaria destra, e in
altri casi ancora si ha una situazione intermedia, dove
sia la coronaria destra sia la coronaria sinistra si
spartiscono il cuore. Riferendoci alla coronaria destra
bisogna precisare che questa non irrora solo il cuore
di destra, così come la coronaria sinistra non irrora
solo il cuore di sinistra, le cose sono un po’ più
complicate. La coronaria sinistra ha un grosso ramo, il
ramo circonflesso che è un ramo discendente, l'arteria coronaria destra fa un lungo giro fino a
terminare anche lei con un ramo discendente.
Il sangue venoso ritorna al cuore attraverso due sistemi: uno, tramite le vene che si aprono
nelle camere del cuore, vena cava superiore e inferiore, l'altro con un sistema venoso più
superficiale che sbocca in una formazione che si chiama seno coronarico, per arrivare poi nell'atrio
di destra.
Quali caratteristiche ha il cuore? Intanto a riposo si porta via circa il 5% della gittata cardiaca:
circa il 5% di quello che il cuore pompa, rientra proprio dentro il cuore, nel tessuto cardiaco,
attraverso le arterie coronarie (con un cuore normale, si ha una gittata cardiaca di circa 5L, di cui
50mL rientrano nel cuore). Simile è la percentuale durante sforzo: se io durante sforzo arrivo ad una
gittata cardiaca di 20L, un litro se ne va al cuore; quindi, più o meno, il cuore si “mangia” sempre la
stessa percentuale di quantità di sangue pompato.
Un altro concetto molto importante è la cosiddetta differenza artero-venosa in O2 , che è la
differenza di ossigeno contenuta nel sangue arterioso e in quello venoso e rappresenta l'ossigeno
ceduto ai tessuti; questa nel cuore è elevata anche a riposo. Normalmente la quantità d'ossigeno
disciolto legato all'emoglobina nel sangue arterioso è di circa 200mL per ogni litro di sangue che
arriva. Di questi 200mL il cuore se ne porta via 130mL/L, circa il 70%, ed è tanto considerando che
in qualsiasi tessuto l'estrazione non supera mai l'80% come massimo: il che vuol dire che se io ho
200mL di ossigeno da sfruttare non sono prelevati più 150-160mL, è proprio un limite fisiologico. Il
cuore a riposo, cioè non tachicardico, ne porta già via 130mL (si tenga presente che il muscolo
scheletrico estrae dai 40mL ai 50mL di ossigeno, quindi quello che porta via il muscolo scheletrico
come capacità di estrazione di ossigeno, è portato via nel cuore solo dai ventricoli). Cosa vuol dire
questo? Vuol dire che il cuore può soddisfare aumenti di richiesta di apporto di ossigeno non
aumentando l'estrazione come avviene nella maggior parte dei tessuti, essendo l’ossigeno residuo
molto poco, ma aumentando il flusso di sangue nel circolo coronarico: infatti il flusso coronarico
cresce in modo quasi direttamente proporzionale all’aumento del consumo metabolico di ossigeno
da parte del cuore.
Il cuore è un organo sempre metabolicamente attivo, anche quando siamo a riposo, non è mai
fermo, quindi ha l’esigenza di dover aumentare l'afflusso di sangue in maniera istantanea ogni
qualvolta abbiamo un aumento della richiesta metabolica, ogni qualvolta ci mettiamo in movimento,
ogni qualvolta aumenta la frequenza cardiaca.
C'è poi un’altra cosa da tener presente: mentre
tutti gli altri organi sono irrorati quando il cuore è in
sistole cioè quando la pressione è massima, nel cuore
questo non può avvenire perché quando siamo in
sistole il cuore è contratto soprattutto nel ventricolo
sinistro e la pressione nei tessuti e all'interno della
cavità cardiaca è più alta della pressione dell'aorta.
(da qui dice una serie di frasi che per come le ha
dette sembrano non avere senso, quindi vi dico un po’
cosa ho capito io dalla spiegazione in aula di questi
grafici e da quello che è risultato cercando di mettere
in ordine la sua registrazione)
Come vediamo nel grafico in alto abbiamo la
pressione endoventricolare, cioè la pressione dentro il
ventricolo cardiaco, in questo caso sinistro, e la
pressione in aorta (la linea continua rappresenta la
pressione all'interno del ventricolo; la linea
tratteggiata, la pressione all'interno dell'aorta) e sotto
abbiamo gli eventi corrispondenti del flusso della
coronaria sinistra: quando il cuore si contrae il flusso
della coronaria sinistra scende perché il cuore si sta
contraendo e oppone resistenza perché in quel
momento la pressione nei tessuti è più alta di quella
dell'aorta, il cuore infatti deve vincere la pressione
aortica per poter eiettare; addirittura abbiamo una fase
in cui il flusso diventa negativo che è la fase della
contrazione isovolumica in cui il cuore cerca di vincere la pressione aortica e sviluppa il massimo
della tensione all'interno del ventricolo. Inizia la fase di flusso pian piano, quando la tensione inizia
a scendere e la pressione che c'è all'interno dell'aorta supera quella che c'è all'interno del ventricolo.
La maggior parte del flusso avviene in questo passaggio, quando i ventricoli si rilasciano e la
pressione dell'aorta diventa più alta di quella delle coronarie.
Guyton: Il flusso nei capillari coronarici del miocardio ventricolare sinistro scende ad
un valore molto basso durante la sistole, al contrario di quanto avviene nei capillari di tutti gli altri
distretti vascolari dell’organismo. La ragione di questo fenomeno risiede nella forte compressione
operata dal miocardio ventricolare sinistro sui vasi intramuscolari durante la contrazione sistolica.
Nella coronaria destra il discorso non è lo stesso, infatti mentre il ventricolo di sinistra pompa
contro una pressione media di 100mmHg circa (pressione aortica), il ventricolo di destra pompa
contro il circolo polmonare, e la pressione del circolo polmonare non è 100mmHg, ma è circa
15mmHg, quindi la tensione che deve sviluppare al suo interno il ventricolo di destra è molto
inferiore, sei volte inferiore rispetto a quella che deve sviluppare il ventricolo di sinistra: per cui la
pressione aortica risulta sempre più alta rispetto alla pressione sviluppata nella coronaria destra,
dove quindi ci sarà il flusso anche in sistole. Non assistiamo a quella drammatica diminuzione del
flusso che avviene nella coronaria sinistra quando il ventricolo si contrae perché la tensione
sviluppata è molto più bassa, e questo consente al territorio irrorato dalla coronaria di destra di
essere sempre più ricco di sangue.
Nel seno coronarico le cose sono completamente diverse perché gran parte dell'afflusso di
sangue, cioè il passaggio di sangue venoso, si ha in sistole, “quando il cuore si spreme, spreme
anche le vene, e anche il seno coronarico”, permettendo il ritorno del sangue all’atrio.
Gran parte dei problemi infartuali avvengono a livello del ventricolo sinistro, perché è un
territorio caratterizzato da una tensione dei tessuti sempre molto alta, cosa che può causare una
stenosi della coronaria sinistra che è responsabile soprattutto dell’irrorazione del ventricolo di
sinistra; per questo motivo un deficit provocato da una stenosi della coronaria sinistra porta a dei
risultati molto più gravi, rispetto ad una stenosi della coronaria di destra, anche se comunque la
corrispondenza “coronaria di destra-ventricolo di destra” non è esatta; questa arteria infatti può
andare ad irrorare regioni che appartengono al ventricolo di sinistra in misura che dipende
dall'anatomia dei singoli soggetti.
Nella regolazione del flusso a livello dell'arteria coronaria vediamo coinvolti dei fattori che
non sono generali come abbiamo visto prima, ma sono soprattutto fattori metabolici locali. Il
sistema nervoso simpatico, come abbiamo visto nella lezione precedente, in gran parte aumenta la
pressione nell'organismo per riduzione del calibro delle arteriole ed è anche responsabile
dell'aumento dell'attività sistolica, non funziona nel cuore, qui ha effetti diversi.
Gran parte della regolazione del flusso a livello del cuore è operata dalla presenza di fattori
metabolici locali: ogni qual volta si accumulano dei metaboliti, in particolar modo l'adenosina, si
assiste ad una vasodilatazione; si tratta di un meccanismo di autoregolazione locale, per cui ogni
qual volta c'è un aumento del fabbisogno di ossigeno a livello del cuore, come per esempio un
aumento della frequenza cardiaca, si ha un maggiore consumo di ATP che porta (e questo vale per
qualsiasi tessuto) all'accumulo anche piccolissimo di una certa quantità di adenosina. Ci sono dei
recettori sulla parete del tessuto endoteliale che, in risposta immediata all'accumulo di adenosina,
vasodilatano e aumentano il flusso nel territorio che ha bisogno in quel momento di più sangue, di
più ossigeno. Non troviamo solo l'adenosina, ma comunque è considerato a oggi il fattore più
importante, tant'è che molti farmaci agiscono proprio sui recettori dell'adenosina per controllare la
vasodilatazione.
Un altro elemento è il lavoro cardiaco che andrà comunque ad agire sull'adenosina: se io
aumento il lavoro cardiaco, per aumento della frequenza cardiaca, ho un accumulo di adenosina per
cui aumento la vasodilatazione.
Abbiamo poi una serie di meccanismi endoteliali e ormonali locali di cui il più importante è
l'NO, è un altro vasodilatatore e viene sintetizzato dalle cellule endoteliali in risposta a diversi
stimoli come l'aumento del lavoro cardiaco, o soprattutto in risposta ad un aumento dello stress
della parete del vaso: se questo stress al passare del sangue supera determinate soglie allora viene
prodotto l'NO che tende a vasodilatare, e a ridurre così lo stress a livello della parete. Sono
meccanismi di autoregolazione locale che esistono in molti tessuti, praticamente quasi tutti, e il
cuore ha la capacità di sfruttarli in maniera ottimale, infatti molti farmaci che vengono dati per
vasodilatare le coronarie, per esempio il Carvasin, pastigliette che usano gli anginosi da mettere
sotto la lingua, agiscono aumentando l'NO e quindi inducendo la vasodilatazione delle coronarie.
Altri fattori sono per esempio le endoteline (che però hanno l'effetto opposto), derivati
dell'acido arachidonico, prostaglandine etc. che mediano degli effetti molto complessi, intricati fra
di loro.
L’aumento della pressione aortica inoltre contribuisce ad aumentare l'afflusso a livello delle
coronarie.
Parlando di meccanismi nervosi sappiamo che una scarica ortosimpatica in linea di massima
causa vasocostrizione, ma nel cuore non è così. I recettori per la noradrenalina, i recettori
adrenergici, si distinguono in due sottoclassi, alfa e beta: quelli alfa, tendono a vasocostringere,
quelli beta riguardano di più gli effetti metabolici; nel cuore predomina l'effetto beta, cioè l'effetto
metabolico, cioè predomina, se io ho una scarica simpatica l'effetto sulla frequenza cardiaca, e un
aumento della frequenza cardiaca, rappresenta un aumento del metabolismo cardiaco, quindi
aumenta l'accumulo di adenosina e si ha vasodilatazione. Non ho un effetto diretto
dell'ortosimpatico sul vaso, ma assisto a un effetto mediato dall'aumento del metabolismo; per cui
l'ortosimpatico in linea di massima, sia l'adrenalina che rappresenta l'ortosimpatico ormonale, sia la
noradrenalina che rappresenta l'ortosimpatico nervoso, causano una vasodilatazione mediata
dall'accumulo di fattori metabolici locali. Il parasimpatico non fa praticamente niente, se non
indirettamente rallentando le frequenza cardiaca, rallentando quindi il metabolismo cardiaco e
causando vasocostrizione, riducendo il flusso di sangue. Sono comunque degli effetti sul
metabolismo e non sui vasi direttamente.
Abbiamo anche dei fattori ormonali:
• l'adrenalina, che abbiamo già visto
• la tiroxina, è un ormone tiroideo, quindi aumenta il metabolismo del cuore causando
vasodilatazione
• il glucagone è simile alle ammine simpatiche, cioè all'adrenalina e alla noradrenalina, anche
come struttura, fa contrarre il cuore con maggiore forza ( Il glucagone provoca un aumento
dell’AMPc intracellulare a cui consegue un aumento della disponibilità intracellulare di calcio)
determinando un aumento del metabolismo cardiaco e una conseguente vasodilatazione, comunque
indiretta
• La vasopressina nel cuore ha due effetti ed è una delle poche sostanze che riesce a
vasocostringere anche a livello del cuore; per valutare il suo effetto dobbiamo tener conto del fatto
che se la vasocostrizione, indotta dalla vasopressina, aumenta, aumenta anche il lavoro cardiaco
perché il cuore deve pompare di più contro un aumento della pressione arteriosa che è
accompagnato da un aumento della resistenza al flusso, quindi si ha un aumento del suo
metabolismo e si accumula adenosina.
Da tener presente comunque che la vasopressina vasocostringe anche le arterie coronarie per
cui, spesso, non si ha nessuna variazione di flusso e le due cose si compensano.
In tutti i tessuti esiste una vasocostrizione miogena, in risposta ad aumenti della pressione, per
cui ad aumenti dello stress di parete, dello stress che deve sopportare l'endotelio, i vasi normalmente
rispondono tendendo a vasocostringere che è il contrario del meccanismo sopra spiegato. Ho due
meccanismi, e nonostante uno tenda a vasodilatare, all'aumento dello stress di parete c'è anche un
tentativo di vasocostringere, un tentativo di opporsi al flusso; anche nel cuore esiste una possibilità
di vasocostrizione miogena, ma in linea generale vince il meccanismo della vasodilatazione.
Ci sono degli esperimenti in cui io noto che, prendendo un’arteria coronaria se lascio integro
l'endotelio e lascio aumentare il flusso e la pressione, vince la vasodilatazione provocata dall’NO;
se con vari sistemi si toglie l'endotelio vince la vasocostrizione.
Passiamo al CERVELLO. Esso viene protetto in maniera assoluta da qualsiasi tipo di calo
della pressione (l'afflusso di sangue al cervello viene garantito anche fino a pressioni di 4050mmHg), infatti se una persona ha un deficit di irrorazione al cervello deve proprio avere un
profondo calo pressorio, fenomeno identificato come sincope: o gli si blocca il cuore, oppure ha una
vasodilatazione pazzesca per cui le resistenze vengono meno con conseguente mancanza di flusso
di al cervello.
Il cervello ha un elevato consumo di ossigeno, il 13% della gittata cardiaca a riposo (siamo
intorno ai 300-400mL di sangue al minuto), vanno al cervello.
Il sistema nervoso autonomo ha scarsi effetti, se io ho un’efficace scarica simpatica, non
vasocostringo e il parasimpatico ha effetti ancora più scarsi. Ancora di più rispetto al cuore, gran
parte della regolazione del circolo cerebrale è metabolica ed è possibile attuare questa regolazione
del flusso sulla base dell'attività cerebrale in quel determinato momento: se io sto usando il centro
del linguaggio, ci sarà una grande vasodilatazione nel centro del linguaggio, se io sto usando la
corteccia motoria per muovere le gambe ci sarà vasodilatazione in quella parte di corteccia.
Il fattore metabolico più importante è rappresentato dall'accumulo di CO2, la quantità di
anidride carbonica prodotta dal tessuto in quel momento, che è proporzionale all'attività cerebrale
(infatti il cervello per lavorare consuma in gran parte glucosio, che entra in ciclo di Krebs di cui uno
dei prodotti finali è proprio la CO2); quindi la quantità di CO2 che io sto producendo è proporzionale
all'impegno metabolico, più CO2 produco più quel tessuto sta consumando glucosio e quindi sta
lavorando. L’anidride carbonica immessa poi nel torrente circolatorio determina un'ipercapnia
locale; anche l'ipercapnia generale causa un'intensa vasodilatazione: se smettiamo di respirare per
uno e due minuti, questo causa un aumento di CO2 in tutto l'organismo, compreso il cervello, e se
aveste la possibilità di misurare il flusso ematico cerebrale, con un doppler alle carotidi per
esempio, vi accorgereste che c'è un intensa vasodilatazione indotta appena dopo 10-15sec.
Anche la mancanza di ossigeno, l’ipossia, determina vasodilatazione, anche se è comunque
molto meno potente rispetto all'ipercapnia: se io vado in montagna, diminuisce la percentuale di
ossigeno nell'aria, e quindi la pressione parziale di ossigeno che respiro, avrò così una
vasodilatazione dovuta all'ipossia, che è comunque molto inferiore rispetto a quella determinata
dall'ipercapnia.
Esistono altri metaboliti implicati, non solo la CO2, quali il potassio, l'adenosina, l'accumulo
di idrogenioni, però comunque nessuno di questi è potente come l'ipercapnia che peraltro non si sa
bene nemmeno come funzioni: secondo alcuni funziona in maniera diretta, cioè l'aumento della
concentrazione di anidride carbonica nel sangue produce un aumento, per il legame con l'acqua, di
acido carbonico, a cui consegue l'accumulo di idrogenioni che vasodilata.
Abbiamo come ultima possibilità un'autoregolazione miogena simile a quella che abbiamo
visto nel cuore, e ad aumenti pressori si tende a ridurre il calibro dei vasi.
Ora vediamo come viene regolato il circolo a livello del MUSCOLO SCHELETRICO.
La muscolatura scheletrica rappresenta, in una persona di normali dimensioni, circa il 40-50%
del perso del corpo, e normalmente la muscolatura, quando si mette in attività, consuma ossigeno in
maniera importante, per cui buona parte della capacità di pompa del nostro cuore è dettata dalle
esigenze dei nostri muscoli; se noi non ci muovessimo mai e avessimo una gittata cardiaca di 5 L
non ci sarebbe nessun bisogno di aumentare la frequenza cardiaca, se noi fossimo sempre fermi il
nostro apparato cardiocircolatorio con i suoi 60-70 battiti al minuto andrebbe benissimo e non ci
sarebbe nessun bisogno di variarne il ritmo. Tutte le variazioni che avvengono in aumento di gittata
cardiaca avvengono per soddisfare le esigenze muscolari: quando durante un esercizio si
raggiungono i 20-25L di gittata cardiaca, quei litri in più rispetto a una condizione di riposo vanno
ai muscoli, e quindi l'esigenza di incrementare la gittata cardiaca è un'esigenza dettata dal bisogno
di soddisfare le richieste muscolari a fronte di un lavoro, di un movimento. A riposo, ai muscoli è
destinato dal 10% al 20% della gittata cardiaca, e da questa quantità di sangue che gli arriva,
estraggono pochissimo ossigeno, 40-50mL per ogni litro di sangue; in esercizio le cose cambiano in
maniera drammatica perché circa l'80% della gittata cardiaca va a finire nei muscoli (se io sotto
sforzo ho una gittata cardiaca di 20L vuol dire che di quei 20L, 16L se ne vanno ai muscoli che in
quel momento lavorano). Non solo arriva molto più sangue, ma da questo sangue i muscoli non
estraggono più solo 40-50mL per litro, ma ne estraggono 150-160mL, quindi aumenta il flusso e
aumenta anche l'estrazione di ossigeno nel tessuto che sta lavorando. C'è un’esigenza pressante ogni
qual volta i muscoli si mettono in movimento di garantirgli un flusso di sangue adeguato. A riposo
domina il tono simpatico costrittore e l’autoregolazione miogena, cioè quando io sono fermo, se mi
arriva una scarica simpatica c'è una forte vasocostrizione muscolare, non solo nei muscoli ma anche
nei muscoli: infatti quando parlo di vasocostrizione da ortosimpatico a riposo abbiamo una
vasocostrizione anche nei visceri, nel rene, e nella cute, tutti siti “prediletti” dall'ortosimpatico
quando induce una vasocostrizione a riposo. Se vado in esercizio non funziona così: non in tutti i
muscoli ma solo nel muscolo che sta lavorando, anche se ho una forte scarica simpatica (e durante
l'esercizio questo avviene), predomina la regolazione metabolica e la vasodilatazione. Durante
l'esercizio aumenta la scarica simpatica, che induce vasocostrizione dappertutto meno che nel
cervello, nel cuore e nel muscolo che sta lavorando, in questi predomina una regolazione metabolica
basata sull'accumulo di sostanze come la CO2, che come nel cervello è un indicatore di attività del
muscolo stesso, l'acido lattico, anche lui indicatore di attività, idrogenioni, adenosina, metaboliti
dell'acido arachidonico, prostaglandine, chinine ecc.. e abbiamo un'inibizione del tono simpatico a
livello locale. Mentre in generale il sistema simpatico è vasocostrittore, l'adrenalina secreta dalle
ghiandole surrenali in questa sede ha un effetto vasodilatatore che predomina, mentre la
noradrenalina vasocostringe negli altri muscoli l'adrenalina vasodilata. L'effetto totale che si ha nel
muscolo dipende per prima cosa dall’attività del muscolo, poi dal tono del simpatico, e ancora dalla
quantità di adrenalina.
Abbiamo un'autoregolazione miogena anche in questi tessuti ma domina anche qui la
regolazione metabolica.
Quando vado in
esercizio i muscoli che
non stanno lavorando,
hanno una forte
vasocostrizione, ma nel
muscolo che invece sta
lavorando, un po’ come
nel cuore, va considerato il
fatto che con la
contrazione oppone
resistenza al flusso, per cui
se io considero un lavoro
ritmico caratterizzato da
contrazione e
rilasciamento, l'aumento
del flusso avverrà in
rilasciamento; se io invece
faccio un lavoro
isometrico, dove il
muscolo si contrae ma non
si accorcia, non ci sarà
un’attività ritmica del flusso ematico ma questo sarà pari a zero o addirittura andrà sotto lo zero, un
po’ come quando il cuore va in contrazione isovolumica. Questo è uno dei motivi per i quali la
contrazione isometrica non può essere mantenuta a lungo, se io cerco di sollevare un peso più forte
rispetto alla capacità del mio muscolo, e il muscolo rimane contratto per più di qualche decina di
secondi dopo un po’ deve mollare per la fatica, e uno dei motivi della fatica è che non arriva più
sangue, perché la contrazione muscolare opprime i vasi e la tensione sviluppata dai muscoli, è
superiore alla pressione arteriosa. La contrazione isometrica non è un tipo di attività che noi
facciamo di continuo, la maggior parte delle attività come camminare o pedalare, prevedono
un’attività ritmica di contrazione e rilasciamento.
Poi abbiamo il CIRCOLO RENALE che abbiamo in parte già visto. Esso a riposo riceve
buona parte della gittata cardiaca per motivi, più che metabolici, dovuti alla filtrazione. Il flusso
renale possiede un'autoregolazione miogena, quindi all'aumento della pressione l'arteria tende a
chiudersi per evitare troppi sbalzi pressori nelle pareti renali. L'ortosimpatico può causare intense
vasocostrizioni e inibire l'autoregolazione. Di fronte ad una forte scarica simpatica anche il rene
tende a vasocostringere e questo ha un'importanza soprattutto clinica: uno dei problemi più grossi
quando si va in shock (= forte abbassamento della pressione arteriosa per periodi prolungati di
tempo), per esempio in caso di forte emorragia, è che la scarica simpatica che tenta di mantenere
costante la pressione induce una vasocostrizione renale troppo intensa che alla lunga porta da
un’ischemia renale e alla morte del rene, cioè all'insufficienza renale. Uno dei problemi che infatti
si trova ad affrontare un rianimatore che deve soccorrere chi è in stato di shock di tipo cardiogeno o
ipovolemico è quella di preservare il rene; quindi l'ortosimpatico gioca un brutto scherzo sul rene
perché una vasocostrizione molto intensa può portare alla morte ischemica renale.
Ci sono una serie di sostanze umorali in grado di vasodilatare i vasi renali tra cui
prostaglandine, bradichinina ecc.. che vengono prodotte lo stesso con un meccanismo di
autoregolazione; ((anche qui cercato di decifrare)) sono importanti le prostaglandine, fattori renali
che vengono inibiti dagli antinfiammatori, per cui una cura antinfiammatoria deve essere prescritta
con una attenzione particolare ai dosaggi proprio per non inibire questi fattori di regolazione renale.
Guyton: Gli ormoni e le sostanze autacoidi che causano vasodilatazione e aumentano
il flusso ematico renale e la VFG includono le prostaglandine e la bradichinina. Anche se queste
sostanze vasodilatatrici non sembrano avere grande importanza nel regolare il flusso ematico renale
o la VFG in condizioni normali, esse potrebbero limitare gli effetti vasocostrittori renali dei nervi
ortosimpatici o dell’angiotensinaII, in modo particolare gli effetti costrittori sulle arteriole afferenti.
Opponendosi alla vasocostrizione delle arteriole afferenti, le prostaglandine potrebbero aiutare a
prevenire una eccessiva riduzione della VFG e del flusso ematico renale. In condizioni di stress,
come per riduzione del volume ematico o dopo un intervento chirurgico, la somministrazione di
sostanze antinfiammatorie non steroidee, come l’acido acetilsalicilico, che inibisce la sintesi di
prostaglandine, potrebbe causare una significativa riduzione della VFG.
Il rene ha dei meccanismi di
autoregolazione che funzionano con
variazioni di pressione molto ampie,
possiamo passare da 50-60mmHg
fino a 160-170mmHg. Le variazioni
di pressione non determinano quindi
una variazione del flusso a livello
glomerulare perché il rene si
autoregola con questi meccanismi:
vasocostrizione miogena, aumento
della sintesi di prostaglandine e il
cosiddetto feedback tubulo
glomerulare. Il rene quindi autoregola
il flusso entro questi ambiti di
pressione automaticamente.
CIRCOLO CUTANEO
Perché è importante il circolo cutaneo? E’ importante soprattutto per la termo-dispersione, per
eliminare cioè calore quando abbiamo bisogno di rimuoverlo.
A riposo abbiamo circa il 5-6% della gittata cardiaca che arriva a questo tessuto, che non
viene sfruttata a fini metabolici ma solamente per termo-disperdere. Durante l'esposizione al calore
il flusso cutaneo può aumentare fino a dieci volte, circa il 60% della gittata cardiaca può
raggiungere la cute per la dispersione del calore. Questo è possibile a livello della cute per l’effetto
di un’intensa vasodilatazione regolata soprattutto a livello nervoso e per l'apertura di circoli
collaterali: in determinati distretti, tra cui la cute, esiste tutta una serie di vasi collaterali di cui la
maggior parte sono chiusi e che si
aprono sulla base delle esigenze
dell'organismo; normalmente sono
chiusi, poi se in una giornata
particolarmente calda ho bisogno di
termo-disperdere, questi circoli si
aprono, facendo arrivare una marea di
sangue nella cute che disperde il calore
del corpo attraverso il sangue stesso e
attraverso la vasodilatazione.
Al contrario l'esposizione al
freddo può quasi annullare il circolo
cutaneo, e possiamo andare incontro a
una vasocostrizione tale da avere
necrosi in alcune parti del tessuto: i
cosiddetti geloni sono il risultato di un
intensa vasocostrizione cutanea in
risposta al freddo che causa necrosi di
una parte del tessuto cutaneo; non sono
altro che tessuto morto e fanno male
proprio nella fase in cui ritorna il calore
e ritorna il flusso che causa
infiammazione per la presenza di tessuto
necrotico che deve essere rimosso. Il
circolo cutaneo è molto importante
perché serve a regolare la temperatura,
diminuendo o aumentando l'afflusso di
sangue a livello della cute in base alla
temperatura corporea.
Bisogna parlare anche della
CIRCOLAZIONE FETALE, che è importante anch'essa: la placenta è il polmone del feto, dove
avvengono gli scambi di gas e sostanze nutritive tra la mamma e la circolazione del feto. Il sangue
arriva alla placenta attraverso due arterie che si chiamano arterie ombelicali (notiamo nella slide la
parte discendente dell'aorta, la biforcazione dell'aorta e poi le due arterie ombelicali), che dopo la
nascita andranno ad obliterarsi e diventeranno tessuto fibroso; da queste arterie arriva il sangue alla
placenta ed esso ritorna al cuore attraverso la vena ombelicale che tramite il dotto venoso confluisce
nella vena cava inferiore che poi porterà il sangue al cuore. Questo sangue, pur essendo sangue
venoso è ricco di ossigeno, non ha nessun bisogno di andare ai polmoni ma può essere
tranquillamente distribuito al corpo senza passare nel circolo polmonare. Il sangue arriva all'atrio
destro, passa poi nel ventricolo di destra e viene pompato nell'arteria polmonare e da qui in gran
parte viene deviato nell'aorta attraverso il dotto arterioso di Botallo che mette in comunicazione
l'aorta con l'arteria polmonare, il sangue passa quindi dalla circolazione destra a quella sinistra. Non
accade solo questo: il sangue che è entrato nell'atrio di destra, sangue venoso che si chiama così
perché torna al cuore ma è ricco di ossigeno e quindi ha una composizione tipica del sangue
arterioso, dall'atrio di destra va in gran parte nell'atrio di sinistra attraverso il forame ovale, da qui
passa al ventricolo di sinistra e poi viene eiettato in aorta.
A livello del dotto di Botallo, che ha la parete formata da fibre circolari lisce, ho una forte
vasocostrizione, e questo come nasciamo per effetto di tutta una serie di fenomeni si chiude così
come le formazioni che caratterizzano la circolazione fetale; uno di questi fenomeni è il tentativo
del neonato di aprire la gabbia toracica anche col primo respiro, e il dotto venoso si chiude per
effetto della contrazione muscolare addominale; si chiude anche il forame ovale, grazie al
meccanismo a valvola che lo caratterizza, grazie all’aumento della pressione della parte sinistra del
cuore rispetto a quella di destra. Il sangue è così obbligato a passare nel ventricolo di destra e
successivamente ai polmoni tramite il circolo polmonare.
Tutte queste formazioni possono persistere dopo la nascita, alcune si chiudono bene, altre non
si chiudono, come per esempio il forame ovale che darà così luogo a una cardiopatia congenita,
anche se benigna, stessa cosa per il dotto arterioso che può rimanere pervio determinando una
situazione in cui è necessario intervenire chirurgicamente per evitare che sangue venoso poco
ossigenato passi a sinistra dove invece il sangue è molto ossigenato. Il passaggio attraverso il foro
ovale nella vita fetale è consentito perchè la pressione nell'atrio di destra è maggiore di quella
dell'atrio di sinistra, quando nasciamo cambia tutto, la pressione dell'atrio di sinistra supera quella
dell'atrio di destra e la valvola si chiude.
Il CIRCOLO LINFATICO è un circolo parallelo a quello sanguigno che serve a portar via
tutto ciò che filtra dai capillari nell'interstizio e non dovrebbe filtrare, in particolar modo liquido
interstiziale e proteine che fuoriescono infatti dal capillare senza però rientrarvi; normalmente le
proteine non dovrebbero passare dai capillari, ma siccome nessuno è perfetto qualcosa passa sempre
e non va bene, dato che prima di tutto richiamano liquidi a livello dell’interstizio. Una delle
condizioni dovute a difetti del sistema linfatico è la cosiddetta elefantiasi, caratterizzata soprattutto
da rigonfiamento degli arti inferiori.
La funzione principale del sistema linfatico è quindi principalmente portar via tutti i liquidi e
tutte le proteine che sono filtrati al di fuori dei capillari , per poi riportarli nel sistema circolatorio.
Altra cosa da considerare è l'aspetto immunologico, dato che nei vasi linfatici è pieno di linfociti, e i
linfonodi si trovano proprio lungo il decorso dei vasi linfatici.
I vasi linfatici sono costituiti da cellule endoteliali disposte con delle particolari fenestrature
che ricordano un po’ le nasse della pesca, ciò che entra dentro non può più uscire perché trovano la
strada sbarrata da questo meccanismo a valvola, il materiale riesce a entrare ma se preme per uscire
la fenestratura si chiude e non esce più.
Guyton: L’esigua quantità di liquido che ritorna in circolo attraverso le vie linfatiche
è estremamente importante perché le sostanze ad alto peso molecolare, come le proteine, non
possono essere riassorbite in alcun altro modo, mentre possono penetrare quasi senza impedimenti
nei capillari linfatici. Ciò è dovuto alla particolare struttura dei capillari linfatici. Le cellule
endoteliali del capillare sono fissate con dei “filamenti di ancoraggio” al connettivo circostante. Alle
giunzioni delle cellule endoteliali adiacenti, il bordo di una cellula endoteliale si sovrappone al
bordo della cellula vicina in modo che il margine sovrapposto può fluttuare liberamente verso
l’interno del capillare; si forma così una minuscola valvola che tende ad aprirsi verso il lume del
capillare. Il liquido interstiziale, insieme alle particelle in esso sospese, spinge la valvola ad aprirsi
ed entra direttamente nel capillare linfatico; una volta entrato, però, il liquido può uscire solo con
difficoltà, perché qualsiasi reflusso tende a chiudere i lembi valvolari.
Nel capillare linfatico sono presenti delle valvole strutturate come quelle delle vene che
consentono un flusso unidirezionale, il liquido non può tornare indietro ma solamente andare avanti.
Questa unidirezionalità è garantita anche da un rivestimento esterno al capillare linfatico di cellule
di tessuto muscolare liscio che ciclicamente danno delle contrazioni, e premendo il flusso va avanti,
obbligato a salire verso vene. La mancanza di capillari linfatici o il loro malfunzionamento
determina una malattia che si chiama elefantiasi, caratterizzata da delle ipertrofie e delle iperplasie
dei tessuti enormi. Quindi la mancanza di strutture così semplici e banali come possono essere le
strutture linfatiche porta a delle serie conseguenze, e non c'è nemmeno granché da fare.
Mauro Badas
03-04-2012 –
Fisiologia Crisafulli
AGGIUSTAMENTI CARDIOCIRCOLATORI DURANTE L'ESERCIZIO FISICO
Con questa lezione concludiamo l'apparato circolatorio. Parliamo di aggiustamenti ma anche
di adattamenti cardiocircolatori durante l'esercizio fisico.
Il sistema più semplice per mettere sotto stress l'apparato cardiocircolatorio e vedere se questo
funziona bene, è far fare al paziente dell'esercizio fisico. Se per esempio una persona ha difficoltà a
fare le scale, ha il fiatone, significa che c'è qualcosa che non va e la causa spesso è un
malfunzionamento dell'apparato cardiocircolatorio. La forma più fisiologica di stress che deve
sopportare l'organismo umano è proprio l'esercizio fisico.
Se noi non avessimo necessità di muoverci, di fare esercizio fisico, andrebbero benissimo i
parametri che si hanno a riposo (GC= 5 L/min). Di fatto invece abbiamo bisogno di una apparato
cardiocircolatorio che in determinate condizioni necessità di aumentare questo valore per arrivare a
valori di 20-25 L/min (in alcuni casi 30-35 L/min); tutto ciò in un cuore normale viene fatto
facilmente, in un cuore patologico, invece, viene fatto con molta difficoltà: Il sintomo cardine
dell'insufficienza cardiaca è la fatica.
Vediamo quindi cosa succede in un cuore normale durante l'esercizio fisico sia in termini di
aggiustamenti che di adattamenti cardiocircolatori.
Aggiustamenti: sono quelli che avvengono in acuto; ovvero io sono fermo, mi metto in
movimento e ciò determina una aumento della frequenza cardiaca (FC), della gittata sistolica (GS)
ecc.
Adattamenti: sono qualcosa di cronico, ovvero ciò che succede in un organo (in questo caso il
cuore) quando io lo espongo ad un determinato stress, in questo caso l'esercizio fisico. Gli
adattamenti sono quindi ciò che viene determinato con il tempo. Questi hanno la funzione di
rendere più efficace il funzionamento del cuore.
In questa immagine vediamo la
distribuzione della gittata cardiaca in
due condizioni differenti, a sinistra
nella condizione di riposo, mentre a
destra durante l'esercizio strenuo.
A riposo:
PC 5 L/min
di questi:
4-5% cuore
25-30% visceri addominali:
fegato, intestino, stomaco...
20-25% reni
3-5% ossa
15% encefalo
5% cute (anche se questa
percentuale varia molto a seconda delle
condizioni esterne, nel caso si debba
liberare calore)
5-20% muscolo
In esercizio strenuo, ovvero
quando la portata cardiaca raggiunge Illustrazione 1: Distribuzione della portata cardiaca a
valori di circa 25 litri al minuto, la
riposo e durante esercizio strenuo
distribuzione varia notevolmente.
L'80-85% va al muscolo, il quale in condizioni di esercizio fisico richiede notevoli quantità di
sangue, e quindi anche di ossigeno. Una certa quota continua ad andare al cuore, e questa
percentuale rimale stabile al 4-5%, mentre ciò che consegue nell'esercizio strenuo è una
diminuzione di tutte le altre quote; per esempio per quanto riguarda il sangue che giunge al rene, dal
20-25% che si ha in condizioni di riposo si giunge al 2-3%. Tutto ciò avviene a causa degli
aggiustamenti che si hanno in acuto: c'è un'intensa vasodilatazione a livello dei muscoli che si
contraggono; questi sono sganciati dal controllo del Simpatico in quanto vince la vasodilatazione
metabolica locale, per cui anche se ho una grossa scarica simpatica che vasocostringe in tutto il
resto dell'organismo, nel muscolo che sta lavorando prevale la vasodilatazione metabolica.
In questa immagine vediamo la distribuzione della gittata cardiaca a riposo e durante
l'esercizio. La quota che va alla cute dipende molto dalla
necessità che ho di liberare calore. Ciò significa che se sto
facendo esercizio in un ambiente freddo questa quota può
essere molto ridotta; se l'esercizio invece viene fatto in un
ambiente caldo una quota più alta di sangue è destinata alla
cute per esigenze di termoregolazione.
Perché è necessario che nei muscoli giunga ossigeno?
Questo serve per risintetizzare in maniera molto rapida l'ATP consumato dalle contrazioni
muscolari. Questo può essere fatto in maniera molto parziale dai metabolismi anaerobici, nel
senso che questi sono in grado di tamponare il consumo di ATP per brevi periodi di tempo (20-30
sec.); dopodiché, se non interviene il metabolismo aerobico, tutto si ferma in quanto il
metabolismo anaerobico (che sia lattacido o alattacido) ha come conseguenza l'accumulo di
metaboliti come fosfato e idrogenioni che determinano fatica. In seguito alla presenza di questi
metaboliti, quindi, giunge a livello del SNC la sensazione di fatica che impone al muscolo di
fermarsi prima che questo accumulo tenda a causare danni. L'accumulo di idrogenioni porta infatti a
delle conseguenze gravi per cui è necessario mettere subito in moto il metabolismo aerobico che
produce come metaboliti solo la CO2 che comunque può essere facilmente eliminata respirando.
• ADP+Pi+PCr ↔ ATP+Cr+Pi
(metabolismo anaerobico alattacido)
• Glucosio+2ADP+2Pi ↔ 2Lattato+2H+ +2ATP
(metabolismo anaerobico lattacido)
• Glucosio+6O2+34ADP+34Pi ↔ 6CO2 +34H2O +34ATP
(metabolismo aerobico)
Per sostenere il metabolismo aerobico, in condizioni di esercizio fisico, i muscoli possono
aumentare il loro fabbisogno di ossigeno di oltre 50 volte rispetto alle condizioni di riposo. Questo
aumento di fabbisogno di ossigeno deve essere sostenuto necessariamente da un aumento della
portata cardiaca che in condizioni di riposo abbiamo detto essere di circa 5-7 L/min, mentre sotto
sforzo si arriva fino a circa 30-35 L/min (in letteratura sono stati riportati valori di anche 40 L/min
relativi ad atleti di livello molto elevato). Valori fino a 30 L sono abbastanza frequenti negli atleti di
livello medio; le persone sedentarie difficilmente riescono a raggiungere questi valori fermandosi ad
una PC di circa 15-16 L/min. Dalla condizione di riposo all'esercizio, la maggiore richiesta di
ossigeno, viene garantita quindi dalla possibilità di quintuplicare la portata cardiaca.
Questo meccanismo di aggiustamento
rapido è molto preciso. Se io in un grafico
metto in ascissa il consumo di ossigeno, e in
ordinata la portata o gittata cardiaca, vediamo
che ciò che ne risulta è un grafico con un
andamento quasi rettilineo. Ci sono dei sistemi
di controllo nell'organismo che fanno quindi in
modo che la PC si aggiusti in maniera perfetta
rispetto alle esigenze metaboliche (consumo di
ossigeno) dell'organismo.
Questo è proprio il grafico reale di un
atleta che si è sottoposto a un test da sforzo al
cicloergometro, contro carichi via via
crescenti, e in questo caso ad ogni
minuto il cicloergometro incrementava
automaticamente il carico e quindi la
resistenza dei pedali della bicicletta di
circa 50W; nello stesso tempo durante il
test viene misurato il consumo di O2
(V02) e la portata cardiaca. Il consumo
di ossigeno viene rilevato con il
metabolimetro mentre la portata cardiaca
viene misurata in questo caso con la
calorimetria indiretta. Questo è un atleta
“ottimo”, 5 litri al minuto è un valore
molto alto (tenete presente che in
condizioni di riposo abbiamo un valore
di consumo di ossigeno che va dai 250-300 mL/min); vedete nel grafico che ad ogni aumento di
consumo di O2 corrisponde un aumento lineare della PC. Questa regolazione è data in gran
parte dal sistema nervoso autonomo.
In quest'altra immagine vediamo la stessa cosa riferita però ad un gruppo di atleti, con la
differenza che il consumo di O2 è in ordinata mentre la portata cardiaca si trova in ascissa (riportata
come CO= cardiac output → gittata cardiaca). Si può vedere che con tutti i valori ottenuti
dall'insieme di atleti si può descrivere una retta di regressione lineare e posso anche calcolare
quanti litri di portata cardiaca mi servono per ogni litro di incremento di O2 consumato; in questo
caso vedete che ogni qualvolta incremento di 1L il consumo di O2 la gittata cardiaca deve
aumentare di circa 5-6 L(nel grafico riportato 5,95 L). Per questo motivo se io conosco il consumo
di ossigeno di un soggetto, indirettamente posso quindi risalire a quella che è la sua portata
cardiaca: se ha per esempio 3 litri di consumo di O2 la sua PC sarà di circa 15L/min.
Riassumendo, quindi, la relazione tra portata cardiaca e consumo di ossigeno è molto preciso
e possiamo rappresentarlo con una linea retta.
Come è possibile
aumentare la portata
cardiaca PC? Abbiamo a
disposizione due strategie: o
aumenta la frequenza
cardiaca FC (in inglese HR
→ heart rate) o la gittata
sistolica GS(in inglese SV).
Come vedete nei
grafici questo gruppo di
soggetti ha aumentato la
portata cardiaca e vedete
cosa è successo alla gittata
sistolica e alla frequenza
cardiaca, cioè sono
aumentate sia l'una che
l'altra: il risultato è un
incremento della PC.
Quindi, perché la
portata cardiaca aumenti,
è necessario che aumenti:
1)
la frequenza cardiaca;
2)
la gittata sistolica;
3)
entrambe → ovvero ciò che succede in condizioni di salute;
Infatti, uno dei problemi dei malati di cuore è proprio l'impossibilità di aumentare la GS in
risposta all'esercizio, anzi in risposta all'esercizio può succedere anche che si abbassi. L'unica
strategia che hanno gli scompensati cardiaci per aumentare la PC consiste nell'aumentare la FC:
infatti queste persone sono tachicardiche, basta un minimo sforzo per far passare la FC da 80-90
bpm a riposo a 150 bpm.
Di quanto può aumentare la frequenza cardiaca? Che riserva di frequenza cardiaca
abbiamo? Un organismo normale passa da valori di FC 1di 60-70 a riposo fino a 200-210; dipende
dall'età del soggetto, infatti esiste una regola empirica2 molto approssimativa che dice che la
1 A riposo la frequenza cardiaca di un essere umano adulto è di circa 70 bpm nell'uomo e di circa 75 bpm nelle donne,
mentre nei neonati la frequenza a riposo è di circa 100-180 bpm.
ETA' – FREQUENZA CARDIACA NORMALE (NEONATI 100-180; BAMBINO 70-110; ADOLESCENTI 70-90;
ADULTI 65-85) – fonte: http://it.wikipedia.org/wiki/Frequenza_cardiaca
2 Formula di Karvonen
frequenza cardiaca massima raggiungibile da un soggetto è data dalla formula:
HR max = 220 – età
e quindi per esempio una persona di 60 anni potrà avere una HR max di 160 bpm; è un
calcolo che ha circa il 20% di errore. Le persone giovani durante l'esercizio fisico possono arrivare
a triplicare i valori di frequenza cardiaca rispetto ai valori di riposo e quindi:
supponiamo di mantenere costante il valore di GS, aumento di 3 volte la FC di conseguenza la
portata cardiaca aumenta di 3 volte (cioè se la PC a
riposo è di 5L/min, triplicando la frequenza
cardiaca arrivo a 15L/min). La portata cardiaca
arriva però anche a valori di 30 L/min, quindi è
necessario che aumenti anche la gittata sistolica
(GS).
Che differenza c'è tra una persona allenata e una
sedentaria nella capacità di aumentare la
frequenza cardiaca? La differenza è quella che si
vede nel grafico. A parità di età consideriamo un
individuo allenato e un individuo sedentario. La
frequenza cardiaca massima alla quale possono
arrivare è la stessa (200bpm); quello che cambia è il
rapporto tra frequenza cardiaca e intensità
metabolica; cambia il fatto che la pendenza della
relazione tra frequenza cardiaca e intensità
metabolica è molto diversa, la pendenza della retta
dell'individuo allenato è molto più bassa rispetto a
quella del sedentario. Questo significa che, a parità
d'intensità metabolica richiesta, l'incremento di
frequenza cardiaca è più elevato nell'individuo
sedentario rispetto a quello allenato, ovvero sono
necessari un numero di battiti cardiaci maggiori per
pompare la stessa quantità di sangue nel torrente
circolatorio e per raggiungere la stessa quantità di
consumo di ossigeno. Qual'è il motivo di tutto ciò? La gittata sistolica di una persona allenata è
molto più alta rispetto alla persona sedentaria e quindi viene sfruttato di meno l'incremento della
FC; praticamente succede l'esatto contrario dello scompensato cardiaco, questo infatti non può
sfruttare l'aumento della GS e quindi aumenta
la PC incrementando la FC.
La gittata sistolica nell'esercizio fisico
può anche raddoppiare (nei soggetti allenati!),
ciò si sfrutta per aumentare la portata cardiaca.
In un soggetto sedentario quindi la GS
non arriva a raddoppiare ma al massimo può
aumentare del 20-30% rispetto alle condizioni
di partenza. L'allenato quindi aumenta la
portata cardiaca aumentando la FC di 3 volte e
la GS di 2 volte; si ha quindi un aumento della
PC di 6 volte(?); la persona sedentaria invece
non è in grado di aumentare la portata cardiaca
di così tanto proprio perché il suo incremento
di GS sarà massimo del 20-30% e quindi la portata cardiaca in questo caso non aumenterà per più di
3-4 volte rispetto a quella a riposo.
Ecco spiegata meglio la differenza tra un sedentario e un allenato: l'allenato sfrutta al meglio
la possibilità di incremento della gittata sistolica, cioè la possibilità di aumentare il volume di
sangue eiettato a ogni singola contrazione. Un'altra cosa importante da dire è che normalmente una
persona allenata, anche in condizioni di riposo presenta valori di gittata sistolica più alti rispetto a
quelli di un sedentario. I sedentari hanno valori di GS a riposo che vanno da 70-80 mL, mentre per
quanto riguarda gli sportivi sono stati visti valori che vanno anche a 120-130 mL, quindi hanno già
di base una gittata sistolica più alta rispetto a quella delle persone non allenate.
Come faccio ad aumentare la
gittata sistolica? Aumentando la
frazione di eiezione, che nei soggetti a
riposo è di circa 55-60; quindi, il cuore
quando si contrae a riposo non eietta tutto
sangue che ha dentro in quanto circa il
40% rimane dentro il cuore stesso.
Aumentando quindi la frazione di
eiezione aumenta la gittata sistolica.
il
In questo grafico vediamo queste
due sigle:
EDV → END DIASTOLIC
VOLUME → volume di fine diastole, cioè
volume che c'è dentro il cuore, in questo caso
il ventricolo sinistro, alla fine della diastole
ovvero prima che il cuore si contragga.
ESV → END SISTOLIC VOLUME
→ quantità di sangue presente nel ventricolo
alla fine della sistole ovvero quando il cuore
ha smesso di contrarsi.
La differenza tra ESV e EDV è la GS. Il
rapporto è la frazione di eiezione. Nella prima
parte del grafico quindi ci sarà una gittata
sistolica di circa 80 ml che aumenterà
progressivamente fino a valori di circa 120
mL, perchè il cuore si è “spremuto” meglio,
con più efficienza e maggiore forza di
contrazione. La frazione di eiezione quindi
passerà da valori di riposo di circa 55-60% a
valori che al massimo dell'esercizio possono
arrivare fino al 90%.
Questo però non mi basta per spiegare il
raddoppio della gittata sistolica in questi soggetti. Alcuni soggetti particolarmente allenati abbiamo
detto che sono in grado di raddoppiare la gittata sistolica rispetto al valore iniziale. Un motivo,
abbiamo detto, dato dall'aumento della frazione di eiezione, ma ciò non è sufficiente a giustificare
un raddoppiamento della GS; quale è quindi il secondo motivo? Il secondo motivo è dato dal fatto
che si ha un aumento del volume di fine diastole EDF. Questo succede solo per quegli atleti di
alto livello per i quali si arriva a un raddoppio della GS per questi due fattori citati prima;
negli atleti di medio livello ciò è dato invece solo da un aumento della frazione di eiezione
pertanto l'incremento della GS non è del 100% rispetto al valore iniziale, ma più basso (50-6070%). Gli atleti eccezionali quindi per arrivare ad un raddoppiamento della GS hanno un aumento
del volume di fine diastole cioè si ha un aumento del riempimento del cuore, della volumetria
cardiaca. Questo aumento della EDF è determinato anche da un aumento della contrattilità cardiaca
dovuto essenzialmente ad un aumento dell'attività del Simpatico e quindi ad un aumento della
scarica di Noradrenalina, ma è determinato anche dall'adrenalina in circolo, ovvero quella secreta
dalle surrenali.
In sintesi quindi, per aumentare la GS è necessario:
– un aumento della frazione di eiezione (atleti e sedentari; gli scompensati cardiaci non
incrementano per niente e partono da una frazione di eiezione di base ancora più bassa)
– “gonfiare” il cuore per aumentare il volume di fine diastole EDF e garantire un maggiore
ritorno venoso (meccanismo che riescono a sfruttare solo gli atleti di livello molto elevato)
Altro importante parametro
da analizzare nelle prove da
sforzo incrementale al
cicloergometro, dove però
non abbiamo grosse
differenze tra sedentari e
allenati, è la pressione
arteriosa: sistolica (SAP
Sistolic Arterial Pressure),
diastolica (DAP Diastolic
Arterial Pressure) e media
(MAP). La pressione
arteriosa media è il valore
più importante e si calcola
con l'equazione: MAP=
DAP+(SAP-DAP)/3.
Perché questa formula? Perché a riposo la sistole cardiaca dura circa 1/3 del ciclo cardiaco mentre
la diastole 2/3. La MAP in genere va da 80 a 110. Come vedete nel grafico, in generale in una prova
da sforzo, la pressione minima tende a rimanere costante o addirittura a scendere, sopratutto se lo
sforzo impiega grosse masse muscolari (per esempio correre); la SAP invece tende a salire portando
così a un lieve incremento anche della pressione arteriosa media. Come già detto questi parametri
non sono dipendenti dall'allenamento, non c'è differenza quindi tra persone sedentarie e persone
allenate. L'unica differenza è nelle persone ipertese le quali hanno la pressione che sale tanto. Uno
dei criteri per capire se una persona è ipertesa, nei casi borderline (in cui non è ben chiaro), oltre a
misurare la pressione a riposo si fa fare al paziente una prova da sforzo per vedere se la pressione
arteriosa media incrementa tanto oppure no; se a riposo si parte da una MAP di 80 mi aspetto che al
massimo arrivi a 100, se invece si arriva a 120-130 allora si tratta di una risposta ipertensiva. Per cui
la pressione arteriosa è un parametro importante da vedere durante la prova da sforzo non tanto
perché si ha una differenza tra persone allenate e persone sedentarie ma piuttosto per individuare
eventuali ipertensioni.
Vediamo altri parametri che possono essere studiati durante l'esercizio: SVR è l'acronimo di
Sistemic Vascular Resistance, ovvero le resistenze che risultano essere dovute al calibro medio delle
arteriole, e che possono essere calcolate semplicemente facendo il rapporto pressione/portata
cardiaca.
Vediamo quindi l'andamento della
portata cardiaca (CO) in L/min in un
soggetto non particolarmente allenato;
la pressione vediamo che tende a salire
in quanto si tratta sempre di una prova
da sforzo al cicloergometro. Vediamo
poi (in verde) l'andamento delle
resistenze vascolari sistemiche che
hanno una profonda riduzione durante
l'esercizio. Tutti gli esercizi dinamici
(ovvero quelli che prevedono una fase
di contrazione e una di rilassamento
muscolare) come correre, andare in bicicletta, nuotare, ecc. hanno questo tipo di andamento;
viceversa gli esercizi statici (per esempio sollevamento pesi), non hanno questi tipo di andamento
ma addirittura hanno un andamento contrario, cioè le resistenze tendono ad aumentare.
Perché negli esercizi dinamici c'è una profonda riduzione delle resistenze vascolari
sistemiche? Durante l'esercizio c'è una scarica simpatica, che in teoria dovrebbe vasocostringere e
quindi in teoria dovrebbe aumentare le resistenze, però, di fatto, troviamo una riduzione di queste
SVR. Questo succede perché nella muscolatura scheletrica che sta lavorando predomina il
controllo vascolare locale, cioè metabolico e la scarica adrenenergica che tende a vasodilatare;
nell'equilibrio totale dell'organismo nonostante ci sia una forte scarica simpatica predomina la
vasodilatazione a livello arterioso. In linea di massima la vasodilatazione predomina perché
l'apertura delle arteriole dei muscoli che stanno lavorando, sopratutto quando le masse muscolari
sono particolarmente grosse (bicipite, tricipite ecc.), è talmente tanta che predomina sulla
vasocostrizione che si ha sui visceri, sulla cute, e su tutti quei muscoli che non stanno lavorando.
L'effetto netto sarà quindi questo; nonostante ci sia una forte attivazione del sistema nervoso
simpatico, nell'organismo normale l'effetto predominante è la vasodilatazione. Se non ci fosse un
aumento contemporaneo della portata cardiaca che cosa succederebbe a fronte di questa
vasodilatazione? La pressione si abbasserebbe e si arriverebbe allo svenimento per mancanza
di perfusione cerebrale.
È quindi necessario un aumento della portata cardiaca a fronte di un esercizio per garantire un
apporto adeguato ai muscoli, e per evitare che la pressione scenda.
Ci sono poi un'altra tipologia di pazienti, ovvero quelli che vasocostringono eccessivamente,
esempio classico: lo scompensato cardiaco. Gli scompensati cardiaci tendono a vasocostringere
anche quei territori che dovrebbero essere vasodilatati, ovvero i muscoli; questo perché non
riescono ad incrementare la portata cardiaca. Per mantenere la pressione, siccome non riescono ad
aumentare la portata cardiaca, tentano di non abbassare le resistenze vascolari sistemiche, e possono
in questo modo andare a vasocostringere quei territori che dovrebbero essere vasodilatati in quanto
stanno lavorando; per cui si ha un deficit perfusorio anche in questi muscoli: in questo caso quindi
non comandano i muscoli ma comanda il sistema
simpatico. In questi soggetti si ha quindi una
vasocostrizione dei muscoli che devono lavorare e
smettono quindi di lavorare; una delle
conseguenze di ciò è lo sviluppo anche di miopatia
periferica. Si ha una progressiva atrofia muscolare
dovuta a deficit perfusorio e quindi questi soggetti
tendono a non muoversi più.
Estrazione di Ossigeno.
Come si calcola l'estrazione di O2 dell'organismo
in toto? Si calcola con l'equazione di Fick3 (flusso/estrazione arterovenosa). Applicata a questo
particolare caso l'equazione di Fick ci dice che l'estrazione è uguale al consumo di ossigeno diviso
la portata cardiaca; il consumo di ossigeno viene misurato con un metabolimetro, la PC viene
misurata con uno dei tanti sistemi che abbiamo (in questo caso è stata usata semplicemente la
calorimetria indiretta). Abbiamo visto che il cuore ha un'alta capacità di estrazione anche in
condizioni di riposo (130mL, sui 200 che ci sono, vengono estratti per ogni L di sangue che arriva) .
Nel muscolo la situazione è diversa: a riposo il muscolo estrae poco ossigeno(40-50 mL) mentre
durante l'esercizio questa estrazione aumenta progressivamente (vedete nel grafico che si arriva fino
a 150 mL;) fino a triplicare l'estrazione di Ossigeno. Vengono messi in atto quindi dei meccanismi
di drive dal globulo rosso al tessuto, che sono in grado di triplicare quindi l'estrazione di O2.
Un litro di sangue arterioso contiene circa 200 mL di O2 nelle persone sane (non
anemiche nè policitemiche); questo è il valore che si ha normalmente, purché uno non si dopi con
EPO (eritropoietina) e quindi abbia valori elevati di Hb.
Nei soggetti più allenati il coefficiente di utilizzazione dell'ossigeno non supera mai
l'85% il che vuole dire che anche le persone più allenate non estrarranno più di 170 mL di O2
su un L di sangue arterioso. Questo è un meccanismo allenabile: chi fa attività di endurance
(maratoneti, ciclisti) arriva ad avere delle estrazioni di ossigeno più alte rispetto a quella delle
persone sedentarie. Tutto ciò dipende da vari fattori: capillarizzazione dei muscoli, numero di
mitocondri per fibra ecc.
Riassunto: Cosa succede durante l'esercizio fisico dal punto di vista degli aggiustamenti (in
acuto)? Aumenta la portata cardiaca per aumento della frequenza cardiaca e della gittata
sistolica; aumenta la contrattilità che si traduce come un aumento della frazione di eiezione. Non
solo: quelli bravi possono anche incrementare il volume di fine diastole (nella maggior parte delle
persone rimane invece costante); le resistenze vascolari periferiche nelle persone sane scendono
però il tutto è compensato da un aumento della PC che impedisce un abbassamento della pressione.
Infine aumenta l'estrazione di ossigeno.
Un qualsiasi problema in uno di questi parametri determina un problema di incremento della
PC. L'estrazione di ossigeno può essere invece un problema in chi soffre di miopatia per cui anche
loro vanno incontro a problemi nell'affrontare l'esercizio.
Da che cosa sono governati
tutti questi meccanismi? Dal
Sistema Nervoso Centrale.
Ricordiamo che i centri di
controllo cardiocircolatori
sono nel bulbo che integra
una serie di informazioni
provenienti dai barocettori,
corteccia motoria, recettori
muscolari... e poi regola la
risposta del cuore e dei vasi
sulla base di tutte queste
3 La formula è questa:
F= Qx/PAx-PVx
dove F= flusso, Qx= quantita di sostanza che scompare (o compare) dal plasma, PAx e PVx=
concentrazione della sostanza nel sangue arterioso e venoso rispettivamente.
informazioni (tutto ciò sarà argomento di seminario → ultima lezione di fisiologia). Il midollo
allungato vuole sapere anche qual'è la pressione arteriosa (→ barocettori) e il tipo di movimento che
si sta facendo.
ADATTAMENTI MORFOLOGICI INDOTTI DALL'ESERCIZIO (A LUNGO
TERMINE)
Finora abbiamo visto gli aggiustamenti, ora vediamo gli adattamenti, cioè ciò che succede
alla lunga sull'apparato cardiocircolatorio nelle persone che fanno esercizio cronicamente, cioè sugli
atleti.
A livello cardiaco abbiamo vari
tipi di adattamento in seguito ad
esercizio cronico (risultati che si
ottengono dopo anni).
Normalmente in chi fa esercizio
di endurance (ciclisti, maratoneti,
atleti che fanno esercizi di lunga durata
a discreta intensità), il cuore si ingrossa
in maniera proporzionata cioè aumenta
sia da un punto di vista volumetrico,
sia dal punto di vista dello spessore
delle camere cardiache e quindi in
questo caso il rapporto M/V rimane
uguale a 1, ovvero tanto aumenta la
massa, tanto aumenta il volume. Si
tratta quindi di un'ipertrofia
appropriata in cui massa e volume aumentano in maniera proporzionata.
Chi fa invece esercizio di potenza, esercizi di tipo isometrico (esempio sollevatori di pesi), ha
un tipo di adattamento per il quale il cuore aumenta il proprio spessore ma non aumenta il volume e
quindi il rapporto M/V è >1 (ipertrofia sproporzionata). Questo succede perché il lavoro che
viene fatto non è un lavoro “di flusso”, cioè non è un lavoro in cui si deve garantire un grande
apporto di ossigeno nei muscoli, perché i muscoli “si tappano” per effetto della contrazione
muscolare. Lo sforzo muscolare è uno sforzo contro pressione per cui il cuore deve vincere la
pressione aumentata, sviluppando quindi questo tipo di ipertrofia detta anche concentrica.
L'ipertrofia inadeguata si ha in una patologia che si chiama cardiopatia dilatativa, nella
quale aumenta il volume ma si riduce molto lo spessore o la massa del ventricolo. In questa
patologia si vedono quindi questi cuori voluminosi i quali però non sono così perché si tratta di
atleti ma perché sono in realtà dei cuori “sfiancati”.
Gli adattamenti quindi possono essere distinti in due tipi: ipertrofia eccentrica (A-C) e
ipertrofia concentrica (B), determinati dal diverso tipo di esercizio fisico. Nella figura B infatti è
raffigurato una persona che fa sollevamento pesi in quanto questi sviluppano un aumento dello
spessore del cuore, al quale non segue un aumento del suo volume (m/v>1).
Lo spessore delle pareti del cuore in questi atleti viene studiato dal punto di vista clinico per
determinare se questo accrescimento delle pareti è normale per il lavoro che si sta facendo oppure è
un accrescimento patologico (cardiomiopatia ipertrofica) dovuto all'ingrossamento e al replicarsi
delle fibre muscolari per fattori genetici.