1. INTRODUZIONE
Le anomalie molecolari responsabili della trasformazione tumorale
consistono in mutazioni multiple a carico di classi fondamentali di geni coinvolti
nella moltiplicazione e differenziazione delle cellule, i proto-oncogeni e i geni
oncosoppressori. Le proteine espresse dai proto-oncogeni stimolano la cellula a
progredire nel suo ciclo cellulare (aumento di volume, replicazione del DNA e
divisione cellulare); i geni oncosoppressori, invece, inibiscono la crescita della
cellula. I proto-oncogeni modificati da mutazioni provocate da agenti esogeni
(chimici, fisici e virali) danno luogo agli oncogeni, la maggior parte dei quali
codifica per una proteina tirosin-chinasi. Quasi tutte le mutazioni associate agli
oncogeni producono una perdita dell’inibizione dell’attività catalitica e, così
facendo, alterano le funzioni che controllano la divisione cellulare1.
Numerosi polipeptidi, come i fattori di crescita, i fattori di
differenziamento e gli ormoni, si sono dimostrati componenti cruciali nella
proliferazione e nel differenziamento cellulare. Molti di questi fattori mediano la
loro azione legandosi ed attivando specifici recettori di membrana associati ad
attività tirosin-chinasica. Il recettore trasferisce il messaggio chimico
extracellulare all’apparato metabolico intracellulare, innescando, attraverso una
cascata di reazioni culminanti nella divisione cellulare, la risposta della cellula
bersaglio. Questi recettori, quando attivati dal ligando, sono proteine in grado di
catalizzare il trasferimento di un gruppo fosfato terminale di una molecola di
ATP al gruppo ossidrilico laterale di un residuo di tirosina della molecola
substrato o della porzione carbossi-terminale del recettore stesso. L’attività
chinasica è indispensabile per la trasduzione del segnale e per l’induzione della
risposta cellulare2. La forma fosforilata di una proteina enzimatica o strutturale
rappresenta in genere la forma attivata, mentre la proteina defosforilata è la sua
controparte inattiva3.
1
Il legame con il ligando provoca la dimerizzazione del recettore, la
fosforilazione della tirosina e poi della chinasi. Tali eventi permettono
l’attivazione di molte molecole effettrici, tra cui le MAP chinasi attive, che
stimolano la sintesi e l’attivazione di fattori trascrizionali, come FOS e JUN, i
quali regolano il ciclo cellulare e che sono, pertanto, coinvolti nella
tumorigenesi. I recettori tirosin-chinasici delle cellule tumorali, infatti, sono
permanentemente
dimerizzati,
quindi
i
recettori
mutati
trasmettono
continuamente segnali mitogeni e ciò provoca una crescita incontrollata e
incontrollabile2 (Fig. 1).
Fig. 1: Dimerizzazione del recettore a
tirosin-chinasi.
Oltre ai recettori a tirosin-chinasi, che sono proteine integrali della
membrana plasmatica, esistono anche tirosin-chinasi definite “non-recettori”, in
quanto sono intracitoplasmatiche, localizzate subito all’interno della membrana
cellulare (come la famiglia delle Src tirosin-chinasi). Esse, comunque, sono
associate con varie membrane, a livello delle quali svolgono la loro attività
chinasica4.
Esistono diverse famiglie di tirosin-chinasi, sia recettoriale che nonrecettoriali, classificate in base all’oncogene dal quale derivano (Fig. 2).
2
Fig. 2: Alcuni esempi di recettori a tirosin-chinasi. In rosso viene evidenziata la
regione conservata responsabile della attività chinasica.
I recettori a tirosin-chinasi sono proteine che presentano un solo dominio
transmembrana; sono dotate di attività chinasica intrinseca. Tra questi grande
rilievo hanno le famiglie di PDGFR, IGFR, EGFR e VEGFR.
Le tirosin-chinasi citoplasmatiche coinvolte nella trasmissione del segnale
comprendono essenzialmente dieci famiglie, tra cui Src, Abl e Jak hanno
suscitato particolare interesse da parte dei gruppi di ricerca.
Analizzando la sequenza amminoacidica delle varie tirosin-chinasi si è
osservato che esse, nonostante la diversità in dimensioni e composizione,
presentano una sequenza comune di circa 250 amminoacidi: questa regione è
stata riconosciuta come responsabile dell’attività tirosin-chinasica3.
I domini catalitici delle tirosin-chinasi eucariotiche, così come quelli delle
Ser/thr chinasi, presentano una struttura e una sequenza altamente conservate. Il
dominio chinasico è costituito da una struttura bilobata. Il lobo N contiene un
foglietto ed una -elica conservata (elica C). Il lobo C è fortemente elicoidale.
All’interfaccia tra i due lobi è presente una tasca di legame per l’ATP, formata
da residui estremamente conservati, e la struttura catalitica. Inibitori di piccole
3
dimensioni delle protein-chinasi si legano al dominio chinasico a livello dello
spazio tra i due lobi, spiazzando l’ATP5 .
Al contrario di quanto avviene nelle cellule normali, dove normalmente
funzionano soprattutto chinasi proteiche, quali serina- e treonina-chinasi, e dove
le tirosin-chinasi sono quasi inesistenti, nelle cellule tumorali il livello di attività
tirosin-chinasica può aumentare di 10-20 volte3. Per questo motivo le tirosine
chinasi e i meccanismi di trasduzione del segnale possono essere identificati
come potenziali bersagli per la progettazione di farmaci6.
Un approccio per l’ideazione di farmaci antineoplastici può essere quello
di interrompere la trasmissione del segnale proliferativo, in modo da prevenire la
divisione cellulare (Fig. 3).
Fig. 3: La via di trasduzione dei segnali di una cellula comprende molte componenti che,
se alterate, possono indurre la crescita tumorale.
La
comprensione
dei
meccanismi
molecolari
implicati
nella
trasformazione neoplastica permette la progettazione di nuovi farmaci ad azione
più specifica, che potrebbero essere in grado di ripristinare la normalità nelle
cellule trasformate o di eliminarle selettivamente senza danneggiare quelle
sane2.
4
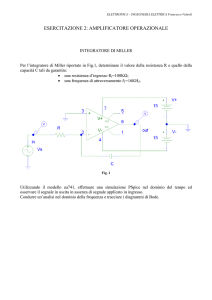
2. INIBITORI DELLE TIROSIN-CHINASI
Le nuove linee di ricerca farmacologica si sono rivolte alla identificazione
di agenti (terapia target) in grado di interferire in maniera selettiva contro
bersagli molecolari specifici al fine di aumentare la selettività del bersaglio e di
ridurne gli effetti collaterali sistemici7.
È evidente che l’inibizione selettiva di singole molecole segnale in
determinati punti della trasduzione è fondamentale per ottenere buoni risultati
dal punto di vista terapeutico.
STI-571 (Gleevec o Imatinib) e Iressa (ZD1839) sono stati i primi farmaci
inibitori della chinasi ad essere immessi in commercio. Sebbene siano stati
studiati diversi inibitori che agiscono mediante un intermedio allosterico e che
non sono competitivi con l’ATP, tutti i farmaci finora commercializzati come
inibitori chinasici hanno come bersaglio il relativamente conservato sito di
legame per l’ATP. Attualmente più di 30 inibitori che competono con l’ATP
sono in varie fasi di studio clinico8(Fig. 4).
Fig. 4: Azione degli inibitori
delle tirosin-chinasi.
Ad oggi sono state cristallizzate 7 strutture del Gleevec co-cristallizzato
con alcune tirosin-chinasi (Fig. 5).
5
Fig. 5: Struttura tridimensionale del Gleevec a
migliore risoluzione (1.57 Å) co-cristallizzata con
la Abl tirosin-chinasi (codice PDB: 1XBB).
STI-571 si è dimostrato efficace nella cura della leucemia mieloide
cronica (CML) in pazienti resistenti all’interferone alfa e che non possono
effettuare il trapianto allogenico di midollo osseo. Essendo estremamente
selettivo nei confronti delle cellule tumorali, risparmiando quelle sane, presenta
un ridotto numero di effetti collaterali.
Fig. 6: Schema del meccanismo di azione del Gleevec.
La CML è caratterizzata nel 95% dei casi dalla presenza del cosiddetto
cromosoma Philadelphia, una traslocazione reciproca tra i bracci lunghi dei
cromosomi 9 e 22. Tale mutazione somatica determina la fusione tra un
segmento del gene Bcr, dal cromosoma 9, ed una regione a monte del secondo
esone del gene c-Abl, dal cromosoma 229. c-Abl è una proteina piuttosto
grande, costituita da circa 1150 residui amminoacidici. La metà N-terminale di
c-Abl (circa 530 residui) presenta un’identità di sequenza del 42% con la
6
famiglia delle Src tirosin-chinasi (escluso il dominio N-terminale in quanto tale),
oltre a condividere con essa una simile organizzazione all’interno del dominio,
che contiene due unità di legame per il peptide (regioni SH2 e SH3) seguite da
un dominio tirosin-chinasico. Comunque, c-Abl si distingue dalle Src chinasi in
quanto manca di un particolare residuo di tirosina, il quale segue il dominio
chinasico di Src. La fosforilazione di tale residuo di tirosina provoca
l’inattivazione delle Src chinasi. La metà C-terminale di c-Abl contiene il DNA
e i domini di legame per l’actina intervallati da siti di fosforilazione ed altre
corte regioni di riconoscimento, inclusi frammenti ricchi di prolina e segnali di
localizzazione nucleare. c-Abl codifica per una tirosin-chinasi non recettoriale,
la quale presenta un’attività fortemente controllata nelle cellule normali5(Fig.
7).
Fig. 7: Strutture del dominio chinasico di c-Src, c-Abl 1b e Bcr-Abl a confronto.
Principale responsabile della comparsa della malattia, invece, è proprio la
proteina chimerica sregolata sintetizzata dal cromosoma Philadelphia, l’enzima
tirosin-chinasi Bcr-Abl, il quale, essendo in stato di continua attivazione,
favorisce la proliferazione incontrollata delle cellule ematopoietiche e, in più, ne
rallenta l’eliminazione fisiologica, frenando il segnale di apoptosi. La stessa
anomalia genetica si riscontra anche nel 5-10 % degli adulti con leucemia
linfoblastica acuta, che risulta positivo al cromosoma Philadelphia5.
7
Fig. 8: Schema del sito sul quale va ad agire il Gleevec e pathways da esso inibiti.
È stata identificata una serie di inibitori, basati sulla classe di farmacofori
delle 2-fenilaminopirimidine, i quali presentano alta affinità e specificità per
Abl10. Il più potente tra questi, STI-571 (Gleevec o Imatinib), come già
accennato, si è mostrato un ottimo agente terapeutico per la CML, in quanto ben
tollerato dal 96% dei pazienti trattati durante i “trials” clinici9.(Fig. 9)
Fig. 9: Struttura chimica dell’inibitore di Abl STI-571 (1) e di una sua variante
(2), nella quale manca il gruppo piperazinilico; tale variante viene usata per
poter cristallizzare la struttura del dominio catalitico della tirosin-chinasi Abl
complessato con il farmaco.
8
L’Imatinib inibisce in modo altamente specifico la Bcr-Abl, mentre si è
dimostrato completamente inattivo nei confronti delle Ser/Thr-chinasi e molte
delle tirosin-chinasi, fatta eccezione per due recettori a tirosin-chinasi correlati,
il recettore PDGF (Platelet-Derived Growth Factor) e c-Kit11. La sua notevole
specificità sottolinea la sua efficacia nel trattamento delle malattie
mieloproliferative croniche che coinvolgono l’attivazione del recettore PDGF e
dei tumori stromali gastrointestinali che prevedono l’attivazione del recettore cKit12,13. Il farmaco, infatti, compete con l’ATP, ligando fisiologico della BcrAbl, per lo stesso sito all’interno della proteina. In presenza dell’inibitore, la
tirosin-chinasi non è più in grado di trasmettere alle cellule del midollo i segnali
difettosi, che causano l’iperproduzione di globuli bianchi. Dal momento che la
tirosin-chinasi Bcr-Abl è espressa solo nelle cellule malate, l’Imatinib è una
sorta di “proiettile antineoplastico intelligente” 11.
Un gruppo di studiosi9 ha dimostrato come sia di cruciale importanza per
la formazione del complesso tra il dominio catalitico di Abl e STI-571 è la
permanenza nella conformazione inattiva da parte della chinasi, stato in cui lo
“anello di attivazione” centralmente localizzato non è fosforilato. La
conformazione di tale anello si distingue da quella delle proteine chinasi attive,
così come la forma inattiva delle Src chinasi strettamente correlate. I composti
che sfruttano i tipici meccanismi di inattivazione delle singole protein-chinasi
possono realizzare sia alta affinità che grande specificità (Fig. 10).
Se messo a confronto con l’inibitore pirazolo-pirimidinico PP1 legato ad
Hck Src-chinasi, STI-571 si estende molto di più nel dominio catalitico, e il suo
gruppo piridinilico è inserito al di sotto dell’C elica nel lobo N-terminale della
chinasi. Il composto è attorcigliato all’ammino-gruppo secondario, e divarica la
regione N-terminale altamente conservata dello “anello di attivazione”9(Fig.
11).
9
Fig. 10: Struttura tridimensionale del dominio
chinasico di Abl in complesso con la variante di
STI-571 (codice PDB: 1FPU).
Fig. 11: Loop di attivazione di Abl. La Tyr393
rappresenta il sito di fosforilazione, anche se la
forma di Abl cristallizzata non è fosforilata.
Il “loop” di attivazione controlla l’attività catalitica nella maggior parte
delle chinasi mediante il brusco spostamento in diverse conformazioni in
maniera fosforilazione-dipendente. In tutte le chinasi attive, il “loop” è
stabilizzato in una conformazione aperta dalla fosforilazione sui residui di
serina, treonina o tirosina all’interno del sito, e in questa conformazione un
filamento nell’anello funge da “piattaforma” per il legame del substrato9.
10
Fig. : Stereochimica della tasca di legame per il nucleotide di Abl. Si è osservata una forte
densità elettronica nella variante di STI-571, la quale occupa il sito dove la base di adenina
dell’ATP generalmente si lega.
Tre residui altamente conservati nella regione N-terminale di questo sito
(un gruppo Asp-Phe-Gly, definito DFG, corrispondente ai residui 381-383 in
Abl) sono, così, tenuti in una conformazione che è appropriata per il legame
dello ione metallico della catena laterale dell’aspartato. Questo stato attivo
dell’anello è molto simile in tutte le strutture conosciute delle chinasi attive.
Esiste, invece, una grande diversità nelle conformazioni di questo sito nelle
protein-chinasi inattive, nelle quali spesso l’anello ostacola il legame del
substrato. Le disposizioni a gomito del “loop” nella regione N-terminale, inoltre,
modificano la posizione della triade Asp-Phe-Gly, inibendo, così, la capacità
della chinasi di legare l’ATP in modo proficuo9.
La Tyr393 nell’anello di attivazione è il più importante sito di
fosforilazione in Abl.
Il “loop” di attivazione è avvolto all’interno del sito attivo della chinasi e
la Tyr393 forma un legame ad idrogeno con l’Asp363, una catena laterale
fortemente conservata che risulta fondamentale per la catalisi. Tyr393 viene
disposto nel sito attivo mediante un piccolo -filamento antiparallelo, il quale fa
parte di una porzione dell’anello di attivazione. Il sito di attivazione mima il
meccanismo di legame del substrato, così come era stato precedentemente
osservato nell’IRK (insulin receptor tyrosine kinase) (Fig. 12).
11
Fig. 12: Il confronto dei domini catalitici di Abl e
IRK inattiva mostra che la parte centrale
dell’activation loop in entrambe le chinasi
occlude la bocca del dominio catalitico e
interferisce con l’utile legame dei substrati del
peptide in maniera simile.
È importante sottolineare che, ad eccezione della regione di ancoraggio
dell’N-terminale, STI-571 non interagisce direttamente con l’anello di
attivazione. La sorprendente somiglianza tra la conformazione del sito di
attivazione e il modo in cui i substrati del peptide si legano alle tirosin-chinasi
suggerisce che il “loop” si trova in una conformazione autoinibitoria naturale.
Sebbene Tyr393 si trovi posizionato esattamente come in un peptide
substrato, il dominio della chinasi non è in una conformazione adatta per il
trasferimento del fosfato alla tirosina, dal momento che il movimento interno
dell’anello di attivazione è accoppiato allo spostamento del gruppo Asp-Phe-Gly
lontano dalla conformazione attiva sia in Abl che in IRK (Asp381 si orienta
lontano dal sito attivo). Nonostante queste somiglianze, STI-571 risulta inattivo
nei confronti di IRK, molto probabilmente perché una catena laterale (Thr 315)
che forma un particolare contatto con l’inibitore alla periferia del sito di legame
per il nucleotide di Abl non è conservato in IRK9.
Al contrario di quanto succede nel sito di attivazione, dove
verosimilmente STI-571 si lega ad una forma naturale della molecola di Abl, le
interazioni del farmaco nel lobo N-terminale della chinasi sembrano far parte di
un meccanismo di attacco indotto. L’anello tra i primi due filamenti , che
normalmente lega i gruppi fosfato dell’ATP, si ripiega in modo da aumentare la
complementarietà di superficie con il farmaco. Questo lembo viene mantenuto
in posizione da un legame ad idrogeno mediato dall’acqua tra Tyr 253, un residuo
12
nel lobo N-terminale della chinasi che segue immediatamente il “loop” 1-2, e
la catena laterale di Asn322.
L’inibitore, inoltre, interagisce con la chinasi attraverso legami ad
idrogeno, alcuni dei quali conferiscono la specificità (Fig. 13).
Fig. 13: Rappresentazione schematica delle interazioni che la chinasi Abl forma con la
variante di STI-571.
L’azoto nell’anello della piridina che è attaccato alla metà della pirimidina
accetta un legame a idrogeno dall’ammide di Met318, che normalmente forma un
legame a idrogeno con l’azoto N1 dell’ATP. La catena laterale di Thr 315 forma
un legame a idrogeno con il gruppo amminico secondario nell’inibitore. Questo
residuo è rimpiazzato da una metionina in molte protein-chinasi, come in IRK.
La metionina non può formare questo legame a idrogeno, e la sua catena laterale
potrebbe anche interferire con il legame del “mezzo-fenile” di STI-571. La
presenza di Thr315 è, quindi, un requisito chiave affinchè questa classe di
composti sia capace di inibire Abl9.
13
Una coppia di ioni tra due catene laterali strettamente conservate (Lys 271 e
Glu286 in Abl) è una tipica caratteristica delle conformazioni attive delle proteinchinasi. Tale coppia di ioni viene smembrata nelle conformazioni inattive di
molte protein-chinasi, come la Src e le chinasi ciclina-dipendenti, ma non nel
complesso di STI-571 con Abl. Invece, una rete di legami a idrogeno, che
coinvolge le catene laterali dei residui di Lys271 e Glu286, così come la catena
principale di Asp381, il gruppo ammidico acido dell’inibitore e due molecole di
acqua, stabilizza ulteriormente il legame.
È presente un certo numero di interazioni di van der Waals tra i residui
proteici Tyr253, Leu370, Phe382, Met290 e Ile313, e gli anelli aromatici dell’inibitore,
che determina un’eccezionale livello di complementarietà di superficie. Lo
stretto attacco tiene fortemente conto di alcuni cambiamenti o sull’inibitore o sul
dominio chinasico senza compromettere l’affinità di legame. Al contrario,
alterazioni nelle sequenze di altre protein-chinasi nelle regioni che compongono
il sito di legame, come il rimpiazzamento della Thr 315 da parte della metionina,
potrebbe interferire col legame9.
In uno studio successivo è stato messo a confronto l’imatinib con un altro
inibitore di Abl, PD1739555(Fig. 14).
Fig. 14: Strutture chimiche di STI-571 e di PD173955.
Entrambi i farmaci si legano al sito di legame per l’ATP del dominio
chinasico, ma lo fanno in modi diversi.
Una conformazione inattiva del dominio chinasico di Abl, come già detto,
sembra essere cruciale per la selettività di STI-571, poiché la struttura studiata
differisce dalle conformazioni inattive di altre tirosin-chinasi, come le Src
14
chinasi strettamente correlate, nei confronti delle quali l’imatinib si mostra
inefficace. Probabilmente un eventuale effetto di STI-571 nei confronti delle
forme attivate di Bcr-Abl deriva dalla natura dinamica delle molecole
chinasiche, che hanno la capacità di passare repentinamente dalla forma attiva a
quella inattiva in modo transitorio, permettendo all’inibitore di raggiungere
l’ingresso del sito di legame5.
PD173955 è un inibitore appartenente alla famiglia di composti con un
centro pirido-2,3-dpirimidinico14. Al contrario di quanto avviene per
l’imatinib, esso si lega ad una conformazione di Abl nella quale il “loop” di
attivazione è simile a quello di una chinasi attiva; esso, pertanto non viene
influenzato dalla conformazione del sito di attivazione della chinasi, attiva o
inattiva che sia5. PD173955, inoltre, riesce ad avere una forza inibitoria di
almeno 10 volte superiore a quella dell’imatinib, dal momento che, mentre STI571 lega in maniera specifica una determinata conformazione di Abl, PD173955
ha la capacità di legare forme diverse di Abl. Così come STI-571, esso
rappresenta un forte inibitore del recettore tirosin-chinasi c-Kit, ma inibisce
anche in modo molto efficace le Src chinasi, al contrario dell’imatinib15.
Sono state determinate le strutture tridimensionali del dominio chinasico
di Abl in complesso con STI-571 e PD173955 a risoluzioni pari a 2.1 e 2.6 Å
(codici PDB: 1IEP e 1M52) rispettivamente. Entrambe le strutture rappresentano
la forma non fosforilata di Abl. In entrambi i cristalli il dominio chinasico risulta
essere molto simile nel suo complesso con l’architettura bilobata tipica sia delle
Ser/Thr- che delle tirosin-chinasi eucariotiche. I residui 225-350 compongono il
lobo N della chinasi, mentre i residui 354-498 comprendono il lobo C.
Esiste una regione in cui la struttura del dominio chinasico risulta
marcatamente differente nei due complessi di STI-571 e PD173955. Tale
diversità è ristretta al “loop” di attivazione, più in particolare nella regione
compresa tra i residui 381-402 in Abl, che rappresenta un elemento di
regolazione localizzato centralmente nelle protein-chinasi5.
15
Mentre le conformazioni delle protein-chinasi attive sono molto simili,
nelle varie forme inattive delle chinasi di diverse sottofamiglie sono visibili
notevoli differenze (Fig. 15).
Fig. 15: Cambiamenti conformazionali
all’interno del sito di attivazione in
seguito al legame delle protein-chinasi
Tali differenze riguardano le alterazioni nell’orientazione della zona tra i
due lobi e la disposizione dell’elica C nel lobo N. Un aspetto fondamentale
della transizione conformazionale tra lo stato attivo e inattivo è l’anello o il
segmento di attivazione, che è di lunghezza e sequenza variabili e rappresenta
spesso il sito di fosforilazione nel dominio chinasico5.
Nelle protein-chinasi che si trovano in uno stato completamente attivo, il
sito di attivazione è in una forma estesa o aperta, che presenta due caratteristiche
di grande importanza. Innanzi tutto, un residuo di acido aspartico (Asp381 in Abl)
all’interno del frammento Asp-Phe-Gly (DFG) al segmento N-terminale
dell’anello di attivazione è posizionato in modo tale da interagire con uno ione
magnesio che coordina i gruppi fosfato dell’ATP. Inoltre, il resto dell’anello si
16
trova lontano dal centro catalitico, cosicchè la porzione C-terminale del sito di
attivazione funga da “piattaforma” per il legame del substrato5(Fig. 16).
Fig. 16: Rappresentazione della struttura del dominio chinasico di Abl in complesso
con STI-571 e PD173955 (codici PDB: 1IEP e 1M52). La regione DFG è localizzata
all’N-terminale del loop di attivazione.
Nel complesso in cui STI-571 si trova legato all’Abl, la regione Nterminale dell’anello di attivazione, incluso il segmento DFG, si trova
totalmente ruotata rispetto alla posizione che assume nella forma attiva, in modo
che Phe382 del frammento DFG è rivolto al sito di legame per l’ATP al posto di
Asp381. La conformazione alterata di Phe382 è la caratteristica principale che
rende possibile il tipico legame di STI-571. L’altra parte del “loop” di
attivazione assume una conformazione in cui la regione circostante alla Tyr 393
(corrispondente al sito di fosforilazione che in questa struttura cristallizzata non
è fosforilata) mima un substrato che si lega all’enzima, bloccando, così, il sito
attivo.
Nel complesso tra PD173955 e Abl, la maggior parte dell’anello di
attivazione si trova in una forma molto simile a quella delle protein-chinasi
attive, lasciando il centro catalitico dell’enzima non bloccato, libero. La
conformazione del gruppo DFG appare, invece, diversa da quella delle chinasi
17
attive; probabilmente questa è una conseguenza dello stato non fosforilato
dell’enzima5 (Fig. 17).
Fig. 17: (A) A sinistra, confronto tra STI-571 (verde) e la sua variante (rosso) legate
ad Abl. A destra, confronto tra STI-571 (verde) e PD173955 (magenta) legate ad Abl.
(B) Confronto tra il loop di attivazione di Abl legato a STI-571 (verde), a PD173955
(rosso) e Lck attiva (magenta).
STI-571 è costituito da una struttura centrale, alla quale sono attaccati un
sostituente piridinico all’estremità del lato sinistro, e un legame peptidico
seguito da un anello fenilico e un anello piperazinilico a destra (Fig. 14). Il
farmaco attraversa completamente la regione centrale della chinasi, da una parte
all’altra. Soltanto la parte sinistra (anelli di piridina e pirimidina) di STI-571,
però, chiude la regione dove normalmente si lega l’adenina dell’ATP. Il resto
del composto penetra all’interno del centro idrofobico della chinasi e si incunea
tra il “loop” di attivazione e l’elica C, bloccando la chinasi in una
18
conformazione inattiva. In totale, il farmaco forma sei legami ad idrogeno con la
proteina e la maggior parte dei contatti avvengono mediante interazioni di van
der Waals5 (Fig. 18).
Fig. 18: Rappresentazione schematica dalle interazioni create da STI-571 e da PD173955 con Abl.
Il PD173955 presenta un anello biciclico centrale e due sostituenti, un
anilino-tiometile sulla sinistra ed un diclorofenile sulla destra. È, quindi, una
molecola più piccola rispetto a quella dell’imatinib e non si estende così in
profondità all’interno del dominio chinasico. Il suo anello pirido-pirimidinico
occupa il posto dove si legano anche gli anelli pirimidinico e piridinico di STI571; il gruppo fenilico ed il tiometile si estendono esternamente nel solvente.
L’anello diclorofenilico si sistema in una posizione simile a quella del fenile di
STI-571. Anche in questo caso la maggior parte delle interazioni tra PD173955
e la proteina sono di van der Waals5.
È interessante notare come, sebbene STI-571 e PD173955 si leghino
essenzialmente allo stesso sito di Abl, la differenza tra le loro dimensioni
provoca dei cambiamenti anche nelle caratteristiche di legame. Mentre STI-571
interagisce con 21 residui della proteina, PD173955 viene a contatto con solo 11
residui e risulta, nonostante ciò, significativamente più efficace nella sua azione
inibitoria nei confronti delle tirosin-chinasi.
Mediante i modelli dei complessi di STI-571 e PD173955 con le
conformazioni di Abl sia aperta (dove il “loop” di attivazione si protrae verso
l’esterno della proteina in una forma estesa, come in una chinasi attiva) che
19
chiusa (in cui il “loop” si attorciglia all’interno della proteina e mima il legame
col substrato) si è potuta analizzare l’azione dei due farmaci sull’enzima (Fig.
19).
Fig. 19: (A) Modello delle interazioni di
van der Waals che PD173955 forma con
il dominio chinasico di Abl complessato
con STI-571. (B) Modello delle
interazioni di van der Waals che
PD173955 forma con il dominio
chinasico di Abl complessato con STI571.
PD173955 nella forma inattiva di Abl può racchiudersi all’interno della
tasca di legame senza avere ulteriori scontri con gli atomi della proteina,
sebbene siano necessarie piccole variazioni del “P-loop”. Questo si è visto
essere estremamente flessibile grazie al suo alto contenuto di glicina. La
conformazione del P-loop, quindi, è probabilmente indotta dalla presenza
dell’inibitore, che provoca un piccolo spostamento a livello della Tyr 253,
affinchè si possa inserire il gruppo fenilico contenente zolfo di PD173955, che è
rivolto verso il solvente5.
In marcato contrasto, STI-571 non può sistemarsi nella forma di Abl
rimodellata da PD173955. Oltre all’ammino-gruppo secondario di STI-571, sia
l’anello fenilico che l’anello piperazinilico collidono con i residui del “loop” di
attivazione nella conformazione aperta, più in particolare con Asp381 e Leu384.
20
Così, PD173955 può probabilmente legarsi ad Abl a prescindere dalla
conformazione del sito di attivazione, mentre STI-571 ha bisogno che questo si
trovi nella forma chiusa. Una volta che STI-571 si è legato ad Abl, si blocca tra
l’anello di attivazione e l’elica C, evitando che avvenga un cambiamento
conformazionale. Per quanto riguarda PD173955, le sue piccole dimensioni non
richiedono che il sito di attivazione debba rimanere bloccato5.
È interessante notare come in realtà STI-571 presenti un’ulteriore metodo
per legarsi al dominio chinasico delle tirosin-chinasi.
La tirosin-chinasi della milza (Spleen tyrosine kinase, Syk) appartiene a
quella classe di protein-chinasi definite non-recettori indispensabili per la
trasmissione del segnale dagli immunorecettori in varie cellule ematopoietiche.
La fosforilazione di due residui di tirosina all’interno del “loop” attivazione del
dominio catalitico della Syk chinasi rappresenta proprio il passaggio
fondamentale per la trasmissione del segnale. L’attività enzimatica di Syk in
vitro, però, non dipende dalla fosforilazione; l’enzima, infatti, adotta una
conformazione dell’anello di attivazione presente in modo specifico nelle
tirosin-chinasi fosforilate e attivate, motivo per cui Syk non richiede la
fosforilazione per essere attivata. È stato osservato che il Gleevec (STI-571 o
Imatinib) inibisce il dominio chinasico isolato sia della Syk non fosforilata sia
dell’Abl fosforilata con una potenza paragonabile. Bisogna, però, sottolineare il
fatto che STI-571 lega Syk in modo particolare, cioè con una cis-conformazione
compatta, che varia notevolmente dal modo di legame osservato con l’Abl non
fosforilata, che è la forma di Abl più sensibile all’Imatinib. Questo suggerisce
l’esistenza di due distinti modi di legame per il Gleevec: un’estesa, transconformazione caratteristica dello stretto legame alla forma inattiva di una
protein-chinasi e una compatta cis-conformazione tipica del legame più debole
alla forma attiva16.
La famiglia delle Syk tirosin-chinasi è caratterizzata dalla presenza di due
domini SH2 N-terminali, una regione linker ed un dominio catalitico C21
terminale. Il lobo N-terminale è costituito da un foglietto a cinque filamenti e
da un’-elica singola. Il lobo C-terminale, più grande, è per lo più -elicoidale
con tre corti -filamenti: uno alla regione principale e due tra il “loop” di
attivazione e il corpo centrale del lobo C16.
È stato precedentemente notato come diverse strutture estese delle forme
attiva e inattiva delle tirosin-chinasi hanno avvalorato l’ipotesi del modello di
regolazione chinasica secondo cui il loop di attivazione subisce sostanziali
cambiamenti conformazionali in conseguenza alla fosforilazione.
Il dominio chinasico della Syk cristallizzata assume una forma a “loopout” simile a quella osservata nelle protein-chinasi fosforilate, attive. È definita
“loop-out” la conformazione in cui la fosforilazione stabilizza una forma aperta
dell’anello di attivazione, il quale non occupa il sito di legame del substrato ed è
compatibile con la catalisi. Sono state sovrapposte le strutture di Syk e della
chinasi Lck, dove Tyr526 di Syk si allinea con la fosfoTyr394 di Lck16(Fig. 20).
Fig. 20: Struttura di Syk (giallo)
sovrapposta con la Lck chinasi
(turchese) (codice PDB: 1XBB).
22
L’orientazione dei domini N- e C-terminali di Syk è la stessa di quella
visibile in Lck. Nella struttura di Syk i siti di legame per il substrato (ATP o
peptide che sia) sono accessibili; la regione DFG alla base N-terminale del loop
di attivazione si sistema in una conformazione a “DFG-in”, con Asp512
posizionato in maniera adeguata per la catalisi. Analogamente, Asp494 del loop
catalitico ed il resto della struttura catalitica sono organizzati come nella Lck,
più attiva, inclusa la parte del ponte salino conservato Glu-Lys16.
La forma “loop-out/DFG-in” maggiormente attiva di Syk preclude al
Gleevec la possibilità di legarsi nella trans-conformazione osservata per il
legame alla forma inattiva di Abl, poiché nella forma DFG-in la fenilalanina
occlude un sito di legame secondario per STI-57116(Fig. 21).
Fig. 21: Rappresentazione schematica
dei diversi modi con cui il Gleevec
lega la tirosin-chinasi.
Il dominio chinasico di Syk rimane invariato nel complesso Syk-Gleevec,
ma la molecola di STI-571 adotta una nuova cis-conformazione, piuttosto
diversa dalla trans-conformazione assunta nel legame alla forma inattiva di Abl.
In questo secondo caso, l’anello piperazinico si estende nella tasca di legame
secondaria, resa accessibile dalla fenilalanina del DFG, che si posiziona
esternamente. Nella cis-conformazione l’orientazione fa in modo che l’anello
aromatico del Gleevec formi una struttura ad U nel sito attivo con l’anello
piperazinico esteso all’esterno verso il solvente16.
Si può, a questo punto senza dubbio, affermare che il Gleevec rappresenta
un’efficace inibitore dell’Abl non fosforilata, mentre risulta essere relativamente
inefficace nei confronti della forma fosforilata, più attiva di Abl.
23
È stata recentemente cristallizzata una struttura ad alta risoluzione (2.0 A)
del dominio catalitico di una forma mutante della tirosin-chinasi Abl (H396P),
che risulta resistente all’inibitore STI-571. La struttura è osservata in complesso
con la piccola molecola dell’inibitore VX-680, il quale blocca l’attività di
diverse forme mutanti resistenti all’imatinib, inclusa una (T315I) che si
manifesta resistente sia all’Imatinib che a BMS-354825 (dasatinib), un inibitore
Src/Abl che sembra essere clinicamente efficace contro tutte le altre forme di
Bcr-Abl resistenti all’imatinib. Si è notato che VX-680 ha una notevole attività
inibitoria contro Bcr-Abl che presenta la mutazione T315I. Il dominio chinasico
di Abl legato a VX-680 non è fosforilato sul loop di attivazione nel cristallo, ma
è tuttavia in una conformazione attiva, non osservata per Abl. L’assunzione di
tale conformazione sembra essere il risultato di una sinergia tra la mutazione
His396Pro, che destabilizza la conformazione inattiva richiesta per il legame
dell’imatinib, e il legame di VX-680, che favorisce la conformazione attiva
attraverso legami a idrogeno ed effetti sterici. VX-680 è legato all’Abl in un
modo che accomoda la sostituzione dell’isoleucina per la treonina al residuo 315
(la posizione del “gatekeeper”). Il fatto che la cavità interna del dominio
chinasico di Abl non sia occupata da VX-680 e il riconoscimento specifico della
conformazione attiva spiegano l’efficacia di questo composto contro le forme
mutanti di Bcr-Abl, incluse quelle con mutazioni alla posizione del
“gatekeeper”17.
La struttura del dominio chinasico di Abl H396P è molto simile a quella
del dominio chinasico della Lck chinasi della famiglia Src nella conformazione
attiva18. La conformazione del loop di attivazione non fosforilato, a partire
dall’aspartato della regione DFG altamente conservata (Asp381) e continuando
con Trp405, è essenzialmente sovrapponibile con il loop di attivazione fosforilato
trovato nella struttura di Lck. Anche l’orientazione del lobo N-terminale
relativamente al lobo C-terminale è molto simile nelle strutture di Abl H396P e
di Lck attivo17 (Fig. 22A).
24
Fig. 22: Confronto del complesso VX-680 con altre strutture. (A) Sovrapposizione del complesso
Abl:VX-680 (blu) con la struttura della forma attiva del dominio chinasico di Lck (arancio; codice
PDB: 3LCK). (B) Conformazione del loop di attivazione di Abl:VX-680 (blu) sovrapposta alla struttura
del loop di attivazione attivo e fosforilato di Lck (arancio). (C) Conformazione dell’anello DFG nel
complesso VX-680 (blu) paragonata con la forma attiva di Lck (arancio), con la conformazione inattiva
in complesso con PD173955 (rosso; codice PDB: 1M52), e con la conformazione inattiva in complesso
con l’imatinib (blu scuro; codice PDB: 1OPJ).
La maggiore differenza tra la struttura di Abl H396P e quella dell’Abl
wild-type legate al Gleevec sta nella conformazione del loop di attivazione (Fig.
22C). L’aggiunta di un gruppo fosfato sulla Tyr393 dovrebbe stabilizzare la
forma attiva dell’anello di attivazione. Due catene laterali cariche positivamente,
Arg362 e Arg384, sono posizionate in modo da formare dei ponti salini con un
gruppo fosfato sulla Tyr393(Fig. 22B). Nell’Abl wild-type una terza catena
laterale basica che potrebbe potenzialmente coordinare il gruppo fosfato basico
viene fornita dall’His396(in figura mutata con la prolina). Questo tipo di
25
interazioni sembrano essere importanti nella proteina intatta per svolgere le
azioni inibitorie dei domini regolatori.
Il dominio chinasico di Abl wild-type è stato cristallizzato anche con
l’inibitore PD173955 (codice PDB: 1M52); la struttura del complesso mostra
che il loop di attivazione si trova in una conformazione estesa, la quale somiglia
alla forma attiva, esclusa la regione DFG e diversi residui (da Asp381 a Gly390)
sono leggermente sfasati rispetto a quanto è visibile nella conformazione
attiva17(Fig. 22C).
VX-680 è una molecola a forma di Y, con un gruppo N-metil-piperazinico
che costituisce la base o la “gamba” della Y, un gruppo pirimidinico alla
biforcazione, un gruppo metilpirazolico che rappresenta un braccio ed un gruppo
fenilico sostituito che va a formare l’altro braccio (Fig. 23).
Fig. 23: Struttura del complesso di VX-680. (A) Struttura del dominio chinasico di Abl mutato
(H396P) legato all’imatinib (a sinistra; codice PDB: 1OPJ) e a VX-680 (a destra). (B) Struttura
chimica di VX-680 e della regione DFG. (C) Legame del VX-680 al dominio chinasico di Abl.
Legami ad idrogeno nella regione cardine (linee tratteggiate in giallo) e legame ad idrogeno
con l’Asp381 (linee tratteggiate in rosso).
26
VX-680 è legato al sito di legame per l’ATP del dominio chinasico di
Abl, con il gruppo N-metil-piperazinico che si estende all’esterno del dominio
chinasico. La molecola si trova ancorata alla regione catalitica mediante quattro
legami ad idrogeno (Fig. 23C). Tre di questi si formano tra due gruppi
carbonilici (Glu316 e Met318) ed un azoto ammidico (Met318) nella così chiamata
“regione cardine” della chinasi, e tre atomi di azoto, uno nel linker tra il gruppo
pirimidinico e il gruppo metilpirazolico, e gli altri due nel gruppo
metilpirazolico. Il quarto legame ad idrogeno si trova tra l’azoto del gruppo
ammidico che lega il gruppo fenilico al sostituente ciclopropilico e la catena
laterale dell’aspartato della regione DFG (Asp381). Quest’ultimo legame a
idrogeno probabilmente rappresenta l’interazione che rafforza la conformazione
attiva del dominio chinasico quando si lega a VX-68017.
L’imatinib forma solo un legame ad idrogeno diretto con la regione
cardine, sull’azoto ammidico della struttura principale di Met 318. Un secondo
legame ad idrogeno tra l’imatinib e la regione cardine avviene sull’ossigeno
della catena laterale della Thr315, nella posizione del “gatekeeper”, rendendo
STI-571 molto sensibile alle sostituzioni in tale regione. Un’altra differenza
chiave tra l’imatinib e VX-680 è rappresentata dal fatto che quest’ultimo si
ancora fermamente alla regione cardine e coinvolge Asp381, ma non penetra così
in profondità nel dominio chinasico come fa l’imatinib. Il gruppo N-metilpiperazinico di VX-680, dunque, si trova esposto al solvente, mentre la
corrispondente regione di STI-571 nella struttura del complesso risulta visibile
dall’esterno (Fig. 24A).
C’è una caratteristica che STI-571 e VX-680 condividono nel modo di
legarsi al dominio chinasico. Il “P-loop” per il legame col fosfato della chinasi
(residui Lys247-Val256) viene distorto dalla conformazione a -forcina che esso
adotta quando si lega l’ATP e, invece, si attorciglia all’interno del sito attivo
della chinasi, cosicchè la catena laterale della Tyr253 vada a formare interazioni
idrofobiche con l’inibitore in entrambi i casi17(Fig. 24B). Il P-loop rappresenta
27
il sito di diverse mutazioni che conferiscono resistenza per il legame
dell’imatinib19.
Fig. 24: Modi di legarsi di VX-680. (A) Strutture a sfere di VX-680 (a sinistra) e dell’imatinib
(a destra). (B) Interazioni tra il loop di legame del fosfato (P-loop) del dominio chinasico e il
VX-680. (C) Residui che interagiscono con VX-680 mediante la catena laterale.
Grazie alla presenza dei quattro legami ad idrogeno che ancorano VX-680
alla regione catalitica, l’inibitore ha la possibilità di venire in contatto con 14
catene laterali all’interno del dominio chinasico (Fig. 24C). Tutte queste, ad
eccezione della Met290, appartengono alla regione centrale del dominio chinasico
coinvolto nel legame con l’ATP e nella catalisi e sono altamente conservate
nelle varie protein-chinasi17.
Le proteine recettoriali tirosin-chinasi (RPTKs) hanno la possibilità di
assumere diverse conformazioni distinte durante la loro attivazione e sono
capaci di tenere a freno l’attività della chinasi nativa anche in assenza di un
segnale di fosforilazione attivante8.
28
Fig. 25: Esempio di autofosforilazione
Molto spesso a livello dei siti di legame per l’ATP delle chinasi,
idrofobici, si è verificata l’autoinibizione soltanto grazie alla presenza di sistemi
di anelli planari coniugati, che sono sinonimo di poca solubilità e penetrazione
cellulare. Ad esempio, a causa della sua potenza contro un largo numero di
bersagli chinasici, l’ingombrante inibitore della chinasi staurosporina e i suoi
analoghi non hanno rappresentato adeguati punti di partenza per l’ideazione di
inibitori selettivi con buone proprietà fisico-chimiche (Fig. 26). Gli inibitori
della PDK-1 bisindolil-maleimide, al contrario, sembrano essere promettenti
nello studio e nella scoperta di inibitori utili dal punto di vista terapeutico8.
Fig. 26: Legame della staurosporina al
sito di legame per l’ATP all’interno delle
chinasi Tec. La Phe435 (in giallo) funge
da “gate” per la tasca idrofobica.
29
La famiglia III delle RPTKs è implicata in diversi tumori umani altamente
maligni. È caratterizzata da domini dell’immunoglobulina extracellulari, una
singola elica transmembrana, un dominio citoplasmatico catalitico ad attività
chinasica e da un dominio juxtamembranario autoinibitorio (JM), il quale
rappresenta la regione di maggiore interesse fra tutte. I complessi RPTK sono
responsabili dei segnali di trasmissione extracellulare all’interno delle cellule.
Nelle cellule normali il dominio autoinibitorio JM regola con specificità
l’attività delle tirosin-chinasi c-KIT e FLT-3 (Fig. 27).
Fig. 27: Organizzazione genica e sequenza del dominio juxtamembranario conservato del
recettore RPTK di tipo III. (A) Schema dell’organizzazione del gene/proteina RPTK di
tipo III: sono mostrati i 5 domini dell’immunoglobulina extracellulari, la membrana
cellulare, la regione juxtamembranaria (JM), i domini intracellulari della chinasi c-Kit
separati dal dominio di inserzione chinasico (KID). (B) Allineamento della sequenza delle
regioni JM di RPTKs di tipo III.
L’intricata base strutturale su come la regione JM è capace di funzionare
sia come dominio autoinibitorio (nello stato nativo non fosforilato) sia come un
dominio di segnale intracellulare fosforilato è stata recentemente rivelata per cKIT (codici PDB: 1T45 e 1T46) e per FLT-3 (codice PDB: 1RJB)8. c-KIT non
fosforilato è stato cristallizzato in presenza ed in assenza dell’inibitore STI-571
legato20.
In assenza di fosforilazione, si è visto che il dominio JM di c-KIT agisce
come una regione autoinibitoria, poiché si inserisce direttamente nella fessura
tra i gruppi N- e C-terminali, svolgendo, quindi, la C-elica e proteggendo anche
il gruppo DFG catalitico dal raggiungimento di una conformazione produttiva.
La fosforilazione del dominio JM di c-KIT, invece, evoca una drastica
variazione conformazionale che scopre il sito di legame per l’ATP. Il confronto
30
tra le strutture non fosforilata e fosforilata rivela che questi ampi cambiamenti
conformazionali rimuovono il dominio JM e la regione di attivazione dal sito
attivo catalitico, permettendo alla C-elica di entrare e di orientare correttamente
gli importanti residui catalitici DFG. Questo è stato proposto come base
molecolare per cui la trans-fosforilazione dal dominio JM nelle due regioni
chinasiche di c-KIT adiacenti sulla ligazione dei domini del recettore
dell’immunoglobulina conduce il segnale nel pathway.
L’inibitore STI-571, inoltre, è capace di stabilizzare una conformazione di
c-KIT che fa da intermedio per le forme attivata e autoinibita. STI-571 sfrutta
molte delle stesse interazioni idrofobiche con c-KIT che stabilizzano l’inusuale
conformazione autoinibita del sito di attivazione, ma scaccia il dominio JM dal
sito
attivo.
STI-571,
dunque,
occupa
una
piccola
tasca
idrofobica
precedentemente occupata dal residuo Trp577 di JM8(Fig. 28).
Fig. 28: Basi strutturali dell’autoinibizione e della inibizione di STI-571 nei confronti di cKit. (a) Senza la fosforilazione, il dominio JM di autoinibizione (rosso) di c-Kit è
stabilizzato dalle ampie interazioni idrofobiche con il loop ricco di glicina ed una
conformazione inattiva del loop di attivazione. (b) STI-571 si lega ad una forma autoinibita
di c-Kit in cui l’anello di attivazione e l’elica C rimangono chiusi. Il dominio JM è liberato
dal sito attivo e STI-571 si inserisce in una tasca idrofobica che era precedentemente
occupata dal residuo Trp577.
La struttura cristallizzata della forma autoinibita, non fosforilata di FLT321 fornisce una visione chiara del meccanismo mediante cui il dominio JM
esercita il suo effetto autoinibitorio e anche il ruolo mutazioni di duplicazione
sequenziale interna (ITDs; avvengono per lo più a livello di JM o nel loop di
attivazione) nell’attivazione costitutiva di FLT-3 nei pazienti con leucemia
31
mieloide acuta (AML)21. Nell’AML, le mutazioni puntiformi dei residui
localizzati nel sito di attivazione danno luogo a forme attivate costitutivamente
di FLT-3. Il sito di attivazione di FLT-3 è simile a quello osservato in altre
chinasi inattive, inclusa c-KIT. È stato spiegato dettagliatamente come la
conformazione inattiva di FLT-3 sia correlata alle mutazioni conosciute che
generano una chinasi attiva costitutivamente anche senza il legame con il
recettore. Si è spiegato un meccanismo in cui un segmento peptidico funge da
linker nella regione JM ed è capace di allineare correttamente e mantenere il
gruppo di scambio di JM nella propria posizione sia durante che dopo la
transizione tra lo stato attivato e quello inattivato di FLT-38.
Un importante inibitore selettivo del processo di autoinibizione di FLT-3
è il CEP-701, il quale si è dimostrato estremamente efficace nella terapia della
leucemia mieloide acuta (AML) 22.
Una conseguenza di questo tipo di passaggio allo stato inattivo è
rappresentata dal fatto che un involucro altamente polare creato dai residui
coinvolti nel legame col metallo e nella catalisi in c-KIT viene sostitutito da una
tasca idrofobica. Quest’ultima è stata prima sfruttata per il legame di STI-571
alla proteina inattiva c-Abl e viene anche usata dallo stesso inibitore per legare
c-KIT. Il successo di STI-571 (Gleevec) mette in risalto come queste strutture
possono dar vita a nuove opportunità per la creazione di inibitori potenti e
selettivi per questi enzimi RPTK inattivi8.
32
3. CONCLUSIONI
Le tirosin-chinasi racchiudono un numero molto vasto di proteine ad
attività enzimatica, di tipo recettoriale e non-recettoriale. Esse, catalizzando la
fosforilazione di varie molecole substrato, svolgono un ruolo fondamentale nella
trasmissione del segnale e nell’induzione della risposta cellulare. Un’aumentata
attività delle tirosin-chinasi può portare alla persistente stimolazione delle vie di
trasduzione del segnale con importanti conseguenze in diversi processi biologici,
in particolare nella crescita e nella differenziazione cellulare, nell’apoptosi e
nella cancerogenesi. Per questo motivo le tirosine chinasi e i meccanismi di
trasduzione del segnale possono essere identificati come potenziali bersagli per
la progettazione di nuovi farmaci.
Avendo le chinasi una loro specificità di substrato, il numero di proteine
bersaglio della loro azione sembra essere relativamente limitato; è, invece,
enorme la quantità di funzioni fisiologiche che esse vanno a svolgere una volta
attive. Le tirosin-chinasi, pertanto, sono in continua fase di studio, in modo tale
da poter essere ben caratterizzate. Questo rende possibile la progettazione di
sistemi adatti a bloccare la loro attività incontrollata per poter ripristinare la
normalità all’interno delle cellule trasformate e, in particolar modo, recuperare il
processo apoptotico delle cellule tumorali.
Il Gleevec (Imatinib, STI-571) ha rappresentato il primo inibitore delle
tirosin-chinasi ad essere immesso in commercio per la sua efficacia contro la
leucemia mieloide cronica. Il suo successo ha lasciato ipotizzare che simili
risultati potessero essere raggiunti nel trattamento di altri tumori, agendo a
livello molecolare su proteine aberranti espresse dalla neoplasia. In realtà la
complessità delle alterazioni molecolari inerente i tumori ha, in alcuni casi,
deluso le aspettative, ma ha sicuramente offerto una nuova via di attacco e
sviluppo farmacologico da associare alla chemioterapia o da utilizzare come
trattamento alternativo ai farmaci citotossici.
33
Bibliografia
1. farmacia.unical.it/cartelle/download/dispense/sisci/
2. Davies DE, Chamberlin SG
Targeting the Epidermal Growth Factor
Receptor for therapy of carcinomas. Biochemical Pharmacology 1996 51:
1101-1110
3. Pontieri GM, Patologia generale, Ed. Piccin, 1987
4. Thomas S, Brugge J Cellular functions regulated by Src family kinase.
Annual Reviews of Cell and Developmental Biology 1997 13: 513-609
5. Nagar B, Bornmann WG, Pellicena P, Schindler T, Veach DR, Miller WT,
Clarkson B, Kuriyan J Crystal structures of the kinase domain of c-Abl in
complex with the small molecule inhibitors PD173955 and Imatinib (STI571). Cancer Research 2002 62: 4236-4243
6. Oliff A, Gibbs JB, McCormick F Nuovi bersagli molecolari per la terapia
dei tumori. Le Scienze 1996 339: 106-111
7. Cognetti F, Felici A Le nuove terapie mirate. Ministero della Salute - Area
editoriale
8. Cheetman GMT
Novel protein kinases and molecular mechanism of
autoinhibition. Current Opinion in Structural Biology 2004 14: 700-705
9. Schindler T, Bornmann WG, Pellicena P, Miller WT, Clarkson B, Kuriyan J
Structural mechanism for STI-571 inhibition of Abelson tyrosine kinase.
Science 2000 289: 1938-1942
10.Zimmermann J, Buchdunger E, Mett H, Meyer T, Lydon NB (Senza titolo)
Bioorg. Med. Chem. Lett. 1997 7: 187-192
11.Druker BJ et al. Activity of a specific inhibitor of the Bcr-Abl tyrosine
kinase in the blast crisis of chronic myeloid leukemia and acute
lymphoblastic leukemia with the Philadelphia chromosome. NEJM 2001 344:
1038
34
12.Apperley JF, Gardembas M, Melo JV, et al. Response to imatinib mesylate
in patients with chronic myeloproliferative diseases with rearrangements of
the platelet-derived growth factor receptor . N. Engl. J. Med. 2002 347:
481-487
13.Demetri G
Targeting the molecular pathophysiology of gastrointestinal
stromal tumors with imatinib. Mechanisms, successes, and challenges to
rational drug development. Hematol. Oncol. Clin. North. Am. 2002 16: 11151124
14.Dorsey JF, Jove R, Kraker AJ, Wu J The pyrido(2,3-d)pyrimidine derivate
PD180970 inhibits p210Bcr-Abl tyrosine kinase and induces apoptosis of
K562 leukemic cells. Cancer Res. 2000 60: 3127-3131
15.Moasser MM, Srethaphakdi M, Sachar KS, Kraker AJ, Rosen N Inhibition
of Src kinases by a selective tyrosine kinase inhibitor causes mitotic arrest.
Cancer Res. 1999 59: 6145-6152
16.Atwell S, Adams JM, Badger J, Buchanan MD, Feil IK, et al. A novel mode
of Gleevec binding is revealed by the structure of spleen tyrosine kinase. J.
Biol. Chem. 2004 279: 55827-55832
17.Young MA, Shah NP, Chao LH, Seeliger M, Milanov ZV, Biggs WH,
Treiber DK, Patel HK, Zarrinkar PP, Lockhart DJ, Sawyers CL, Kuriyan J
Structure of the kinase domain of an imatinib-resistant Abl mutant in
complex with the Aurora kinase inhibitor VX-680. Cancer Res. 2006 66:
1007-1014
18.Yamaguchi H, Hendrickson WA Structural basis for activation of the human
lymphocyte kinase Lck upon tyrosine phosphorylation. Nature 1996 384:
484-489
19.Shah NP, Nicoll JM, Nagar B, et al.
Multiple Bcr-Abl kinase domain
mutations confer polyclonal resistance to the tyrosine kinase inhibitor
imatinib (STI-571) in chronic phase and blast crisis chronic myeloid
leukemia. Cancer Cell 2002 2: 117-125
35
20.Mol CD, Lim KB, Sridhar V, Zou H, Chien EY, Sang BC, Nowakowski J,
Kassel DB, Cronin CN, McRee DE Structure of a c-kit product complex
reveals the basis for the kinase transactivation. J. Biol. Chem. 2003 278:
31461-31464
21.Griffith J, Black J, Faerman C, Swenson L, Wynn M, Lu F, Lippke J, Saxena
K The structural basis for autoinhibition of FLT3 by the juxtamembrane
domain. Mol. Cell. 2004 13: 169-178
22.Smith BD, Levis M, Beran M, Giles F, Kantarjian H, Berg K, Murphy KM,
Dauses T, Allebach J, Small D
Single-agent CEP-701, a novel FLT3
inhibitor, shows biologic and clinical activity in patients with relapsed or
refractory acute myeloid leukemia. Blood 2004 103: 3669-3676
36