La linea laterale dei pesci
La linea laterale (LL) è un organo sensoriale presente nei vertebrati acquatici, principalmente
nei pesci, che consente di valutare il movimento dell'acqua. Il sistema della LL appare
esternamente come una linea che parte dalle branchie ed arriva sino alla coda (Fig. 1). La maggior
parte delle larve degli anfibi ed alcuni anfibi acquatici hanno un sistema meccano-sensitivo simile
alla LL. Le cellule recettrici sono cellule epiteliali ciliate, simile a quelle presenti nell'orecchio
interno dei mammiferi, dove valutano i suoni e le accelerazioni lineari ed angolari del capo. In
alcune specie le cellule recettrici della LL si sono modificate, divenendo elettrorecettori.
Linea laterale
Linea laterale
A
B
D
C
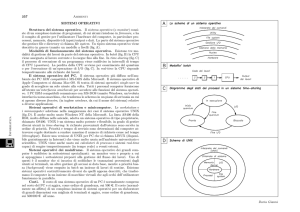
Fig. 1. La linea laterale in due specie di pesci (A e B). In Xenopus la LL assomiglia ad una cucitura
con ago e filo, "stitch" in Inglese (C e D).
Il neuromasto
L'unità funzionale della LL è il neuromasto (NM) (Fig. 2), costituito da un gruppo di cellule ciliate
e da una cupola che le racchiude. Le cellule ciliate sono cellule cilindriche, aventi sulla superficie
che guarda all'esterno (verso il fluido) numerose ciglia, circa 150, aventi lunghezza
progressivamente crescente man mano che si avvicinano al chinociglio, una ciglia nettamente più
lunga, disposta lateralmente. In ciascun neuromasto le ciglia sono orientate lungo due direzioni
opposte, che definiscono la direzione di maggiore sensibilità al movimento dell'acqua.
L'orientamento delle ciglia è differente nei vari neuromasti.
Le ciglia sono racchiuse da un materiale gelatinoso a forma di cupola, che si interpone tra
l'ambiente esterno e le ciglia. Il polo basale è in contatto sinaptico con una fibra afferente (la
cellula ciliata è l'elemento presinaptico) ed una efferente (la cellula ciliata è l'elemento
postsinaptico). Le cellule ciliate secernono glutammato, attivando la via afferente. La via efferente
secerne acetilcolina e sembra avere effetto inibitorio.
Cupola
Chinociglio
Ciglia
Cellula di
supporto
Cellule
ciliate
Fibre
nervose
Fig. 2. Struttura del neuromasto ( sinistra) e microfotografia di una cellula ciliata (destra)
I neuromasti possono essere localizzati superficialmente. Alternativamente, sulla superficie del
pesce sono presenti dei pori che sfociano in un canale in cui sono posizionati i neuromasti (Figg.
3 e 4).
Sistema di canali nel capo
Sistema di canali del tronco
Canale della linea
laterale
Flusso di acqua Apertura Epidermide
Vie nervose
Neuromasti
Fig. 3.
Struttura della
linea laterale.
Fig. 4. La linea laterale dei pesci. I neuromasti superficiali sono indicati da punti, mentre i pori
dei neuromasti annidati nei canali sono indicati da cerchi. In molte specie di pesci, questi sono
disposti sopra e sotto l'occhio e sulla mandibola inferiore.
Alcune specie di pesci hanno meno di 50 NM superficiali, mentre altre arrivano sino a parecchie
migliaia. I canali della testa dei pesci hanno ramificazioni estremamente complesse, il cui
significato funzionale è ignoto (Fig. 5).
Fig. 5. Ramificazioni dei canali della
linea laterale nella testa dei pesci.
Che cosa stimola i neuromasti?
Un'usuale sorgente di movimento dell'acqua è il movimento degli altri animali (Fig. 6). Il nuoto di
Lepomis gibbosus, avente dimensione di circa 20 cm genera complessi movimenti dell'acqua, che si
propagano per più di 200 mm (Fig. 7).
Fig. 6. Il nuoto di un pesce genera una scia composta da vortici e dei flussi d'acqua orientati nella
direzione del movimento.
mm/s
5
4
3
2
1
A
B
C
100 mm
Fig. 7. A e B due esempi di movimenti generati da Lepomis gibbosus. Gli pseudocolori indicano la
velocità dell'acqua, la barra la distanza a cui si propagano i movimenti; C, Lepomis gibbosus.
Sebbene i neuromasti superficiali e quelli disposti nei canali abbiano proprietà strutturali e
funzionali simili, la diversa localizzazione contribuisce a modulare la risposta agli stimoli. I
neuromasti superficiali sono investiti da stimoli nelle tre dimensioni, probabilmente di intensità
maggiore rispetto ai neuromasti profondi e la loro risposta è proporzionale alla velocità del fluido. I
neuromasti nei canali sono invece stimolati da un flusso che scorre lungo l'asse del canale, a sua
volta generato dal flusso esterno. Di fatto, i neuromasti del canale sembra rispondano non alla
velocità del liquido circostante ma alla sua accelerazione.
I neuromasti sono in grado di percepire i movimenti d'acqua generati anche da organismi molto
piccoli, appartenenti, per esempio, allo zooplankton. In Fig. 8 sono riportati gli spostamenti
dell'acqua (asse y, non indicato) generati da organismi con dimensioni dell'ordine del mm.
Fig. 8. Due piccoli organismi generano spostamenti del liquido di diverso andamento
temporale.
I movimenti dei liquidi si smorzano all'incirca con il cubo della distanza.
E' opportuno rilevare come gran parte degli esperimenti venga effettuata in un ambiente di
acque ferme, cosicchè raramente viene valutare l'effetto stimolatoriodei movimenti autonomi
dell'acqua, quali quelli generati dalle correnti e dal movimento delle onde e del loro rifrangersi.
La linea laterale ed il comportamento dei pesci
La LL consente di percepire i movimenti d'acqua generati dalla preda, della quale si valuta la
posizione, la dimensione e forse lo stile di nuoto; nel complesso, le informazioni consentono di
orientare il movimento verso la preda o di allontanarsi dal possibile predatore e potrebbero
consentire il riconoscimento del tipo di pesce che produce il movimento dell'acqua, favorendo, per
esempio, l'accoppiamento. Gli insetti che si muovono freneticamente perchè intrappolati nel pelo
dell'acqua generano movimenti ad alta frequenza che pilotano i pesci predatori. I pesci ciechi che
vivono nelle grotte usano i movimenti dell'acqua generati dal loro muoversi per valutare, tramite la
propria linea laterale, la presenza di oggetti nelle vicinanze, che modificano i movimenti d'acqua
generati dal pesce. Possono attraversare una barriera costituita da una serie di bastoni immersi
nell'acqua senza toccarli. La meccano-recezione è inoltre essenziale nel pesce per la formazione dei
banchi (schooling). Si è visto che, dopo essere riusciti a formare un banco di pesci in un contenitore,
un singolo pesce rimosso e poi immesso nuovamente nel contenitore si integrava nuovamente nel
banco. Anche i pesci bendati erano capaci di reintegrarsi nel banco, mentre quelli con la LL non
funzionante erano incapaci: la LL era necessaria per pilotare questo comportamento.
La trasduzione del segnale
La struttura della cellula ciliata è mostrata in Fig. 9. Il corpo della cellula è inglobato nelle
cellule di supporto, che formano tight junction sia con la cellula ciliata che tra di loro. Nel polo
apicale troviamo le ciglia, nel polo basale le terminazioni nervose afferenti ed efferenti.
Ciglia
Chinociglio
Microvilli
Tj
Tj
Nucleo
Vescicole
sinaptiche
Fibra
efferente
Fig. 9. Struttura della
cellula ciliata. I due cerchi
tratteggiati individuano le
tight junction (Tj).
Fibra
afferente
Il piegamento del chinociglio lontano dalle ciglia apre canali cationici meccano-sensibili e tende
a depolarizzare la cellula. Il movimento in direzione opposta tende ad inibire l'apertura dei canali
meccano-sensibili, iperpolarizzando la cellula. La depolarizzazione aumenta la liberazione di
mediatore chimico, aumentando la frequenza di scarica delle fibre afferenti. L'iperpolarizzazione
ha un effetto opposto. Come già evidenziato, in ciascun neuromasto le ciglia sono orientate lungo
due direzioni opposte, che definiscono la direzione di maggiore sensibilità al movimento
dell'acqua. Una fibra nervosa afferente è attivata da neuromasti le cui ciglia hanno il medesimo
orientamento. Uno spostamento della cupola che allontana il chinociglio dalle ciglia in più cellule,
le depolarizza ed aumenta la frequenza di scarica della fibra afferente ad esse collegata. Le ciglia
con orientamento opposto sono tutte iperpolarizzate e diminuiscono la frequenza di scarica della
fibra afferente ad esse collegata secondo la linea di orientamento aumenta la scarica afferente (Fig.
10). In altre parole, una fibra nervosa afferente non è collegate a cellule ciliate che, in seguito ad
un movimento della cupola, in parte si iperpolarizzano ed in parte si depolarizzano.
A
B
Nervo
Fibra nervosa
Fig. 10. Uno spostamento della cupola nella direzione ciglia corte-chinociglio depolarizza la
cellula e provoca un aumento della frequenza di scarica nella fibra afferente connessa con la
cellula A. Nella cellula B l'orientamento delle ciglia è ruotato di 180 ° ed avremo
un'iperpolarizzazione della cellula ed una diminuzione della frequenza di scarica nella fibra
afferente connessa con la cellula B.
La codificazione del segnale
La scarica afferente globale del nervo afferente non varia invertendo la direzione del flusso. Di
fatto, stimolando i neuromasti lungo una direzione, alcune fibre aumenteranno la frequenza di
scarica, altre la diminuiranno (vedi Fig. 10). Invertendo la direzione, le fibre prima inibite
aumenteranno la frequenza di scarica, e viceversa, cosicchè la frequenza globale non è sensibile
alla direzione del flusso (Fig. 11).
Fig. 11. Frequenza di scarica del nervo in funzione della velocità (ampiezza e direzione).
Le singole fibre proiettano però in modo specifico ai neuroni del mesencefalo, che rispondono
specificamente in funzione della direzione del movimento (Fig. 12).
Fig. 12. Il neurone in esame aumenta o diminuisce la frequenza di scarica in funzione della
direzione del movimento.
L'organizzazione strutturale della linea laterale suggerisce che vi siano almeno quattro gruppi di
informazioni relativi al movimento dell'acqua che, in parallelo, arrivano ai centri nervosi:
informazioni provenienti dai neuromasti superficiali, informazioni proveniente dai neuromasti
disposti nei canali, e, all'interno di questi due gruppi, informazioni relative all'orientamento delle
ciglia dei neuromasti rispetto al movimento dell'acqua.
In analogia con il comportamento delle cellule ciliate dell'organo uditivo, anche i neuromasti
hanno una sensibilità preferenziale alla frequenza dello stimolo. Alcuni neuromasti rispondono a
frequenze tra i 30–200 Hz, altri a frequenze inferiori a 30 Hz.
Come già riportato, l'innervazione efferente colinergica è inibitoria; l'inibizione sembra essere
attivata in sincronia con il movimento del pesce. Sembrerebbe avere la funzione di inibire
l'attivazione dei neuromasti a causa del proprio movimento, facilitando il riconoscimento dei
flussi d'acqua generati dagli altri animali.