PROGRAMMA ESAMI IDONEITA’
SCIENZE
(Chimica – Biologia)
indice
LA CELLULA
LA RIPRODUZIONE DELLE CELLULE (MITOSI, MEIOSI)
GREGOR MENDEL E LE SUE LEGGI
LA SCOPERTA DELLE PARTICELLE SUBATOMICHE E I MODELLI ATOMICI
LA TAVOLA PERIODICA
LE REAZIONI CHIMICHE
IL METABOLISMO DEL CORPO UMANO: LA DIGESTIONE
LA TASSONOMIA
CHARLES DARWIN E LE TEORIE EVOLUTIVE
I FOSSILI
IL CARBON FOSSILE
GLI OGM
L’ACCIAIO
I METALLI
La cellula
Le cellule sono le unità fondamentali della vita; le più piccole entità definite vive. La
cellula possiede la capacità di mantenere al proprio interno condizioni chimicofisiche diverse dall'ambiente circostante: di fatti è un vero piccolo ma potentissimo
laboratorio chimico all'interno del quale si susseguono continuamente reazioni che
stanno alla base del metabolismo vitale, è perciò necessario che l'ambiente all'interno
del quale si svolgono tali reazioni mantenga tutte le condizioni chimiche e fisiche
indispensabil per una corretta attività. Tra le attività metaboliche più comuni
troviamo: l'acquisizione di energia e l'utilizzo di questa per mantenere le condizioni
chimiche ottimali all'interno della cellula e la sintetesi dimolecole organiche
necessarie per la crescita e per la riproduzione. Gli esseri viventi possono essere
costituiti da una singola cellula come nel caso dei batteri, lieviti o alghe nel regno
vegetale, oppure costituiti da più cellule; o meglio detti multicellulari.
Gli organismi multicellulari sono il risultato dell'evoluzione in riposta a difficoltà
derivate dai limiti di crescita a cui una singola unità vitale può andare incontro in
quanto l'eccessivo volume interno comporta un incremento delle distanze tra la
membrana ed il nucleo, nonché una peggiore utilizzazione dei metaboliti interni.
Gli organismi multicellulari risultano quindi costituiti da numerose cellule originatesi
da continue divisioni partite da una singola cellula di origine. In un individuo
multicellulari tutte le cellule devono compiere le ordinarie operazioni di
mantenimento ed inoltre ogni cellula possiede delle diverse competenze che
apportano speciali contributi al corpo, ad esempio le cellule muscolari sono
specializzate nella contrazione, le cellule nervose sono particolarmente efficienti
nella produzione e nella ricezione di segnali chimici ed elettrici, in tal modo si viene
a formare una specializzazione delle attività che stanno alla base della formazione di
tessuti e organi differenti.
Le cellule posseggono tre porzioni principali:
Membrana plasmatica o membrana cellulare: riveste l'esterno della cellula e funziona
come un filtro controllando tutto ciò che entra e che esce dalla cellula, come vedremo
in seguito non è un semplice filtro passivo ma possiede la capacità di regolare
attivamente il passaggio di metaboliti sia in entrata che in uscita.
Citoplasma: costituito da acqua, Sali, molecole organiche ed enzimi metabolici. Il
citoplasma accoglie poi particolari componenti chiamati organuli all'interno dei quali
si svolgono specializzate operazioni metaboliche.
Organuli: sono molto numerosi e rappresentano degli elementi molto importanti per il
metabolismo
cellulare.
Tra
gli
organuli
di
nostro
interesse
troviamo:
- Ribosomi: sono il luogo in cui avviene la sintesi delle proteine, questi organuli
eseguono
in
prima
persona
le
direttive
contenute
nel
nucleo.
- Mitocondri: sono le centrali energetiche della cellula, all'interno dei quali si
svolgono i processi di respirazione: impiegando l'ossigeno degradano molecole
organiche per trarne energia.
Nucleo cellulare: al suo interno si trova il materiale genetico della cellula, DNA,
RNA. Il materiale genetico contiene tutte le istruzioni necessarie per la sintesi delle
proteine e di conseguenza il materiale genetico risulta essere la fonte di tutte le
informazioni necessarie alla cellula per guidare le proprie attività vitali.
La riproduzione delle cellule
La riproduzione, in biologia, è l'insieme dei meccanismi mediante i quali gli esseri
viventi provvedono alla conservazione della propria specie generando nuovi individui
simili a sé e che subentreranno al genitore, o ai genitori, nella popolazione.
Una cellula eucariota
può
dividersi
mantenendo
invariato
il
proprio
corredo cromosomico e generando due cellule figlie geneticamente uguali, oppure
può dimezzare il corredo cromosomico generando da una a quattro cellule figlie che
avranno un corredo cromosomico differente.
Il
primo
fenomeno,
noto
come mitosi,
coinvolge
sia
cellule aploidi sia
cellule diploidi e si identifica con la moltiplicazione cellulare. Lo scopo della
moltiplicazione cellulare è quello di far accrescere un organismo oppure di
incrementare una popolazione di organismi geneticamente uguali.
Il secondo fenomeno, noto come meiosi, coinvolge solo cellule diploidi e rappresenta
la prima fase di un fenomeno sessuale, in quanto si alterna con la cariogamia, evento
inverso attraverso il quale i corredi cromosomici di due cellule aploidi,
dette gameti si fondono per formare un'unica cellula diploide detta zigote, oppure
una meiospora che si sviluppa in un individuo aploide. Lo scopo biologico della
sessualità è quello di introdurre in una popolazione elementi di variabilità
genetica per mezzo delle mutazioni e della ricombinazione genica.
La divisione mitotica negli organismi unicellulari si identifica sempre con la
riproduzione, in quanto da una cellula individuo si originano due cellule individuo;
attraverso la divisione mitotica gli organismi unicellulari ottengono perciò sia la
perpetuazione della specie sia l'incremento numerico della popolazione. La sessualità
degli organismi unicellulari non contribuisce alla crescita della popolazione, in
quanto da due individui se ne forma solo uno. In questi casi la sessualità ha il fine di
perpetuare la specie nel tempo oppure di incrementare la variabilità genetica in una
popolazione.
Negli organismi pluricellulari la mitosi è fondamentalmente il meccanismo con cui si
svolge l'accrescimento del singolo individuo e, in alcuni microrganismi, anche la
riproduzione. L'alternanza fra meiosi e cariogamia comporta anche la riproduzione
(nel senso di moltiplicazione) in quanto un solo organismo pluricellulare produce più
gameti nel corso della sua vita.
Sulla base di questi elementi, si riconoscono due forme di riproduzione: una
presuppone la mitosi e viene detta riproduzione assessuata, l'altra presuppone
l'alternanza fra meiosi e cariogamia e viene detta riproduzione sessuata.
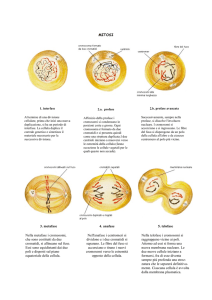
La mitòsi è la riproduzione per divisione equazionale della cellula eucariote. Il
termine viene spesso utilizzato anche per la riproduzione delle cellule procariote, un
processo molto più semplice e più correttamente chiamatoscissione binaria o amitosi.
Il termine mitosi deriva dal greco mìtos, "filo"; nome dovuto all'aspetto filiforme dei
cromosomi durante la metafase.
La mitosi, nell'uomo, riguarda le cellule somatiche dell'organismo (ossia tutte le
cellule fuorché quelle che hanno funzione riproduttiva: i gametociti primari, i quali
vanno incontro alla meiosi) e le cellule germinali ancora indifferenziate.
Il ciclo cellulare si suddivide in tre fasi: l'interfase, in cui la cellula si prepara alla
divisione; la mitosi, periodo di gran lunga più breve in cui la cellula si divide; il
periodo G2 - più o meno definitivo -, nel quale la cellula si specializza nella sua
funzione. In quest'ultimo periodo, la cellula non è in grado o non è stimolata
a riprodursi.
Il processo inizia con la condensazione della cromatina, che avviene grazie alla
presenza di proteine istoniche che fungono da centri primari di organizzazione del
riavvolgimento del DNA - primo ordine di spiralizzazione - e della topoisomerasi II,
che, oltre alla sua funzione catalitica, agisce come centro di organizzazione del
secondo ordine di spiralizzazione. Segue un terzo ordine di cui non si conoscono le
proteine implicate; forse è conseguenza della tensione accumulata dalle precedenti
spiralizzazioni. Questo grosso superfilamento viene prima impaccato formando delle
anse che poi si riuniscono formando il cromosoma visibile. La durata media di questo
meccanismo di riproduzione cellulare varia in media, negli organismi superiori, tra le
15 e le 30 ore.
Eventi precedenti la mitosi
Prima della mitosi avviene l'interfase, momento molto importante nella vita della
cellula. Difatti proprio in questa fase gli organelli della cellulaaumentano e ne
permettono la duplicazione. L'interfase si suddivide in tre sottofasi: la fase G1, in cui
la cellula si accresce; la fase S, nella quale la cellula replica il materiale nucleare e
il DNA; la fase G2, durante la quale la cellula si prepara a dividersi.
Profase
Durante la prima fase della mitosi, nel caso della specie umana, i cromosomi si
condensano e sono visibili anche al microscopio ottico sotto forma di doppi
bastoncelli basofili: i cromatidi gemelli (o fratelli). Questi sono agganciati tra loro in
un punto centrale, detto centromero, grazie ad un complesso sistema di interazioni tra
il DNA e numerose proteine chiamato fuso acromatico.
Viene sintetizzato un secondo centrosoma, ed entrambi appaiono circondati da una
coltre di microtubuli: è il fuso mitotico, formato da dimeri di sub-unità
proteiche tubulina alfa e beta. Il ciclo si potrebbe interrompere in questo punto se alla
coltura si aggiungesse la tossina falloidina, che impedisce la formazione dei filamenti
di microtubuli; ciò si fa quando si vogliono visualizzare al microscopio i cromosomi
per studiarne le caratteristiche. L'apparato del Golgi e il reticolo endoplasmatico in
questa fase si scompongono in piccole vescicolette che si distribuiscono
uniformemente in tutto il citoplasma; anche la membrana nucleare, grazie alla sua
doppia struttura, si scompone similmente ai suddetti organelli.
Metafase
Questa fase inizia attraverso una sub-fase, la prometafase, in cui avviene
l'improvvisa dissoluzione della membrana nucleare, che si frammenta in tante
vescicolette.
Tale
processo
viene
innescato
dalla fosforilazione,
attraverso
delle chinasi, dell proteine delle lamine (filamenti intermedi) che costituiscono la
lamina nucleare; in conseguenza della fosforilazione i filamenti si dissociano negli
elementi costitutivi.
I due centrosomi si portano ai poli opposti della cellula ed agiscono come centri di
organizzazione microtubulare, catalizzando l'allungamento ed assicurando il corretto
orientamento dei microtubuli che andranno a breve a legarsi al centromero di uno dei
due cromatidi gemelli. In questa fase si possono verificare degli errori e due
microtubuli si possono agganciare allo stesso cromatidio dando poi una cellula figlia
mutilata e non vitale.
Le ventitré coppie di cromatidi vengono portate nella parte mediana della cellula,
formando la piastra equatoriale, in cui un piano immaginario, passante per i
centromeri, divide le coppie di DNA. È questo il momento più favorevole per lo
studio dei cromosomi, che sono ora al massimo della loro spiralizzazione e affiancati
ordinatamente lungo la piastra equatoriale posta al centro della cellula.
Anafase
Durante l'anafase, i cromatidi migrano verso i due centrosomi ai poli opposti della
cellula. Si riconoscono due momenti, detti anafase A e anafase B. Nella prima si
assiste alla separazione dei due cromatidi fratelli ad opera di un enzima, detto
separasi, con relativa migrazione degli stessi grazie a proteine motore (tipo dineine
citoplasmatiche) presenti a livello del cinetocore. Nell'anafase B si assiste al
reciproco scorrimento dei microtubuli polari del fuso mitotico con conseguente
allontanamento dei due centrosomi verso direzioni opposte. Pertanto si ottiene il
ripristino, per ogni polo, del numero originario di cromosomi.
Telofase
Nell'ultima fase della mitosi, i cromosomi si despiralizzano. Intorno ai due nuovi
complessi cromosomici ricompaiono le membrane nucleari e gliorganelli si
ricompongono. La telofase si conclude con una sottofase: la citodieresi, in cui si
separa il citoplasma in modo equivalente in entrambe le nuove cellule. La cellula si
divide al centro formando due cellule figlie, esattamente identiche alla cellula madre
ma più piccole. Questo avviene grazie ad un anello di actina creatosi al centro della
cellula madre che, contraendosi, stringe la cellula al centro. A tal punto le proteine
specializzate operano la fusione e la separazione della membrana in punti specifici e
le due cellule si separano.
In alcune cellule la telofase non avviene e si accumulano all'interno di uno stesso
nucleo di una stessa cellula da due ad alcune decine di corredi cromosomici. Questo
tipo di cellule si chiama plasmodio. L'esempio principale sono i protozoi del
genere plasmodium come il P. malariae. Anche cellule umane vanno incontro a
questo processo o patologicamente, come le cellule tumorali, o fisiologicamente
come nelmegacariocita.
La meiosi è
un
processo
della
riproduzione sessuale mediante
il
quale
una cellula eucariotica con corredo cromosomico diploide (raccolto in un cariotipo)
dà origine a quattro cellule con corredo cromosomico aploide, ovvero che da una
cellula madre si formano quattro cellule figlie, tutte diverse fra loro. Potrebbe
sembrare molto simile alla mitosi ma, al contrario di questa, si ha la riduzione da
corredo in doppia copia a corredo a semplice copia, e tramite il cosiddetto crossingover (incrocio esterno), si ha lo scambio e la ricombinazione genetica.
Nella
riproduzione sessuale,
la
ricombinazione
dell'informazione
genetica
proveniente dalle cellule di due organismi differenti (padre e madre), produce risultati
ogni volta diversi, e naturalmente diversi anche dai due genitori.
Ogni genitore fornisce quindi un corredo cromosomico "semplice" aploide (detto
anche "dimezzato"), cellula uovo nella femmina e spermatozoo nel maschio; la
fusione (fecondazione), dei due corredi dimezzati (materno e paterno) e "rimescolati"
ricostituisce il corredo intero, e dà origine ad una singola nuova cellula,
detta zigote che diverrà il nuovo individuo.
Il cariotipo cromosomico è l'insieme del numero, della forma e delle dimensioni
dei cromosomi che costituiscono il corredo genetico completo dello zigote, e quindi
del nuovo individuo.
Ad una duplicazione del materiale genetico, che avviene nella fase pre-meiotica S,
corrispondono due divisioni nucleari:
1. Prima divisione meiotica o meiosi I (fase Riduzionale)
2. Seconda divisione meiotica o meiosi II (fase Equazionale)
Fase pre-meiotica S o interfase I
Avviene la duplicazione del materiale genetico: da ogni cromosoma risultano
due cromatidi fratelli identici, attaccati in corrispondenza deicentromeri.
La meiosi I si apre con la profase, un processo più lungo e complicato della profase
mitotica.
Si
suddivide
in
5
stadi:
leptotene, in cui il materiale genetico si condensa a formare strutture bastoncellari;
zigotene,
durante
il
quale
avviene
la
sinapsi
dei cromosomi
omologhi;
pachitene, in cui avviene il crossing-over, con scambio del materiale genetico;
diplotene durante il quale inizia la desinapsi e i cromosomi restano appaiati tramite i
chiasmi;
diacinesi, in cui avviene la dissoluzione della membrana nucleare e del nucleolo.
Durante la profase I, inoltre, si sviluppa il fuso, costituito da due coppie di centrioli,
situate ai poli opposti della cellula, da cui fuoriescono fibre dimicrotubuli. Tali fibre
agganciano i cromosomi mediante il cinetocore, una piastra proteica situata a livello
del centromero. La profase I può durare per giorni o anche più a lungo e occupa il
90% del tempo richiesto per quasi tutta la divisione meiotica.
Metafase I
I cromosomi omologhi sono trascinati dai microtubuli all'equatore cellulare. Si ha
l'assortimento indipendente dei cromosomi omologhi. In pratica i cromosomi della
madre e del padre si allineano in modo casuale all'equatore.
Anafase I
A differenza dell'anafase mitotica, durante questa fase i cromatidi fratelli restano
attaccati per i centromeri, mentre i cromosomi omologhi si staccano e migrano ai poli
opposti della cellula. In questo modo si ha un corredo cromosomico aploide proprio
perché sono gli omologhi parentali a separarsi.
Telofase I
La telofase I può variare a seconda della specie. In seguito alla migrazione dei
cromosomi omologhi verso i poli opposti della cellula, si può verificare la formazione
della membrana nucleare e la citodieresi con la conseguente scissione cellulare, come
avveniva nella mitosi; oppure vi è la semplice migrazione dei cromosomi senza
scissione.
Interfase
In alcuni casi, terminata la Meiosi I, può avvenire l'Interfase in cui i cromosomi si
despiralizzano; in molte specie si passa invece direttamente dalla Telofase I alla
Profase II.
Meiosi II
La
seconda
divisione
meiotica
è
identica
alla mitosi,
solo
che
genera
due cellule aploidi, perché non è preceduta da un ciclo cellulare adeguatamente
fornito di fase S, e quindi avviene in presenza di un corredo cromosomico n invece
che 2n.
Profase II
Compaiono nuovamente le fibre del fuso che agganciano i cinetocori dei cromosomi.
Nel caso si sia verificata una scissione durante la telofase I, la membrana nucleare si
dissolve affinché i microtubuli del fuso possano attaccarsi ai cromosomi.
Metafase II
I cromosomi si toccano sulla piastra equatoriale; ogni cromosoma è costituito da 2
cromatidi omologhi fratelli.
Anafase II
I centromeri dei cromosomi dei cromatidi fratelli si staccano e i cromatidi si
dividono, migrando ai poli della cellula.
Telofase II
Ai poli opposti della cellula si cominciano a formare i nuclei e avviene la citodieresi,
con la conseguente scissione cellulare.
Ricapitolando: nella meiosi si passa da una cellula immatura con patrimonio genetico
(corredo cromosomico) diploide 2n con contenuto di cromatina 4C a quattro cellule
mature aploidi n con contenuto di cromatina 1C.
Sinapsi e crossing-over
La sinapsi è il processo che permette la spartizione dei cromosomi omologhi in due
corredi aploidi durante la profase I. Ciascun cromosoma è formato da due cromatidi,
e quindi quattro cromatidi formano una tetrade. In seguito alla sinapsi degli omologhi
avviene il crossing-over, un processo mediante il quale i cromosomi omologhi si
scambiano parti equivalenti, determinando nuove combinazioni di geni. Il risultato
visibile del crossing over è una struttura a croce chiamata chiasmo. In ciascun
chiasmo i cromosomi omologhi possono scambiarsi segmenti di cromatidi.
Mendel e le sue leggi
Gregor Johann Mendel (Hynčice, 20 luglio 1822 – Brno, 6 gennaio 1884) è stato
un biologo e un canonico agostiniano ceco, considerato, per le sue osservazioni sui
caratteri ereditari, il precursore della moderna genetica.
Il nome "Gregor" - con cui Mendel è oggi universalmente noto - gli fu in realtà
attribuito all'atto della sua ordinazione sacerdotale[1].
Il concetto base concepito dal monaco era molto innovativo, egli infatti dedusse che
l'ereditarietà era un fenomeno dovuto ad agenti specifici contenuti nei genitori, al
contrario di quanto creduto all'epoca. Non si può tuttavia ancora parlare di genetica.
Mendel identificò dopo sette anni di selezione sette "Linee pure": sette varietà di
pisello
che
differivano
per
caratteri
estremamente
visibili
(forma
del
seme: liscio o rugoso; colore del seme giallo o verde): e proprio grazie a queste
caratteristiche che tale pianta (Pisum sativum) si prestava particolarmente, oltre che a
un semplice sistema riproduttivo; il monaco poteva impollinare a piacimento i suoi
vegetali. Egli operò con un vastissimo numero di esemplari perché sapeva che le
leggi della probabilità si manifestano sui grandi numeri.
Mendel prese due varietà di piante di pisello completamente diverse,appartenenti alle
cosiddette linee pure (ovvero quelle nelle quali l'aspetto è rimasto costante dopo
numerose generazioni), ed iniziò ad incrociare le suddette per caratteri specularmente
diversi: ad esempio una pianta a fiori rossi con una pianta a fiori bianchi. Notò che la
prima generazione filiale (detta anche F1) manifestava soltanto uno dei caratteri delle
generazioni parentali (detta anche P) e ne dedusse che uno dei due caratteri doveva
essere dominante rispetto all'altro: da questa osservazione trae origine la legge sulla
dominanza. Incrociando poi tra loro le piante della generazione F1, Mendel osservò
la ricomparsa, in parte della successiva generazione, di caratteri "persi" nella F1 e
capì quindi che essi non erano realmente scomparsi, bensì erano stati "oscurati" da
quello dominante. Osservando la periodicità della seconda generazione filiale o F2,
(tre esemplari mostrano il gene dominante e uno il gene recessivo) Mendel portò le
scoperte ancora più avanti:
L'esistenza dei geni (detti in un primo momento caratteri determinanti
ereditari);
I fenotipi alternativi presenti nella F2 sono definiti da forme diverse dello
stesso gene, tali forme sono chiamate alleli;
per dare origine alla periodicità della F2 ogni tipo di gene deve essere presente,
nelle piante di pisello adulte, con due coppie per cellula che si segregano al
momento della produzione dei gameti.
Le leggi di Mendel
1. Legge della dominanza (o legge della omogeneità di fenotipo): Gli individui
nati dall'incrocio tra due individui omozigoti che differiscono per una coppia
allelica, avranno il fenotipo dato dall'allele dominante.
2. Legge della segregazione (o legge della disgiunzione): gli alleli di un
singolo locus segregano indipendentemente l'uno dall'altro (in seguito fu
evidente che ciò era dovuto all'indipendente segregazione degli autosomi).
3. Legge dell'assortimento indipendente (o legge di indipendenza dei caratteri): I
diversi alleli si trasmettono indipendentemente l'uno dagli altri,secondo precise
combinazioni
Esistono anche delle eccezioni alla legge della dominanza: una pianta può presentare
fiori di vario colore. Se si incrocia una pianta con fiori rossi con una con fiori bianchi,
si ottiene un fiore rosa (bocca di leone) perché entrambi i geni dei colori sono
dominanti; in questo caso si chiama dominazione incompleta.
Le leggi di Mendel si applicano solo a caratteri in cui il fenotipo deriva
dall'espressione di un singolo gene (come appunto i caratteri esaminati dall'abate),
non si possono applicare per caratteri dovuti all'interazione tra molti geni e l'ambiente
esterno (es. altezza, vigore, forza, produzione, capacità cognitive ecc).
Al principio del '900, con la riscoperta delle teorie di Mendel, le scienze
evoluzionistiche "incrociarono" le sue scoperte con le ipotesi di Darwin, si ebbe così
la nascita della cosiddetta "sintesi moderna", ovvero la teoria evolutiva più
autorevole, che rimase in auge fino agli anni '70 del '900. Questa teoria postulava la
graduale selezione dei caratteri più favorevoli, alla luce delle teorie genetiche,
seguendo un adattamento delle specie all'ambiente. Questa teoria è stata in parte
modificata e resa più rispondente alle prove empiriche dalla "teoria degli equilibri
punteggiati", che comunque riconosce le leggi di Mendel e il fondamentale contributo
della genetica per studiare i processi evolutivi.
La scoperta delle particelle subatomiche
Modelli atomici di Thomson e di Rutherford
Nella seconda metà del 1800 alcuni scienziati, indagando la natura dei fenomeni
elettrici, cominciarono a mettere in dubbio la validità dell'ipotesi di Dalton secondo
cui
l'atomo
era
indivisibile.
Gli
interrogativi
sorgevano,
in
particolare,
dall'osservazione che:
1. certe sostanze si scioglievano in acqua formando soluzioni in grado di
trasportare la corrente elettrica per la presenza di particelle cariche di elettricità
positiva (+) e negativa (−) dette ioni;
2. gas rarefatti all'interno di un tubo contenente due elettrodi (anodo e catodo)
collegati a un generatore elettrico producevano, in seguito al passaggio di
corrente elettrica, radiazioni dirette dal catodo all'anodo (raggi catodici).
Fu proprio studiando gli effetti delle scariche elettriche attraverso gas rarefatti in un
tubo catodico che il fisico inglese J. J. Thomson (1856-1940) giunse a stabilire (1897)
che i raggi catodici sono formati da particelle dotate di carica negativa presenti negli
atomi di tutti gli elementi. Tali particelle subatomiche, di massa molto più piccola di
qualsiasi atomo conosciuto, furono chiamate elettroni (e−). La carica dell'elettrone è
considerata la carica elementare negativa e per convenzione le viene attribuito valore
unitario −1.
Effettuando altre esperienze con un tubo a raggi catodici modificato (1886), erano
stati scoperti raggi aventi direzione opposta a quella dei raggi catodici, in quanto
associati a particelle dotate di carica positiva (raggi positivi,meglio noti come raggi
anodici o raggi canale). La massa di queste particelle risultava variabile in rapporto al
gas presente nel tubo e molto più grande della massa dell'elettrone (si trattava di ioni,
cioè di atomi privi di cariche negative). Lo stesso Thomson studiò in seguito questo
fenomeno e, insieme ad altri, osservò che la massa delle particelle positive costituenti
i raggi anodici assumeva un valore minimo quando il gas utilizzato era l'idrogeno. In
questo caso si formavano ioni idrogeno (H+) che, in anni seguenti, furono
definitivamente identificati come particelle subatomiche, chiamate protoni, presenti
negli atomi di tutti gli elementi. Il protone (p) è una particella dotata di carica
elettrica unitaria positiva (+1), e con una massa pari a 1836 volte quella dell'elettrone,
corrispondente, con buona approssimazione, a 1 unità di massa atomica (uma).
Il modello atomico di Thomson
Nel 1904, Thomson, in base ai dati di cui disponeva avanzò l'ipotesi, detto modello
atomico di Thomson, secondo cui l'atomo era rappresentabilecome una massa sferica
con cariche elettriche positive uniformemente distribuite e contenente immersi nel
suo interno un ugual numero di elettroni, in modo che il tutto risultasse elettricamente
neutro.
Il modello di Thomson si rivelò ben presto inadeguato a spiegare una serie di
fenomeni fisici.
Il modello atomico di Rutherford
Un importante passo successivo nella comprensione della struttura dell'atomo fu
compiuto dal fisico neozelandese E. Rutherford (1911), attraverso l'impiego di
particelle alfa (α) emesse da un materiale radioattivo, per bombardare un bersaglio
costituito da una sottilissima lamina d'oro. La maggior parte delle particelle α
attraversava la lamina metallica in linea retta, mentre una loro piccolissima frazione
veniva deviata o addirittura respinta dalla lamina. Rutherford ne dedusse che le
particelle α potevano essere deviate o respinte solo ammettendo che gli atomi siano
formati da una piccolissima zona centrale, detta nucleo, di carica positiva e nella
quale è concentrata tutta la massa dell'atomo e da un grande spazio circostante dove
sono presenti elettroni che ruotano intorno al nucleo secondo orbite circolari (modello
di Rutherford o modello atomico planetario, 1911).
Rutherford valutò in seguito che la carica positiva del nucleo corrisponde al numero
di protoni. Tale numero è detto numero atomico (simbolo Z) e in un atomo neutro
corrisponde al numero degli elettroni.
Rutherford rilevò inoltre che la massa del nucleo calcolata sulla base del numero di
protoni, risultava sempre inferiore alla massa reale dell'atomo. Era quindi ipotizzabile
la presenza nel nucleo di un altro tipo di particelle.
Queste particelle, chiamate neutroni (n), furono poi individuate nel 1932 dal fisico
inglese J. Chadwick: esse risultarono prive di carica e dotate di una massa circa
uguale a quella del protone.
Riassumendo,
un atomo è
formato
da
un nucleo, costituito
da protoni e
neutroni (detti nucleoni), intorno al quale sono disposti elettroni (v. tab. 3.1).
In un atomo elettricamente neutro, il numero dei protoni, detto numero atomico
(Z) è uguale al numero degli elettroni. Il numero atomico è caratteristico di ogni
elemento. La somma del numero dei protoni e del numero dei neutroni (N) è detto
numero di massa (A):
Un atomo di un elemento di cui sono noti il numero atomico (Z) e il numero di massa
(A) è detto nuclide. Per un generico elemento di simbolo X il rispettivo nuclide viene
così rappresentato:
Esempio: 818O (o anche ossigeno −18) indica il nuclide dell'ossigeno con numero
atomico 8 e numero di massa 18.
La tavola periodica
La tavola periodica degli elementi è lo schema con il quale vengono ordinati
gli atomi sulla base del loro numero atomico Z.
Ideata dal chimico russo Dimitrij Mendeleev nel 1869, contemporaneamente ed
indipendentemente
dal
chimico
tedesco Julius
Lothar
Meyer(1830 -
1895),[1] inizialmente contava numerosi spazi vuoti, previsti per gli elementi che
sarebbero stati scoperti in futuro, taluni nella seconda metà del 1900.
In onore del chimico russo, la tavola periodica degli elementi è anche detta tavola
periodica di Mendeleev.
La tavola periodica si articola in gruppi e periodi:
ogni gruppo (colonna della tabella) comprende gli elementi che hanno la stessa
configurazione elettronica esterna (modo in cui gli elettronisi dispongono attorno
al nucleo). All'interno di ogni gruppo si trovano elementi con caratteristiche
chimiche simili.
ogni periodo (riga delle tabella) inizia con un elemento il cui atomo ha come
configurazione elettronica esterna un elettrone di tipo s, o nsdove n è il numero
quantico principale, e procedendo verso gli atomi successivi del periodo, il
numero atomico Z aumenta di una unità ad ogni passaggio.
La necessità di ordinare le conoscenze che (grazie a ricerche empiriche) venivano
accumulandosi sulle diverse sostanze venne sentita fin dai primordi della chimica,
e Lavoisier propose (nel 1789) una prima forma di sistematica chimica. Bisogna,
però, aspettare all'incirca il 1870 affinché si giunga al modello che, con le opportune
aggiunte, è utilizzato ai giorni nostri.
La tavola originaria fu creata prima della scoperta delle particelle subatomiche o della
formulazione delle teorie attuali per quanto riguarda la meccanica quantistica e
l'atomo.
Se gli elementi vengono disposti in ordine crescente di numero atomico, inserendo in
seguito altre proprietà, si può notare una ondulazione o una periodicità di queste
proprietà in funzione del numero atomico dell'elemento stesso. Il primo che
riconobbe queste ricorrenze fu il chimico tedesco Johann Wolfgang Döbereiner, che
nel 1829 per primo notò una certa quantità di triadi, gruppi di tre elementi con queste
similarità.
A questa intuizione fece seguito l'inglese John Newlands, che sottolineò (nel 1865)
come gli elementi di tipo simile fossero ricorrenti ad intervalli regolari di 8 posizioni,
che lui assimilò alle ottave musicali,anche se questa sua cosiddetta legge delle
ottave venne messa in ridicolo dai suoi contemporanei.
Fu proprio in quel periodo che Meyer e Mendeleev (indipendentemente l'uno
dall'altro) assunsero il peso atomico come parametro per la classificazione periodica
degli elementi, il che costituì un passo decisivo verso il più raffinato concetto
di numero atomico (del quale, nella chimica odierna, le proprietà fisico-chimiche
degli elementi sono considerate funzioni periodiche). Meyer pubblicò i propri risultati
qualche mese dopo Mendeleev, ed è per questo che molto spesso ci si riferisce alla
tavola periodica degli elementi col solo nome di quest'ultimo. Egli ipotizzò anche che
esistessero altri elementi, al momento non conosciuti, che occupavano le celle vuote
della tabella; teoria che trovò conferme con la scoperta della struttura elettronica
degli elementi tra la fine del XIX e gli inizi del XX secolo.
Negli anni '40, Glenn Theodore Seaborg sintetizzò e separò con metodi radiochimici i
primi 5 elementi transuranici, che sono stati sistemati all'interno della tabella (o in
alcuni casi al di sotto, come si può vedere dallo schema). Negli anni successivi i
primi di tali transuranici furono identificati in "ultratracce" nei minerali uraniferi,
come prodotti naturali di attivazione. Dal 2003 la IUPAC ha attribuito il nome
definitivo alla serie 4f dei lantanoidi Ln e 5f degli attinoidi An.[2] [3]
La ragione della particolare periodicità per serie di lunghezza diversa (2, 8 , 8, 18, 18,
32, 32) è stata scoperta solo in seguito (ad opera principalmente di Niels Bohr), ed è
da ricercarsi nella tendenza al massimo riempimento possibile degli orbitali
atomici da parte degli elettroni, che ha valore diverso a seconda del tipo di orbitale
interessato, e al modo in cui si susseguono i diversi tipi orbitali per numeri atomici
crescenti; si hanno infatti al massimo due elettroni per orbitale di tipo s, al più sei
elettroni per orbitali di tipo p, al più dieci per orbitali d, ed al più quattordici per
orbitali tipo f; inoltre gli orbitali si possono susseguire solo nell'ordine: 1s 2s 2p 3s 3p
4s 3d 4p 5s 4d 5p 6s 4f 5d 6p 7s 5f 6d 7p 8s (secondo la regola di Aufbau).[4]
A diversi gradi di riempimento dell'orbitale più esterno corrisponde una diversa
reattività dell'intero atomo, per cui ad orbitali "completi" corrisponde la
configurazione energetica più stabile (e quindi una reattività nulla), e ad orbitale
esterno parzialmente "completo" corrispondono reattività via via maggiori quanto più
il numero di elettroni si allontana da quello di una configurazione stabile; questa
diversa reattività a livello macroscopico determina molte delle proprietà chimiche
dell'elemento, che si ripetono in modo simile al crescere del numero atomico Z.
Pertanto, a diversi numeri atomici (ossia ad elementi diversi ordinati per massa)
corrispondono diverse proprietà chimiche macroscopiche, che si ripetono
periodicamente in modo simile per configurazioni elettroniche che hanno una
stabilità energetica comparabile, e questo accade per analoghi gradi di riempimento
dell'orbitale, che si ripetono con periodo variabile perché variabile è il massimo
riempimento dei vari orbitali.
La forma corretta della tavola periodica degli elementi fu pertanto inizialmente
determinata solo da osservazioni macroscopiche, senza la presenza di un modello
microscopico a cui fare riferimento. Successivamente questa forma è stata
"confermata" con la scoperta della struttura microscopica degli atomi e quindi delle
loro modalità di interazione.
Le reazioni chimiche
Una reazione chimica è una trasformazione della materia che avviene senza
variazioni misurabili di massa, in cui uno o più reagenti iniziali modificano la loro
struttura e composizione originaria per generare i prodotti.
Alcuni processi in cui intervengono reazioni chimiche sono:
la corrosione del ferro a ruggine (che è composta da ossidi di ferro);
la combustione del metano o altri combustibili (il metano con l'ossigeno si
trasforma in anidride carbonica evapore acqueo);
la digestione (gli alimenti sono decomposti dai succhi gastrici in sostanze
chimiche assimilabili dall'organismo).
La materia è composta da atomi. Ogni atomo possiede proprietà peculiari, derivanti
dalla suastruttura atomica. Gli atomi possono legarsi tra loro per formare le molecole.
Un composto chimico è un tipo particolare di molecola nella quale gli atomi sono
diversi tra loro.
Ad esempio, l'ossigeno forma una molecola fatta con due atomi di ossigeno, mentre
l'acqua è una molecola composta da due atomi di idrogeno legati ad un atomo di
ossigeno, e quindi è anche un composto chimico.
Le molecole si formano attraverso una reazione chimica, che consiste in una rottura e
formazione di legami chimici tra atomi. Più in generale, le reazioni chimiche possono
coinvolgere anche altre specie chimiche (ioni, radicali, ecc.) oltre le molecole.
Le reazioni chimiche non provocano un cambiamento di natura della materia, perché
non influenzano i suoi costituenti fondamentali (gli atomi) ma solo la maniera in cui
sono aggregati in molecole; non influenzano nemmeno l'aggregazione di molecole
simili, quindi le trasformazioni puramente fisiche, come i cambiamenti di
stato (fusione, solidificazione, evaporazione, ebollizione, ecc.), l'usura e l'erosione,
lafrattura, ecc. non sono reazioni chimiche.
Allo stesso modo, non fanno parte delle reazioni chimiche le trasformazioni
dei nuclei atomici, cioè le reazioni nucleari. Tuttavia tali reazioni assumono anche un
certo interesse in chimica e vengono studiate dalla chimica nucleare.
Le reazioni chimiche, dunque, riguardano esclusivamente le variazioni dei legami tra
gli atomi (legame covalente, legame ionico, legame metallico).
Una reazione può sviluppare calore, in tal caso è detta esotermica, o assorbire calore,
ed essere quindi endotermica. Una reazione esotermica è quindi una reazione che
comporta un trasferimento di calore dal sistema all'ambiente. Similmente
una reazione endotermica è una reazione che comporta un trasferimento di calore
dall'ambiente al sistema. Necessita dunque di energia esterna per procedere.
Il sistema è la parte dell'universo oggetto di studio (nel nostro caso sistema chimico,
ad es. solvente, reagenti e prodotti presenti in un becher(che rappresenta il contorno
del sistema), mentre l'ambiente è tutto ciò che circonda il sistema stesso. Sistema +
ambiente costituiscono un sistema isolato: l'universo è un sistema isolato.
I composti chimici presenti all'inizio della reazione sono detti reagenti, quelli che si
ottengono alla fine della reazione sono invece i prodotti di reazione.
I fenomeni che hanno luogo durante una reazione chimica vengono rappresentati
mediante una equazione chimica. Un'equazione chimica è scritta in maniera simile ad
un'equazione matematica, ed in essa compaiono due membri: al primo membro (cioè
a sinistra della freccia o altro simbolo di reazione) compaiono i reagenti, mentre al
secondo membro (cioè a destra della freccia o altro simbolo di reazione) stanno i
prodotti.
Una reazione non può avere luogo, o viene rallentata fino a fermarsi o addirittura a
regredire se non è soddisfatta una serie di condizioni, come presenza dei reagenti in
misura adeguata e condizioni di temperatura, pressione e luce adatte alla specifica
reazione.
Dal postulato fondamentale di Lavoisier, che dice: nulla si crea, nulla si distrugge,
tutto si trasforma ne deriva necessariamente che la somma delle masse dei reagenti è
necessariamente uguale alla somma delle masse dei prodotti di reazione. Siccome
la materia è costituita da atomi, anche il numero degli atomi a destra e a sinistra
dell'equazione deve restare invariato.[1] Ad esempio nell'equazione:
che
rappresenta
la
reazione
tra idrossido
di
sodio ed acido
cloridrico per
produrre cloruro di sodio, che conosciamo bene come sale da cucina, troviamo
esattamente lo stesso numero di atomi dello stesso tipo sia nella parte sinistra
(reagenti) che nella parte destra (prodotti) della reazione, ma combinati in maniera
diversa.
In questo caso, essendo questa una reazione tra un acido (HCl) e una base (NaOH) la
reazione procederà verso la neutralizzazione completa, a meno che uno dei reagenti
non sia in eccesso rispetto all'altro: in questo caso, la soluzione rimarrà acida o basica
a seconda del reagente in eccesso.
Oltre al bilanciamento delle masse, nelle equazioni chimiche deve essere soddisfatto
il bilanciamento delle cariche.[1] Le reazioni chimiche infatti possono avvenire anche
tra specie chimiche cariche elettricamente, dette ioni.
Alcune reazioni per avvenire hanno bisogno, o vengono facilitate, della presenza di
una terza sostanza (rispetto a reagenti e prodotti) dettacatalizzatore.
Il catalizzatore permette o facilita la reazione, ma viene ritrovato invariato (o quasi)
tra i prodotti di reazione. In biologia i catalizzatori sono denominati enzimi.
Le trasformazioni che hanno luogo durante una reazione chimica spontanea portano
ad una diminuzione dell'energia totale del sistema. In effetti, in una molecola o in
un cristallo, l'organizzazione reciproca degli atomi implica un'energia, l'energia di
legame; perché un legame venga rotto è necessario fornire al sistema una quantità di
energia almeno pari all'energia di legame. Quando gli atomi si ricombinano,
formando nuovi legami, tale energia viene liberata. Al termine di una reazione,
l'energia immagazzinata nei legami dei prodotti di reazione è minore di quella
inizialmente presente nei legami dei reagenti iniziali.
Durante la reazione, tuttavia, esiste un momento in cui i vecchi legami si sono rotti e
quelli nuovi non si sono ancora formati, è lo stato di transizione dove l'energia del
sistema è massima, cosa che costituisce una vera barriera per la realizzazione della
reazione (vedi: energia di attivazione).
Lo studio dell'aspetto energetico delle reazioni chimiche è la termodinamica, che ci
permette di verificare se una reazione può o meno avere luogo e quanta energia è
necessario fornire per superare la barriera dell'energia di attivazione; ma esiste un
altro parametro importante: lavelocità di reazione.
Alcune reazioni sono molto rapide, addirittura violente, come le esplosioni, altre sono
talmente lente che possono continuare per anni, o secoli. Alcune sono talmente lente
che i reagenti coinvolti sembrano in realtà composti stabili, come nel caso
dell'ossidazione dell'alluminio, si parla in tal caso di composti "metastabili" (la forma
stabile, in ambiente con presenza di ossigeno, è l'ossido di alluminio, mentre quella
metastabile è l'alluminio metallico); ad occuparsi di studiare la velocità di reazione è
la cinetica chimica.
Per quantificare la velocità di una reazione si utilizza il grado di avanzamento della
reazione α, definito globalmente come la proporzione dimiscela che ha già reagito
(α=0 all'inizio della reazione, α=1 quando la reazione è completa). Si può così
definire la velocità di reazione come laderivata del grado di avanzamento rispetto al
tempo:
Alcune reazioni sono reversibili, cioè il guadagno di energia avuto con la reazione è
minimo, in tal modo risulta possibile anche la reazione inversa; è questo il caso
della dissociazione dell'acqua, H2O, negli ioni: H3O+ e OH-. In questi casi il sistema
evolve in generale verso un equilibrio dinamico, ossia il valore di α rimane stabile e
compreso tra 0 e 1, il numero di molecole che reagiscono in un senso è quindi
compensato dal numero di molecole che reagiscono nell'altro.
La cinetica di una reazione dipende da numerosi fattori, il più importante è la
temperatura: l'energia termica permette sia di superare la barriera dell'energia di
attivazione più facilmente, sia di avere un numero maggiore di collisioni tra le
molecole reagenti.
Un altro parametro importante è la fase in cui si trovano i reagenti. Da questo punto
di vista le reazioni maggiormente favorite sono quelle in fase gassosa o liquida, dove
i reagenti sono mescolati tra loro e possono facilmente venire a contatto.
In tutti gli altri casi, cioè per reazioni tra:
un solido e un gas;
un solido e un liquido;
un solido e un solido;
un liquido e un gas;
due liquidi immiscibili;
dette reazioni eterogenee, la reazione può aver luogo esclusivamente nei punti di
contatto tra le due fasi, quindi sarà più veloce se i reagenti vengono dispersi l'uno
nell'altro come nel caso di:
aerosol (fini gocce di liquido disperse in un gas);
emulsioni (dispersioni di gocce di un liquido in un altro immiscibile);
miscugli di polveri;
sol (dispersioni di polveri in un liquido);
schiume (bolle di gas disperse in un liquido).
in questo modo vengono massimizzate le superfici di contatto tra i reagenti e quindi
la possibilità di reazione.
Per i solidi questo può esser quantificato misurando la superficie specifica, ossia la
superficie esposta per unità di massa; una polvere o un solido poroso hanno elevati
valori di superficie specifica.
A seconda del modo in cui si combinano i reagenti per dare luogo ai prodotti, si
possono avere le seguenti tipologie di reazioni chimiche:
Decomposizione: un reagente da luogo a più prodotti;
Sintesi: più reagenti danno luogo a un prodotto;
Sostituzione: un gruppo di una specie chimica viene sostituite da un altro
gruppo;
Metatesi: scambio di due o più ioni fra elementi e gruppi aventi la stessa
valenza.
Una reazione viene detta di ossido-riduzione (o redox) se durante il suo svolgimento
alcune specie chimiche modificano il proprio numero di ossidazione.[2] Le reazioni
che non sono di ossido-riduzione sono reazioni acido-base (ovvero i reagenti di tali
reazioni sono un acido e una base).
Il metabolismo del corpo umano: la digestione
La digestione è il processo meccanico-chimico che trasforma e riduce il cibo ingerito
in sostanze più semplici e più facili da assorbire ed assimilare dall'organismo. La
digestione è una forma di catabolismo ovverosia la riduzione di molecole complesse
del cibo in molecole più semplici
La digestione comincia dalla bocca, che viene aperta o chiusa da una parte fissa,
la mascella, a cui si articola una parte mobile, la mandibola. La bocca contiene
la lingua. Il cibo ingerito riceve dalle ghiandole salivari la saliva, viene portato alla
temperatura ottimale per la digestione ed è rotto meccanicamente dai denti, mentre è
in parte demolito chimicamente dalla saliva; di questa ne viene prodotta circa
un litro e mezzo ogni giorno dalle ghiandole salivari. La saliva è prodotta da tre
ghiandole: la parotide, la più grande, situata sotto l'orecchio, la sottomascellare e
lasottolinguale. La saliva è una soluzione acquosa al 99.5 %, e contiene
l'amilasi salivare, un enzima che a pH 7 inizia la demolizione degli amidiin
frammenti più piccoli, come il maltosio, costituito da due molecole di glucosio. Il
cibo, impregnato di saliva e impastato con i movimenti della mascella e della lingua,
forma il bolo, che viene spinto nella faringe dove viene deglutito. La faringe fa parte
anche dell'apparato respiratorio ed è la via attraverso cui l'aria entra nella laringe e
quindi passa nella trachea. Durante la deglutizione la respirazione cessa e l'entrata
della laringe è coperta dall'epiglottide, in modo che il bolo imbocchi la via giusta,
cadendo nell'esofago. L'esofago, un tubo lungo anche 30 cm, attraversa il diaframma,
e termina nello stomaco. Il diaframma è il muscolo a forma di cupola che separa la
cavità toracica dalla cavità addominale. Nell'esofago il cibo si muove spinto da
contrazioni peristaltiche che avanzano come onde restringendo il passaggio al di
sopra del bolo e allargandolo al disotto. Esse ci permettono di inghiottire e far
avanzare il cibo nell'esofago anche se ci troviamo a testa in giù.
L'ingresso del bolo nello stomaco è controllato da un muscolo sfintere, il cardias. Lo
stomaco è come un sacco dalla capacità di più di un litro, dalle robuste pareti
muscolari. La parete più interna dello stomaco, la mucosa, presenta numerosi
microvilli all'interno delle quali si annidano le ghiandole che, a ogni pasto, producono
circa 500 ml di succo gastrico, composto da enzimi, muco e acido cloridrico. L'acido
cloridrico dissolve la sostanza cementante posta tra le cellule del cibino, uccide i
batteri e rende attivo un enzima secreto dalle ghiandole gastriche, il pepsinogeno.
Quando, nella cavità dello stomaco, il pepsinogeno è raggiunto dall'acido cloridrico,
esso diventa pepsina (dal greco pèpsis = cottura). Mentre il pepsinogeno è inattivo e
non può danneggiare le cellule delle ghiandole che lo producono, la pepsina agisce
sui legami peptidici che legano tra loro gli amminoacidi delle proteine. Si formano
così piccoli frammenti di proteine, i peptidi, composti da alcuni amminoacidi ancora
legati tra loro. La mucosa dello stomaco è rivestita da una patina di muco, che ha la
funzione di proteggere le cellule dai succhi digestivi. La protezione è indispensabile
perché essi, così come attaccano e digeriscono gli alimenti, potrebbero attaccare e
digerire la stessa parete dello stomaco. Talvolta però, la mucosa può essere
danneggiata da sostanze irritanti come l'alcol o l'acido acetilsalicilico; oppure il muco
può venire prodotto in quantità insufficiente. Quando ciò si verifica, gli enzimi e
l'acido cloridrico iniziano a "digerire" lo stomaco: si forma una lesione
chiamata ulcera peptica. Il sintomo tipico è un dolore profondo nella parte superiore
dell'addome a stomaco vuoto. Recentemente si è scoperto che nello stomaco di molti
individui è presente un batterio, l'Helicobacter pylori, che potrebbe avere un ruolo
importante nell'insorgenza del disturbo. Lo stomaco ha funzione di dissolvimento e di
digestione, ma in genere non di assorbimento. Tuttavia alcune sostanze come l'acqua,
le vitamine, l'aspirina, il glucosio e l'alcool, possono essere direttamente assorbite
nello stomaco senza arrivare nell'intestino. Ciò avviene perché le loro molecole, di
piccole dimensioni, passano direttamente nel sangue che scorre nei vasi delle pareti
dello stomaco. Per questo motivo l'alcool, anche se ingerito da poco, può avere effetti
quasi immediati. Il contenuto dello stomaco, quando esce per entrare nell'intestino, è
una poltiglia semi-solida lattiginosa e acida (il pH dello stomaco è 1-2) ed è
detto chimo.
L'intestino si distingue in intestino tenue,lungo e sottile, e intestino crasso, più corto,
con un diametro maggiore e con una superficie solcata da profonde pieghe.
Nell'intestino si completa la digestione chimica e ha luogo l'assorbimento del
materiale digerito. Al fine di massimizzare l'area disponibile per queste funzioni,
l'intestino tenue è estremamente lungo ed è altamente ripiegato. È diviso in tre
sezioni: duodeno, digiuno e ileo.
Pur essendo lungo 30 cm,ovvero 12 pollici, il duodeno è la porzione dell'intestino e
dell'intero apparato digerente che sostiene la maggior parte della digestione chimica.
Le trasformazioni avvengono per opera del succo enterico (dal greco énteron =
intestino), secreto da ghiandole situate nella mucosa del duodeno, del succo
pancreatico, prodotto dal pancreas, e della bile, prodotta dalfegato. Il succo
pancreatico è prodotto dal pancreas, un organo di forma allungata situato vicino al
punto in cui lo stomaco comunica con l'intestino. In esso vi sono due distinti
raggruppamenti di cellule. In uno viene prodotto il succo pancreatico che con un
condotto, il dotto pancreatico, viene convogliato nell'intestino. Nell'altro le cellule
formano dei piccoli ammassi, detti isole di Langerhans, in cui sono prodotti ormoni,
come l'insulina, che sono riversati nel sangue. Il succo enterico e quello pancreatico
contengono numerosi enzimi, muco e sali, tra cui il bicarbonato che ha la funzione di
neutralizzare il pH acido del chimo, riportandolo vicino alla neutralità. Gli enzimi del
succo
enterico
e
pancreatico
agiscono
su zuccheri, proteine, grassi e acidi
nucleici demolendoli in molecole più piccole. Enzimi sono: l'amilasi per la digestione
dell'amido, la lipasi per la digestione dei grassi, latripsina, contenuta nel succo
pancreatico, per la digestione delle proteine.
La digestione dei grassi è operata dagli enzimi, ma è resa possibile dalla bile.
La bile è un liquido giallo-verde che viene immagazzinato in una piccola ampolla
annessa al fegato, la cistifellea, un serbatoio che circa 30 minuti dopo il pasto spruzza
il suo contenuto, mediante un condotto, ilcoledoco, nell'intestino. I sali biliari
contenuti nella bile hanno una struttura simile a quella dei detergenti, con una
estremità idrosolubile e un'estremità liposolubile. Il loro ruolo è quello di emulsionare
i grassi, ossia di suddividere le grosse gocce di grasso in numerose goccioline più
piccole che restano separate le une dalle altre. Questo processo ha la funzione di
aumentare la superficie su cui possono agire gli enzimi. In conclusione, nell'intestino,
per l'azione chimica dei succhi digestivi e per quella meccanica dei movimenti
peristaltici, il chimo proveniente dallo stomaco viene trasformato in un liquido
lattescente, detto chilo, che contiene in soluzione molecole piccole che possono
attraversare la parete intestinale. Infatti, gli amidi che in bocca erano già stati
intaccati dall'amilasi salivare sono completamente demoliti in glucosio. Le lunghe
catene di amminoacidi delle proteine, già ridotte nello stomaco in peptidi, sono
completamente
demolite
in amminoacidi .
Anche
gli acidi
nucleici,
pur
rappresentando una quota molto piccola degli alimenti, sono decomposti
nei nucleotidi che li costituiscono. I grassi che non avevano subito né in bocca né
nello stomaco alcun attacco enzimatico, sono demoliti in glicerolo e acidi grassi.
L'ultima fase della digestione, l'assorbimento, è il passaggio nel sangue, attraverso le
pareti
intestinali,
delle
sostanze
ottenute
dalla
digestione
delle macromolecole contenute negli alimenti: principalmente glucosio (derivato da
amido e saccarosio), amminoacidi(derivati delle proteine), glicerolo e acidi grassi
(derivati dai grassi), vitamine e sali minerali. Gran parte dell'assorbimento ha luogo
attraverso le pareti del digiuno e dell'ileo. Per aumentare la superficie di
assorbimento, le pareti dell'intestino tenue sono tutte sollevate in pieghe o pliche a
loro volta ricoperte da migliaia di sottili estroflessioni a forma di dito, i villi. Ogni
villo è lungo circa 1 mm e ce ne sono circa 3000 per ogni centimetro quadrato. La
superficie di ogni villo è ulteriormente aumentata perché le cellule che lo formano
hanno la loro stessa membrana sollevata in migliaia di microvilli. Le molecole
passano attraverso la membrana dei microvilli. Ogni villo è percorso all'interno da
una rete di capillari in cui scorre il sangue e qui, nel sangue, si riversano il glucosio,
gli amminoacidi, i sali e le vitamine. I capillari intestinali convergono infine in un
vaso sanguigno, la vena porta epatica, che entra nel fegato. I grassi seguono un'altra
via. Dopo aver superato la membrana dei microvilli, il glicerolo e gli acidi grassi si
riuniscono e formano di nuovo i trigliceridi, che vanno infine in un piccolo condotto,
un vaso linfatico in cui scorre la linfa. La linfa è un liquido che ha una composizione
simile al sangue, ma senza globuli rossi. Essa scorre in un sistema di tubicini, i vasi
linfatici. Dopo un certo percorso i vasi linfatici confluiscono in un unico condotto che
termina nel torrente circolatorio. La linfa si unisce quindi al sangue e in quest'ultimo
arrivano, direttamente o indirettamente, tutte le molecole provenienti dalla digestione
del cibo. Il contenuto intestinale, dopo l'assorbimento nell'intestino tenue, prosegue
lentamente il suo cammino nell'intestino crasso. Il primo tratto, a forma di sacca, è
detto cieco perché porta un'estroflessione vermiforme, l'appendice, che nell'uomo ha
un ruolo secondario in quanto contribuisce in piccola parte alle difese immunitarie.
La sua infezione è detta appendicite. Al cieco segue il colon, il quale è percorso
trasversalmente da solchi e dotato di una forte muscolatura. Il colon termina con il
retto, che sbocca all'esterno con un muscolo sfintere, l'ano. Nell'intestino crasso le
ghiandole della mucosa producono solo muco e non enzimi. Anche se la parete è
lubrificata dal muco, il tempo di transito dei materiali intestinali è piuttosto lungo,
dalle 12 alle 36 ore. Nel crasso avviene il riassorbimento dell'acqua e dei sali
minerali, e l'eliminazione con le feci del cibo non digerito. Il riassorbimento
dell'acqua è importante perché ogni giorno vengono riversati nel tubo digerente sotto
forma di succhi digestivi ben 7 litri di liquidi. Se il materiale digerito si muove troppo
velocemente lungo il colon, si ha un riassorbimento insufficiente di acqua che
provoca diarrea e disidratazione; al contrario, se il movimento è troppo lento, l'acqua
viene riassorbita in quantità eccessiva, causando stitichezza.
La tassonomia
La tassonomia (dal greco ταξις, taxis, "ordinamento", e νοµος, nomos, "norma" o
"regola") è, nel suo significato più generale, la scienza della classificazione.
Abitualmente, si impiega il termine per designare la tassonomia biologica, la scienza
dell'ordinare gli organismi in un sistema di classificazione composto da una gerarchia
di taxa annidati.
Con il termine tassonomia, dunque, ci si può riferire sia alla classificazione
gerarchica di concetti, sia al principio stesso della classificazione. Praticamente tutti i
concetti, gli oggetti animati e non, i luoghi e gli eventi possono essere classificati
seguendo uno schema tassonomico. La tassonomia è la scienza che si occupa
genericamente dei modi di classificazione (degli esseri viventi e non).
Per classificazione si intende la descrizione e la collocazione in un sistema
tassonomico di una entità, mentre per determinazione si intende il riconoscimento o
l'identificazione
di
un
soggetto.
Soprattutto
in
ambito
scientifico
(es.botanica, zoologia) è importante non confondere questi termini.
Secondo la matematica, una tassonomia è una struttura ad albero di istanze (o
categorie) appartenenti ad un dato gruppo di concetti. A capo della struttura c'è
un'istanza singola, ilnodo radice, le cui proprietà si applicano a tutte le altre istanze
della gerarchia (sotto-categorie). I nodi sottostanti a questa radice costituiscono
categorie più specifiche le cui proprietà caratterizzano il sotto-gruppo del totale degli
oggetti classificati nell'intera tassonomia.
In biologia è la disciplina scientifica che si occupa di attribuire un nome agli
organismi viventi e di classificarli. Di conseguenza, la Tassonomia Biologica è una
sottodisciplina della Biologia Sistematica, che studia le relazioni di parentela fra gli
organismi e la loro storia evolutiva. Attualmente, la Tassonomia agisce dopo avere
risolto l'albero filogenetico degli organismi studiati, cioè, una volta che vengono
risolti i cladi, o rami evolutivi, in funzione delle relazioni di parentela fra essi.
Attualmente esiste il consenso nella comunità scientifica del fatto che la
classificazione
deve
essere
interamente
consistente
con
ciò
che
si
sa
della filogenesi dei taxa, poiché solo allora fornirà il servizio che si spera essa dia al
resto dei rami della Biologia (vedere per esempio Soltis e Soltis 2003 [1]), ma ci sono
scuole dentro la Biologia Sistematica che definiscono con sfumature differenti la
maniera nella quale la classificazione deve corrispondere alla filogenesi conosciuta.
Più in là della scuola che la definisce, il fine ultimo della Tassonomia è organizzare
l'albero
filogenetico
in
un sistema
di
classificazione.
Perciò,
la
scuola cladistica (quella che predomina al giorno d'oggi) converte i cladi in taxa.
Un taxon è un clade al quale fu assegnato unacategoria tassonomica e un nome in
latino, di cui si faceva una descrizione, associandone un esemplare "tipo", e che fu
pubblicato in una rivista scientifica. Quando si fa tutto questo, il taxon ha un nome
corretto. La nomenclatura è la sottodisciplina che si occupa di regolamentare questi
passi, e far sì che ci si attenga ai principi di nomenclatura. Di conseguenza, i sistemi
di clasificazione che ne nascono, funzionano come contenitori di informazione da un
lato, e come predittivi dall'altro.
Una volta terminata la classificazione di un taxon, si estraggono i caratteri
diagnostici di ognuno dei suoi membri, e sopra questa base si confezionano chiavi
dicotomiche di identificazione, le quali sono utilizzate allo scopo di determinare o
identificare gli organismi, ubicando un organismo sconosciuto in un taxon conosciuto
del sistema di classificazione dato. La determinazione o identificazione è inoltre la
specialità, dentro la tassonomia, che si occupa dei principi di elaborazione delle chavi
dicotomiche e altri strumenti utilizzati allo stesso fine.
Le norme che regolano la creazione dei sistemi di classificazione sono in parte
convenzioni più o meno arbitrarie. Per comprendere queste arbitrarietà (per esempio,
la nomenclatura binomiale delle specie e la uninominale delle categorie superiori
alla specie, o anche la quantità di categorie tassonomiche e i nomi delle stesse) è
necessario studiare la storia della Tassonomia, che ci ha lasciato come eredità
i Codici Internazionali di Nomenclatura alle cui regole tecniche debbono attenersi i
sistemi di classificazione.
Oltre alla scuola che la definisce, il fine ultimo della tassonomia è presentare
un sistema di classificazione che raggruppi tutte le diversità degli organismi in unità
discrete dentro un sistema stabile, sopra le quali sia reso possibile il lavoro dei
ricercatori.
I sistemi di classficazione sono composti da taxa (dal greco ταξα, taxa) inseriti nelle
loro rispettive categorie tassonomiche. La decisione di quali cladi dovrebbero
convertirsi in taxa e in quali categorie tassonomiche dovrebbe stare ogni taxon, è un
po' arbitraria, però ci sono alcune regole non scritte che i ricercatori utilizzano
affinché il sistema di classificazione risulti "utile". Affinché un sistema di
classificazione risulti utile, esso deve essere maneggevole, e perciò deve organizzare
l'informazione in modo che sia più facile da ricordare. Judd e collaboradori (2002)
concordano sul fatto che:
1. ogni taxon deve avere evidenza affidabile del fatto che esso formi un
gruppo monofiletico: per convertire un clade in taxon deve avere
molte sinapomorfie che lo giustifichino, e deve avere una quantità di caratteri
diagnostici che permettano di diferenziarlo dal resto dei taxa, agevolando la
stabilità del sistema di classificazione;
2. alcuni sistematici appoggiano l'idea del fatto che ogni taxon dovrebbe
possedere caratteri morfologici ovvi che permettano di identificarlo, e ciò
favorirebbe l'identificazione per quelli non sistematici, oltre che aiutare a
inferire molti aspetti della sua biologia;
3. i taxa che compongono un sistema di classificazione devono avere
possibilmente fra 3 e 7 sottotaxa, un numero che possa maneggiare con facilità
la memoria umana (Stevens 1998[8]). Nelle parole di Davis e Heywood
(1963:83): "Dobbiamo essere capaci di ubicare i taxa in taxa di categoria più
alta in modo che possiamo incontrarli di nuovo".[16]
4. Altro criterio è la stabilità della nomenclatura. I gruppi che già sono stati
nominati nel passato dovrebbero continuare con lo stesso nome nei limiti del
possibile.
Una volta deciso quali cladi convertire in taxa, i sistematici devono decidere in quali
categorie tassonomiche ubicarli, il che è arbitrario. Per ragioni storiche si utilizzano
le categorie
linneane di
classificazione: regno, phylum o divisione, classe, ordine, famiglia, genere e specie (v
edere nella sezione della storia della tassonomia). Gli stessi criteri utilizzati per
sapere se denominare un taxon, possono essere utilizzati per sapere in quale categoria
tassonomica ubicarlo,[8] specialmente quello della stabilità nella nomenclatura.
I sistemi di classificazione che nascono come risultato della tassonomia hanno due
utilità:
Servono come contenitori di informazione. Gli scienziati di tutto il mondo
utilizzano i taxa come unità di lavoro, e ne pubblicano i risultati in relazione al
taxon studiato. Pertanto i nomi scientifici degli organismi sono la chiave di
accesso a un immenso corpo di informazione, disperso in molte lingue e
proveniente da molti campi della Biologia.
Permettono
di
fare predizioni riguardo
alla fisiologia, ecologia ed evoluzione dei taxa. Per esempio, è molto comune che
quando si trova un composto di interesse medico in una pianta, si investiga se
questo composto o altri similari si trovino anche in altre specie ad essa
imparentate.
I taxa o gruppi, nei quali vengono classificati gli esseri viventi, strutturati in una una
gerarchia di inclusione, nella quale un gruppo abbraccia altri minori ed è, a sua volta,
subordinato
a
uno
maggiore.
Ai
gruppi
viene
assegnato
un rango
tassonomico o categoria tassonomica che accompagna il nome proprio del gruppo.
Alcuni
esempi
conosciuti
sono: genere Homo,famiglia Canidae (canidi), ordine Primati, classe Mammalia (ma
mmiferi), regno Fungi (funghi).
Sono ranghi anche quelli di specie e sue subordinate. Il nome delle specie si distingue
da quelli dei taxa di altri ranghi perché va ad essere costituito di due parole, il che
rende ozioso scriverne la categoria.
Le categorie tassonomiche fondamentali si denominano, incominciando da quella che
più comprende (include):
Dominio
Regno
Phylum, (o Divisione, in questo caso viene esclusa la Zoologia)
Classe
Ordine
Famiglia
Tribù
Genere
Specie
Charles Darwin e le teorie evolutive
Charles Robert Darwin (Shrewsbury, 12 febbraio 1809 – Londra, 19 aprile 1882) è
stato unbiologo, geologo, zoologo e botanico britannico, celebre per aver formulato
la teoria dell'evoluzione delle specie animali e vegetali per selezione naturale agente
sulla variabilitàdei
caratteri
(origine
delle
specie),
e
per
aver
teorizzato
la discendenza di tutti i primati (uomo compreso) da un antenato comune (origine
dell'uomo). Pubblicò la sua teoria sull'evoluzione delle specie nel libro L'origine delle
specie (1859), che è rimasto il suo lavoro più noto. Raccolse molti dei dati su cui
basò la sua teoria durante un viaggio intorno al mondo sulla nave HMS Beagle, e in
particolare durante la sua sosta alle Isole Galápagos.
Con la teoria evoluzionistica Darwin dimostrò che l'evoluzione è l'elemento comune,
il filo conduttore della diversità della vita.Secondo una visione evolutiva della vita, i
membri dello stesso gruppo si assomigliano perché si sono evoluti da un antenato
comune.Secondo l'opinione di Darwin, le specie nascono mediante un processo di
“discendenza con variazione”. Fatto ancora più importante, nel suo trattato
sull'origine delle specie, Darwin oppose la teoria della selezione naturale per spiegare
con quali meccanismi avviene l'evoluzione.
La teoria evoluzionistica di Darwin si basa su tre presupposti fondamentali:
1. Riproduzione: tutti gli organismi viventi si riproducono con un ritmo tale che,
in breve tempo, il numero di individui di ogni specie potrebbe non essere più
in equilibrio con le risorse alimentari e l'ambiente messo loro a disposizione.
2. Mutazioni: all'interno della stessa specie ci sono esseri diversi tra loro; ve ne
sono di più lenti e di più veloci, di più chiari e di più scuri, e così via.
3. Selezione: esiste una lotta continua per la sopravvivenza all'interno della stessa
specie e anche all'esterno. Nella lotta sopravvivono gli individui più favoriti,
cioè quelli meglio strutturati per giungere alle risorse naturali messe loro a
disposizione, ottenendo un vantaggio riproduttivo sugli individui più deboli.
La selezione naturale avviene quando variazioni ereditabili vengono esposte a fattori
ambientali che favoriscono il processo riproduttivo di alcuni individui rispetto ad
altri. Egli affermò che l'evoluzione di nuove specie deriva da un accumulo graduale
di piccoli cambiamenti. Ciascuna specie presenta una propria serie di adattamenti,
ossia di caratteristiche che si sono evolute mediante la selezione naturale;
comprendere in che modo gli adattamenti si sono evoluti per selezione naturale è di
estrema importanza nello studio della vita quindi nella biologia.
I FOSSILI
Introduzione
Fossile In geologia, termine usato per indicare qualsiasi reperto fisico di organismo
animale o vegetale vissuto in tempi geologici passati e rimasto conservato all’interno
di formazioni rocciose sedimentarie. Il termine non indica soltanto parti di organismi,
quali possono essere ossa, denti o interi scheletri di animali, ma qualunque traccia
lasciata da antichi organismi: ad esempio, impronte sul terreno o escrementi. I fossili
sono oggetto di studio della paleontologia, che da essi trae le informazioni necessarie
per ricostruire la storia della vita sulla Terra.
La fossilizzazione
I processi di fossilizzazione sono una combinazione di trasformazioni biologiche,
chimiche e fisiche che agiscono su un organismo morto, una sua parte o una sua
traccia, alterandone la composizione e rendendolo chimicamente stabile.
Le prime trasformazioni che un organismo subisce dopo la morte sono quelle di
decomposizione biologica, che normalmente portano alla sua totale disgregazione.
Nel caso di un fossile, tali trasformazioni sono state evidentemente ostacolate dal
processo di sedimentazione in cui l’organismo si è trovato coinvolto. Se i sedimenti
sono sufficientemente fini, infatti, e se la loro velocità di deposizione è relativamente
rapida, l’organismo viene inglobato in un deposito compatto di sedimenti e in tal
modo protetto da qualunque fattore disgregante, di natura biologica o meccanica. In
questo senso, l’ambiente marino presenta le caratteristiche più adatte alla
fossilizzazione: gli animali morti che si depositano sul fondo possono contare su una
buona velocità di sedimentazione e su sedimenti sottili e compatti. Per questo, il
numero di fossili marini è nettamente superiore a quelli terrestri, e le informazioni di
cui i paleontologi dispongono sulla vita nei mari delle passate ere geologiche sono
più numerose di quelle relative ad altri ambienti.
La mineralizzazione
Una volta interrato, l’organismo morto attraversa la fase più significativa del
processo di fossilizzazione, quella chimica: le sostanze organiche di cui è composto
vengono via via sostituite da sostanze inorganiche, attraverso meccanismi di
mineralizzazione. Nella maggior parte dei casi, l’acqua che filtra attraverso i
sedimenti impregna l’organismo dei sali che porta disciolti in soluzione (calcare,
silice, fosfato di calcio). Più raramente, avviene una vera e propria sostituzione a
livello molecolare delle sostanze organiche di cui è costituito l’organismo con i sali;
il risultato di quest’ultima modalità di mineralizzazione (detta metasomizzazione) è
un fossile che conserva nei minimi dettagli le forme dell’organismo originario, ma
che presenta una composizione chimica completamente diversa da quella originaria.
Altri meccanismi di fossilizzazione
Nel caso specifico dei resti vegetali, una delle modalità possibili di fossilizzazione è
la carbonificazione: in seguito all’attacco di batteri specifici, il materiale vegetale si
impoverisce di ossigeno e azoto e quindi aumenta la propria concentrazione relativa
di carbonio; da qui la formazione dei giacimenti di carbon fossile, che altro non sono
se non resti di piante fossili.
Un altro meccanismo di fossilizzazione interessa resti organici venutisi a trovare in
vicinanza di una sorgente o di un bacino di acqua calcarea: il carbonato di calcio
contenuto nell’acqua a poco a poco si deposita sul resto organico incrostandolo
completamente; ne rimane un calco che riproduce fedelmente la forma
dell’organismo originario.
Casi particolari di fossilizzazione, in cui si conservano non solo le parti dure
dell’organismo morto, ma anche i suoi tessuti molli, possono avvenire quando
l’inglobamento in un mezzo avviene prima che abbia inizio la decomposizione o
intervenga qualunque altro fattore disgregante. È il caso dei fossili di insetti, di
polline o di altri piccoli animali rimasti invischiati nella resina, successivamente
trasformata in ambra, o dei giganteschi mammut perfettamente conservati dai ghiacci
del loro antico habitat.
Classificazione e tipi di fossili
La classificazione dei fossili segue fedelmente i criteri della classificazione
tassonomica dei viventi. Ogni resto viene quindi identificato in base alle stesse
categorie sistematiche degli organismi che popolano oggi il pianeta: specie, genere,
famiglia, ordine, classe, phylum e regno.
La maggior parte dei fossili che si rinvengono consiste di parti della struttura
originaria dell’organismo, ad esempio ossa, che hanno subito un processo di
mineralizzazione. Numerosi sono anche gli stampi e i calchi naturali che si formano
quando anche le strutture coriacee di un organismo (scheletro, conchiglie, gusci)
vengono disciolte dalla circolazione di acqua: le cavità che si producono in seguito
alla dissoluzione vengono riempite da sedimenti che, consolidandosi, formano
repliche degli originali. Altri tipi di fossili sono costituiti da impronte di animali; da
animali interi rimasti custoditi in suoli gelati o in torbiere; dai coproliti – escrementi
fossili che, se possono essere attribuiti con sicurezza a un dato animale, talvolta
consentono di risalire al suo tipo di dieta.
Importanza dei fossili
I fossili costituiscono una fonte preziosa di informazioni sugli ecosistemi che si sono
succeduti nella storia della Terra: da essi è possibile risalire alle caratteristiche delle
forme viventi del passato e degli ambienti in cui queste sono fiorite.
Va tuttavia tenuto presente che le informazioni fornite dai fossili sono parziali, in
quanto riguardano soltanto alcune tipologie di organismi, e in particolare quelli che,
dotati di parti dure come un guscio, una conchiglia o uno scheletro, non si sono
completamente disgregati prima dell’inizio del processo di fossilizzazione. Così,
benché non si possa escludere che organismi pluricellulari popolassero i mari del
Precambriano, non si dispone di resti fossili che permettano di accertarne l’esistenza:
essendo a corpo molle, questi organismi non avrebbero comunque potuto lasciare
traccia di sé. La documentazione fossile diviene enormemente ricca solo a partire da
570 milioni di anni fa, quando si affermarono i primi animali dotati di conchiglie e di
parti scheletriche.
Fossili e teoria dell’evoluzione
L’importanza dei fossili risiede anche nel ruolo determinante che hanno avuto nella
nascita e nell’affermazione della teoria dell’evoluzione di Darwin. Prima che essi
venissero scoperti e riconosciuti come resti di organismi vissuti in epoche passate,
infatti, si riteneva che il mondo fosse sempre stato popolato dalle stesse forme
viventi.
Fossili e stratigrafia
In campo geologico, e più propriamente stratigrafico, i fossili costituiscono un
utilissimo sistema di datazione delle successioni di strati sedimentari, dette serie
stratigrafiche: la presenza dei fossili al loro interno permette infatti di stabilire l’età
relativa delle formazioni sedimentarie, le une rispetto alle altre. In questo tipo di
datazione sono particolarmente utili i cosiddetti fossili guida: si tratta di particolari
organismi che, al tempo in cui sono vissuti, hanno goduto di una straordinaria
diffusione geografica in un periodo di tempo limitato; queste caratteristiche fanno sì
che essi si trovino oggi all’interno di strati rocciosi ben definiti, in diverse zone del
globo.
Basandosi sull’analisi dei fossili, i geologi e i paleontologi, a partire dal XIX secolo,
hanno quindi potuto stabilire una cronologia dell’evoluzione delle forme di vita sulla
Terra nei suoi ultimi 500 milioni di anni di vita, dal Cambriano all’Olocene.
CARBON FOSSILE
Il Carbon fossile è un Combustibile solido di origine vegetale. Nelle passate ere
geologiche, in particolare nel Carbonifero (circa 300 milioni di anni fa), gran parte
della superficie terrestre era occupata da paludi in cui cresceva una vegetazione
lussureggiante che comprendeva molte varietà di felci, alcune grandi come alberi.
Man mano che morivano, le piante venivano sommerse dall'acqua: la materia
organica dunque non si decomponeva, ma cominciava a subire un lento processo di
carbonizzazione, una particolare forma di fossilizzazione consistente nella perdita
graduale e continua di atomi di idrogeno e di ossigeno, con il conseguente accumulo
di un'alta percentuale di carbonio.
In tal modo si formarono i primi giacimenti di torba, ricoperti col passare del tempo
da strati di terreno più o meno spessi. In migliaia e milioni di anni la pressione degli
strati sovrastanti, i sommovimenti della crosta terrestre e, talvolta, il calore dei
vulcani compressero e compattarono gli originari depositi di torba, trasformandoli
progressivamente in carbone.
TIPI DI MINIERE
I diversi tipi di carbon fossile vengono classificati secondo la loro età, e quindi
secondo il loro contenuto percentuale di carbonio. La torba, che rappresenta il primo
stadio della carbonizzazione, ha un basso contenuto di carbonio e un alto grado di
umidità. Il contenuto di carbonio è maggiore nella lignite, che costituisce lo stadio
immediatamente precedente il carbon fossile vero e proprio, rappresentato dal
litantrace, che contiene ancor più carbonio, e che quindi ha un potere calorifico
relativamente alto, e dall'antracite, che ha il massimo contenuto di carbonio e il potere
calorifico maggiore.
Se sottoposto a pressione e calore ulteriori, il carbon fossile può trasformarsi in
grafite, che è praticamente carbonio puro. Altri componenti del carbon fossile sono
alcuni idrocarburi volatili, zolfo e azoto, oltre ai minerali che residuano dalla
combustione sotto forma di cenere. Alcuni dei prodotti di combustione del carbon
fossile hanno effetti nocivi sull'ambiente, come, ad esempio, il diossido di carbonio o
anidride carbonica (CO2).
Alcuni scienziati ritengono che, a causa dell'uso generalizzato di carbone e altri
combustibili fossili, la quantità di diossido di carbonio nell'atmosfera terrestre possa
aumentare tanto da influenzare il clima del pianeta (vedi Effetto serra).
Durante la combustione, inoltre, zolfo e azoto contenuti nei combustibili fossili
formano ossidi che contribuiscono alla formazione di piogge acide, risultato di una
complessa serie di reazioni che coinvolgono sostanze chimiche di varia provenienza,
sia naturali sia prodotte da attività industriali o dai gas di scarico dei mezzi di
trasporto con motore a combustione interna. In molte nazioni, le emissioni di diossido
di zolfo o anidride solforosa (SO2) provenienti dalle moderne centrali termoelettriche
a carbone sono state poste sotto controllo, riuscendo a ottenerne la diminuzione,
malgrado l'aumento dell'impiego di carbone.
Ogni tipo di carbon fossile ha il suo valore economico. La torba è stata usata per
secoli, nelle miniere di carbone, per produrre il cosiddetto fuoco in cantiere, e
attualmente torba e lignite servono ad alimentare i forni, dopo essere state compresse
in mattonelle. Le centrali termoelettriche e varie industrie utilizzano come
combustibile il litantrace, mentre l'industria siderurgica fa largo uso di coke
metallurgico, un combustibile ad altissima percentuale di carbonio, che si ottiene
dalla distillazione del carbon fossile. Dall'inizio del XIX secolo alla seconda guerra
mondiale il carbon fossile fu usato anche per la produzione di gas combustibili e di
idrocarburi liquidi.
Questa produzione è molto diminuita da quando è cominciato lo sfruttamento delle
enormi riserve di gas naturale, benché negli anni Ottanta si sia risvegliato l'interesse
delle nazioni industrializzate verso questi processi, che rientrano fra le tecnologie
innovative del 'carbone pulito'. Si può citare l'esempio del Sudafrica, dove l'intero
fabbisogno di olio combustibile è fornito dalla liquefazione del carbon fossile.
TECNOLOGIE DEL CARBONE PULITO
Si tratta di una nuova generazione di processi di utilizzazione del carbon fossile,
alcuni dei quali potranno essere sfruttati commercialmente all'inizio del XXI secolo.
Le tecnologie del carbone pulito sono diverse, ma tutte accomunate dal principio di
riuscire ad alterare la struttura di base del carbone, prima o durante una delle fasi di
trattamento o di utilizzo.
In questo modo sarebbe possibile ridurre l'emissione di impurità, quali ossidi di zolfo
e di azoto, durante il processo di combustione, e aumentare la resa energetica. Queste
tecniche innovative comprendono raffinati metodi di pulizia del carbone, di
combustione in letto fluido, di ciclo combinato di gasificazione integrata, di totale
desolforazione dei gas di combustione.
GLI OGM
Gli organismi geneticamente modificati (OGM), sono organismi caratterizzati da un
patrimonio genetico (genoma) alterato rispetto a quello tipico della propria specie,
per l’introduzione artificiale di uno o più geni provenienti da altri organismi
Per ottenere organismi transgenici si utilizzano le tecniche dell’ingegneria genetica. Il
frammento di DNA in cui si trova il gene da inserire viene iniettato in una cellula
batterica, o in una cellula uovo (che verrà successivamente fecondata) o in un
embrione. Per potere essere attivo, il frammento di DNA deve essere associato a un
vettore d’espressione, ossia a un’altra porzione di DNA specifica che controlla le
modalità di espressione del gene da trasferire; ad esempio, esso permette che il gene
si esprima (cioè svolga la propria attività) soltanto in determinati tessuti.
Il DNA estraneo viene inoculato per microiniezione nella cellula ricevente; dopo
l’inoculazione, il nuovo gene si integra con il DNA di questa, e può di conseguenza
venire trasmesso a tutte le cellule che derivano per successive mitosi dalla cellula
ricevente.
Nel caso si utilizzino embrioni, i frammenti di DNA contenenti i geni possono essere
anche inseriti tramite un virus-vettore, ossia tramite un virus infettivo nel quale, a sua
volta, è stato inoculato il frammento di DNA. Si calcola che la percentuale di
successo di questa tecnica, che si traduce con il numero di organismi transgenici
vitali e nei quali i geni estranei sono funzionanti, sia dell’1%.
Il controllo dell’avvenuta integrazione del gene nel patrimonio genetico
dell’organismo ricevente può essere fatto prelevando da alcune cellule transgeniche
campioni di DNA ed esaminandoli, in genere mediante la tecnica nota come reazione
a catena della polimerasi (PCR).
L’IMPIEGO DEGLI ORGANISMI TRANSGENICI
Nella ricerca biologica e genetica, l’impiego di organismi transgenici è rilevante
nell’ambito degli studi sulla funzione di geni specifici; infatti, l’immissione di un
gene estraneo in un organismo determina l’insorgenza in questo di particolari
caratteristiche (come la resistenza a un erbicida o la capacità di sintetizzare una data
proteina) che, confrontate con quelle degli individui della stessa specie, permettono la
comprensione del ruolo di quel gene.
A scopo di ricerca, sono impiegati anche particolari tipi di organismi transgenici, i
cosiddetti knock-out, in cui un gene dell’organismo viene eliminato o inattivato;
alcuni topi così modificati, ad esempio, sono stati utilizzati per studiare il ruolo
funzionale di alcuni geni specifici nello sviluppo embrionale. Disattivando in animali
da laboratorio il gene corrispondente a un gene non funzionale nei pazienti affetti da
una particolare malattia, si possono creare modelli utili a fini diagnostici e terapeutici.
UNA QUESTIONE CONTROVERSA
L’impiego di organismi geneticamente modificati è uno dei più dibattuti temi della
bioetica. Infatti, già da tempo la creazione di nuove culture vegetali o di
microrganismi modificati può essere siglata da brevetto; la possibilità di estendere
questa pratica anche a organismi più complessi, e ai procedimenti industriali che ne
permettono l’ottenimento, suscita attualmente atteggiamenti diversi: da un lato
entusiasmo, per le nuove prospettive economiche e scientifiche che potrebbero
derivarne; dall’altro, preoccupazione, per tutte le implicazioni, soprattutto etiche e
sociali.
Si ritiene che la questione dell’impiego delle specie transgeniche non debba limitarsi
a un’analisi dei costi e dei benefici economici, e che le attuali leggi sui brevetti,
relative a strumentazioni, non possano essere semplicemente estese a organismi
viventi. Sono inoltre oggetto di discussione le possibili conseguenze sulla biodiversità
e sugli equilibri degli ecosistemi dell’immissione nell’ambiente di organismi
modificati, con caratteri che potrebbero venire trasmessi alla discendenza; inoltre,
suscitano perplessità i possibili effetti a lungo termine sulla salute umana del
consumo di prodotti derivanti da organismi geneticamente modificati.
L’ACCIAIO
Nella seconda metà dell’800 si capì che il carbonio era la causa della fragilità delle
ghise, perciò si studiò il modo di ridurne la percentuale, senza rinunciare però alla
durezza. si sperimentò cosi’, l’aggiunta di altri metalli in lega.
L’acciaio si ottiene per CONVERSIONE DELLA GHISA, entro appositi forni a
1600°C (forni CONVERTITORI) dove l’INSUFFLAGGIO di OSSIGENO e l’uso di
rottami vecchi ricchi di ruggine (ossidi capaci di liberare ossigeno) sono in grado di
ridurre il carbonio sotto l’1,7%.
ACCIAI ATTUALMENTE IN COMMERCIO
ACCIAI COMUNI O AL CARBONIO
acciai tradizionali con tracce di silicio e manganese. La durezza cresce al crescere
della percentuale di carbonio, mentre la tenacità ne è inversamente proporzionale.
(extradolce, dolce, semiduro, duro, extraduro)
ACCIAI SPECIALI
sono simili agli acciai comuni, ma con un drastico controllo della percentuale di
fosforo e zolfo, che non devono superare lo 0,035%. Ne risultano migliorate tutte le
prestazioni.
ACCIAI LEGATI
Oltre che col carbonio, entrano in lega altri metalli, quali, silicio, manganese, cromo,
nichel o rame, che ne migliorano le qualità.
trattamenti termici
TEMPRA
consiste nel portare il metallo a 800-900°C e poi raffreddarlo rapidamente. In tal
modo i cristalli non fanno in tempo a formarsi e restano piccoli e incompleti. La
superficie esterna è quella che estremizza il fenomeno in quanto è la prima a
raffreddarsi. Essa risulta ricca di legami distorti che le donano una forte durezza.
Eccessiva durezza può però causare alta fragilità, non bisogna quindi eccedere.
RINVENIMENTO
serve per eliminare l’eccessiva fragilità derivante dalla tempra, si tratta di ririscaldare
il metallo fino a 500-600°C e di farlo raffreddare lentamente per recuperare un po'
della tenacità persa in durezza.
BONIFICA
Operazione simile al rinvenimento che si attua però su metalli lavorati in lamiere e a
freddo, al fine di renderli meno soggetti alla frantumazione.
CEMENTAZIONE
Arricchimento superficiale di carbonio, al fine di ottenere una superficie esterna dura
e un interno tenace.
RICOTTURA
Cancella tutti i trattamenti subiti. La temperatura è portata oltre 900°C e il metallo
viene lasciato raffreddare lentamente.
PROPRIETA CHIMICO FISICHE
MALLEABILITA’
Capacità di lasciarsi lavorare al laminatoio, un apparecchiatura formata da cilindri
accoppiati rotanti in senso opposto attraverso i quali viene fatto passare un lingotto
rovente.
DUTTILITA’
Capacità di lasciarsi lavorare per trafilatura. Il materiale viene fatto passare attraverso
un orifizio calibrato e viene “passato” e trasformato in fili.
ALLIGABILITA’
Possibilità del metallo di legarsi con altri. E’ ormai impensabile usare del metallo
puro, tranne che per il rame da impiantistica elettrica.
SALDABILITA’
Possibilità del materiale di essere unito mediante fusione parziale o colatura di
materiale fuso tra due parti solide.
DUREZZA
Capacità di resistere a penetrazione, scalfittura, usura per strofinio. si accompagna
purtroppo alla fragilità.
TENACITA’
Semplicisticamente è definita la capacità di resistere agli urti, o meglio “la capacità
del metallo di assorbire l’energia di una sollecitazione dinamica”.
PASSIVABILITA’
Capacità di resistere all’aggressione degli agenti chimici presenti nell’atmosfera.
I METALLI
I Metalli sono Elementi chimici caratterizzati in generale da lucentezza, durezza,
malleabilità, duttilità, buona conducibilità elettrica e termica; a temperatura ambiente
sono solidi cristallini (con l’eccezione del mercurio). I metalli (fra i quali, ad
esempio, alluminio, argento, bario, berillio, bismuto, cadmio, calcio, cerio, cobalto,
cromo, ferro, iridio, litio, magnesio, manganese, mercurio, molibdeno, nichel, oro,
osmio, palladio, piombo, platino, potassio, radio, rame, rodio, sodio, stagno, tallio,
tantalio, titanio, torio, tungsteno, uranio, vanadio, zinco) possono combinarsi fra loro
e con alcuni altri elementi, formando composti, soluzioni solide o miscele.
Una sostanza costituita da due o più metalli, oppure da un metallo e da particolari non
metalli, come ad esempio il carbonio, si chiama lega. Le leghe formate da mercurio e
altri elementi metallici sono invece dette amalgami.
Nell’ambito delle definizioni generali che permettono di classificare un elemento tra i
metalli, le proprietà di ogni singola sostanza possono essere molto variabili. La
maggior parte degli elementi metallici ha colore grigiastro, ma il bismuto è rossastro,
il rame è rosso e l’oro è giallo. In alcuni casi, il colore varia in rapporto alla direzione
di assorbimento della luce, un fenomeno noto come pleocroismo.
I punti di fusione possono variare da un minimo di -39 °C per il mercurio a un
massimo di 3410 °C per il tungsteno. L’iridio (che ha densità relativa pari a 22,4) è il
metallo più denso e il litio (che ha densità relativa di 1,53) è il meno denso. La
maggior parte dei metalli cristallizza nel sistema cubico, ma non sono rare celle
esagonali o tetragonali (vedi Cristallo). A temperature ordinarie, il bismuto presenta i
valori più bassi di conducibilità elettrica, mentre l’argento presenta quelli più alti (per
quanto riguarda le basse temperature, vedi Criogenia e Superconduttività). La
conducibilità della maggior parte degli elementi metallici può essere ridotta
trasformandoli in leghe. Tutti i metalli si dilatano se scaldati e diminuiscono di
volume se raffreddati; alcune leghe, ad esempio quelle di platino e di iridio, hanno
coefficiente di dilatazione termica molto basso.
Proprietà fisiche
I metalli sono in genere molto duri e resistenti a diversi tipi di sforzi. Nonostante le
differenze fra un metallo e l’altro siano notevoli, la loro classificazione avviene
esclusivamente in base a proprietà fisiche quali: la durezza (cioè la resistenza a
deformazioni superficiali o abrasioni); la resistenza a trazione (cioè la resistenza alla
rottura); l’elasticità (cioè la capacità di assumere la forma originaria quando cessa la
deformazione); la malleabilità (cioè la capacità di essere ridotti in lamine sottili senza
subire rotture); la resistenza alla fatica (cioè la capacità di resistere a sforzi ripetuti);
la duttilità (cioè la capacità di venire deformati senza subire rotture). Vedi anche
Scienza e tecnologia dei materiali.
Proprietà chimiche
Nella maggior parte dei composti chimici, i metalli hanno stati di ossidazione
positivi, cioè cedono con facilità uno o due elettroni agli atomi a cui si legano. Sono
inoltre caratterizzati da potenziali di ionizzazione relativamente bassi e possono
facilmente trasformarsi in ioni positivi (cationi). Di conseguenza formano ossidi di
tipo basico e possono formare sali, come cloruri, solfuri e carbonati. Vedi Reazione
chimica.
Struttura elettronica
Le varie caratteristiche dei metalli, in particolare l’elevata conducibilità termica ed
elettrica, possono essere spiegate sulla base della struttura chimica ed elettronica di
questi elementi.
Secondo la teoria universalmente accettata, nota come teoria delle bande, il legame
tra atomi di un metallo è dovuto alla sovrapposizione degli orbitali atomici.
Interagiscono fra loro solo gli orbitali di energia simile, quindi da ogni n orbitali
atomici di energia equivalente, si formano n orbitali molecolari, estesi a tutto il
solido, disposti su livelli energetici molto ravvicinati. A ciascuno di questi gruppi di
orbitali molecolari si dà il nome di banda.
Le bande sono fra loro separate da salti energetici, detti gap, la cui ampiezza dipende
dalla differenza di energia fra i gruppi di orbitali atomici da cui sono state formate.
Gli elettroni di valenza di ogni atomo del metallo si dispongono, a coppie, su tutti gli
orbitali molecolari di ogni banda, a partire da quelli a energia più bassa. Di solito, i
metalli non hanno elettroni in numero sufficiente per popolare tutti gli orbitali
molecolari, quindi le bande a energia più alta sono vuote o solo parzialmente
occupate. Assorbendo energia dall’esterno, gli elettroni dei livelli più bassi possono
spostarsi in quelli più alti, vuoti: tali spostamenti giustificano le proprietà tipiche dei
metalli.