Le basi
di ACER
Sintesi dei principali
concetti di arboricoltura
A cura di Alessio Fini, Dipartimento di Ortoflorofrutticoltura, Università degli Studi di Firenze
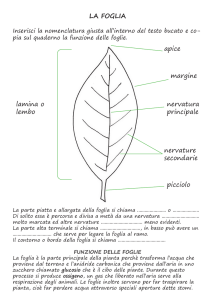
LA STRUTTURA DELLE FOGLIE
L’interno della foglia
105 • ACER 5/2007
Effetti dell’ombra
sulle foglie
A
B
C
D
E
F
A. Fini e F. Ferrini
T
Diverse tipologie di foglie; semplici: A= ovata a margine intero
(Berberis Thunbergii ‘Golden Ring’); B= cordata a margine
dentato (Davidia involucrata); C= palmata a margine seghettato
(Acer pseudoplatanus ‘Brillantissimum’); D= lobata (Quercus
robur ‘Piramidalis’); composte: E= palmato-composta (Aesculus
x carnea); F= pennata (Fraxinus oxycarpa).
l’età della foglia e l’aumentare
dall’umidità relativa dell’aria (3).
Koch et al. (7), per spiegare la
variabilità della deposizione
delle cere in funzione dell’umidità relativa, hanno ipotizzato un
meccanismo di cotrasporto dell’acqua e delle cere dall’epidermide fino alla superficie esterna
della cuticola. La cuticola serve
principalmente per limitare le perdite non stomatiche di acqua, ma
anche per proteggere i tessuti sottostanti dalle radiazioni UV, dal
surriscaldamento eccessivo e
dall’ingresso di parassiti (3).
La cuticola può essere attraversata per diffusione da sostanze
lipofile, mentre ioni e composti
polari la possono attraversare solo
in corrispondenza di specifici
canali, di cui ancora si sa poco.
• Epidermide. È il tessuto tegumentale di origine primaria che
riveste le foglie, proteggendo i
tessuti sottostanti ed è generalmente costituito da un solo strato
di cellule appiattite di grandi
dimensioni, saldamente attaccate
l’una all’altra. Le cellule epidermiche sono normalmente prive di
clorofilla e di cloroplasti ben differenziati, non fanno fotosintesi e
risultano semitrasparenti alla PAR
(radiazione fotosinteticamente attiva), consentendo alla luce di giungere ai tessuti sottostanti. In alcune specie l’epidermide è provvista
di peli che, se abbondanti, contribuiscono a creare uno strato di
vapore acqueo che riduce le perdite d’acqua per traspirazione.
L’epidermide ospita anche gli stomi: localizzati nella pagina inferiore nelle foglie ipostomatiche,
comuni nelle dicotiledoni; in
entrambe le pagine fogliari nelle
foglie anfistomatiche, comuni nelle monocotiledoni; oppure esclusivamente nella pagina supe-
▼
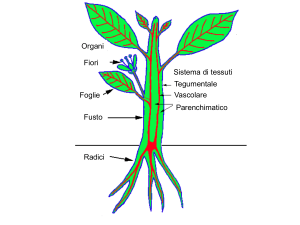
Le foglie sono il principale organo fotosintetizzante della pianta e
svolgono un ruolo primario nella
strategia adattativa dei vegetali (6).
La grande diversità riscontrabile
nella morfologia fogliare delle
diverse specie vegetali è basata
su fattori genetici e su adattamenti evolutivi che le diverse specie
hanno sviluppato in risposta alla
pressione ambientale. A prescindere da queste diversità, le foglie
presentano tessuti e strutture ricorrenti, le cui dimensioni e distribuzione relativa nella lamina fogliare
sono fondamentali per determinare e influenzare l’assimilazione del
carbonio atmosferico.
Le foglie devono massimizzare
l’assimilazione del carbonio atmosferico, limitando nel contempo le
perdite di acqua per traspirazione. Per questo hanno sviluppato
strutture altamente specializzate.
Analizzando la sezione di una
foglia di una pianta dicotiledone si
notano, dalla pagina superiore
verso quella inferiore, cuticola, epidermide, mesofillo (composto dai
tessuti a palizzata, lacunoso e dai
fasci conduttori), epidermide inferiore e cuticola.
• Cuticola. La cuticola è una
membrana extracellulare continua
secreta dall’epidermide e composta da lipidi polimerizzati (principalmente cutina e, in alcune specie, cutano) in cui sono immerse
delle cere, dette intracuticolari.
Sulla superficie esterna si ritrovano altre cere, dette epicuticolari, la
cui quantità è determinata sia dalla specie, sia dalle condizioni
ambientali. In generale, la quantità di cere depositate aumenta
con l’intensità luminosa e con la
temperatura e diminuisce con
ra i vari organi della pianta,
le foglie risentono maggiormente della variabilità ambientale che può modificarne
l’anatomia e le caratteristiche
funzionali. Tra le variabili ambientali, la luce è in grado di indurre profonde alterazioni sulla
morfologia e sulla struttura fogliare. Recenti studi hanno evidenziato come la variazione
dell’intensità luminosa incidente sulla lamina fogliare modifichi le caratteristiche morfologiche e fisiologiche in funzione
del genotipo delle specie vegetali considerate (5, 1). Tali studi
evidenziano come la diminuzione della radiazione incidente sulla lamina fogliare provochi un aumento della superficie
media delle foglie. Parallelamente si assiste a un calo dello spessore medio della foglia,
così come nei tessuti interni
(cuticola, epidermide, tessuto
a palizzata e lacunoso), mentre
la dimensione media delle cellule aumenta. Il numero medio
di strati del tessuto a palizzata
diminuisce poiché la foglia cerca di ridurre l’ombreggiamento
intrafogliare. Il numero medio di
stomi per unità di superficie diminuisce al diminuire della radiazione incidente. Queste alterazioni anatomiche fanno sì
che le foglie d’ombra abbiano
bassi punti di compensazione
e saturazione luminosa, ovvero riescono a fare fotosintesi in
modo efficiente a basse intensità luminose ma, in presenza
di elevata radiazione, non raggiungono livelli di assimilazione tanto alti quanto le foglie
adattate alla luce solare piena.
Palizzata
M
e
s
o
f
i
l
l
o
Fascio conduttore
Epidermide inferiore
Lacunoso
Sezione longitudinale di una foglia dorso ventrale di Photinia x
fraseri fotografata al microscopio ottico a 100 ingrandimenti.
▼
riore nelle foglie epistomatiche,
esclusive di alcune piante acquatiche. Gli stomi sono strutturati in
modo da potersi aprire in risposta
allo stimolo luminoso, permettendo l’entrata della C02 necessaria
per la fotosintesi e chiudersi al buio
o quando la pianta si trova in stato
di stress, soprattutto idrico, al fine
di limitare la perdita di acqua. Uno
stoma è formato da due cellule
affiancate dette di guardia che
delimitano una rima stomatica, che
conduce alla camera sottostomatica, uno spazio intercellulare in
comunicazione con gli spazi intercellulari del mesofillo.
• Mesofillo. Comprende tutti i tessuti inclusi tra l’epidermide superiore e quella inferiore ed è formato da tessuti parenchimatici (a
palizzata e lacunoso) nei quali
decorrono i fasci conduttori (2). Il
parenchima a palizzata è il principale tessuto fotosintetizzante della foglia e il più ricco in clorofilla. È
costituito da cellule allungate,
orientate perpendicolarmente
rispetto all’epidermide e appressate le une alle altre, che formano
degli strati il cui numero è variabile in funzione del genotipo e del-
le condizioni ambientali (8). Un elevato numero di strati di palizzata
è utile al fine di incrementare il
punto di saturazione luminosa in
presenza di molta radiazione
(vedi box pag. 105) ma, in
ambienti ombreggiati, non è una
strategia vantaggiosa in quanto
riduce l’efficienza fotosintetica per
unità di azoto fogliare (9). Sotto il
palizzata si trova il tessuto lacunoso, caratterizzato da cellule di
forma irregolare separate da
ampi spazi intercellulari. Sebbene siano presenti numerosi cloroplasti, la principale funzione di
questo tessuto è assicurare un
ricambio d’aria sufficiente per
consentire un efficiente livello di
fotosintesi. Il mesofillo ospita i
fasci conduttori, che derivano
direttamente da quelli del fusto e
si ramificano nelle foglie formando le nervature. Queste possono
essere retinervie (dicotiledoni),
quando da una nervatura principale si dipartono e ramificano
quelle secondarie, oppure parallelinervie (monocotiledoni), se le
nervature decorrono più o meno
parallele, senza mai intersecarsi.
Un fascio conduttore, al suo inter-
Notizie curiose
• Le foglie degli alberi, soprattutto se tomentose, molto fitte o persistenti anche in inverno, sono estremamente efficaci nel fissare gli
inquinanti gassosi e il particolato. Ogni albero sequestra mediamente 0,024 kg/anno di inquinanti quali ozono, diossido di azoto e
di zolfo, monossido di carbonio (10).
• Un faggio adulto, di 25 m di altezza, ha una superficie fogliare di
circa 1600 m2: per avere la medesima superficie assimilante sono
necessari circa 600 giovani alberi (4).
• Una foglia completamente espansa di camelia esposta alla luce
solare piena è spessa circa 200 µ; in condizioni di forte ombreggiamento tale spessore si riduce a 150 µ.
A destra,
particolare delle cere
depositate sulla
pagina inferiore
di una foglia
di viburno,
fotografata al
microscopio
elettronico
(ESEM) a 1200
ingrandimenti.
Sotto, particolare
della pagina inferiore
di una foglia
ipostomatica
di fotinia fotografata
al microscopio
elettronico
(ESEM) a 600
ingrandimenti.
no, presenta una porzione xilematica e una floematica, racchiuse da una serie di cellule modificate dette guaina del fascio.
Altre tipologie di foglie
Quanto riportato finora vale per
le foglie dorso-ventrali, tipiche delle dicotiledoni. Strutture differenti
possono essere ritrovate in altre
divisioni o classi. Le monocotiledoni hanno foglie ortotrope, in cui
il mesofillo non è distinto in palizzata e lacunoso. Anche nelle
foglie di conifere non è evidente
alcuna distinzione all’interno del
mesofillo che, inoltre, risulta compatto e poco valido per effettuare
gli scambi gassosi. In alcune specie le foglie possono subire modificazioni morfologiche tali da comportare la riduzione o la perdita
totale della funzione fotosintetica e
l’assunzione di funzioni alternative
(protezione, vessillare). È il caso
delle foglie trasformate in spine o
delle brattee fiorali.
Bibliografia
1) Amoroso G., Ferrini F., Frangi
P., Ferrante A., Fini A., Piatti
R., 2007. Crescita e caratteristiche fisiologiche di arbusti
allevati in condizioni di diversa intensità luminosa. Italus
Hortus, 14: 95.
2) Arrigoni. O. 1986. Elementi di
biologia vegetale – botanica
generale. Casa Editrice Ambrosiana, Milano. 541 pp.
3) Brine K., Schumacher C.F.A.,
Schimtz-Eilberger M., Steiner
U., Oerke E.C., 2006. Ontogenic variation in chemical
and physical characteristics
of adaxial apple surface. Phytochemistry, 67:161-170.
A. Fini e C. Giordano
Epidermide superiore
A. Fini e C. Giordano
Cuticola
A. Fini e C. Giordano
Le basi
di ACER
4) Chiusoli A., 1999. La scienza
del paesaggio. Cleb (Ed.),
Bologna, pp. 395.
5) Fini A., Amoroso G., Ferrini F.,
Frangi P., 2006. Effetto dell’ombreggiamento sulla crescita di tre specie ornamentali. Lineaverde, aprile.
6) Fleming A.J., 2005. The control of leaf development. New
Phytologist, 166: 9-20.
7) Koch K., Hartmann K.D., Schreiber L., Barthlott W.,
Neinhuis C., 2006. Influence
of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Environmental and experimental botany, 56:1-9.
8) Lee D.W., Oberbauer S.F.,
Johnson P., Krishnapilay B.,
Mansor M., Mahamad H.,
Yap S.K., 2000. Effect of irradiance and spectral quality
on leaf structure and function in seedlings of two
southeast asian Hopea (Dipterocariaceae) species .
American Journal of Botany,
87(4): 447-455.
9) Reich P.B., Ellsworth D.S., Walters M.B., 1998. Leaf structure
(specific leaf area) modulates
photosynthesis-nitrogen relations: evidence from within
and across species and functional groups. Functional Ecology, 12: 948-958.
10) Thompson J.R., Novak D.J.,
Crane D.A., Hunkins J.A.,
2004. Iowa, U.S., communities benefit from a tree-planting program: characteristics of newly planted trees.
Jour nal of Arboriculture,
30(1): 1-10.
ACER 5/2007 • 106