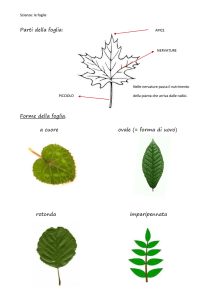
La foglia
Struttura e funzioni
La foglia è un organo complesso costituisce un’appendice laterale
del fusto e dei rami.
Colore verde e forma appiattita più o meno espansa o cilindrica più
o meno allungata consentono l’esposizione alla luce e continui
scambi gassosi con l’atmosfera.
Funzioni principali(nomofilli):
Fotosintesi
Traspirazione
Funzioni secondarie di alcune:
Protezione (brattee)
Richiamo per i pronubi(ipsofilli)
Sostegno (cirri o viticci)
Difesa da erbivori(spine)
Immagazzinamento(catafilli, cotiledoni embriofilli)
Assunzione sostanze azotate(ingerendo insetti)
Tipi basilari di stele nelle piante vascolari.
(A) Protostele, (B) sifonostele, (C-D) eustele
atactostele.
La struttura primaria del fusto è anche condizionata
dalle foglie.
L’articolazione del fusto in nodi e internodi dipende
ovviamente dalla presenza delle foglie, la
determinazione dei cordoni procambiali e, il loro
differenziamento, dalla genesi delle bozze fogliari.
La stessa distribuzione dei fasci nel cilindro centrale
dipende dalle foglie.
La struttura eustelica di Gimnosperme e Dicotiledoni,
con relativamente pochi fasci, è in rapporto con il
tipo di nervature delle foglie: una sola e non
ramificata nelle Gimnosperme, ancora una, anche
se poi ramificata, nelle Dicotiledoni, nervatura che
comunque contiene sempre uno o pochi fasci.
La struttura atactostelica delle Monocotiledoni, con
decine di fasci, di differenti dimensioni e che
occupano tutto il cilindro centrale, dipende dalle
nervature parallelinervie delle foglie, ognuna delle
quali contiene molti fasci, più grossi al centro, più
piccoli lateralmente .
Ancora dalla foglia, o meglio dalla necessità di
produrne continuamente di nuove, dipende quella
profonda modificazione della struttura di fusto e
radice che va sotto il nome di “struttura
secondaria”.
La foglia è un organo ad accrescimento definito e, breve è la
durata della sua vita.
Nei casi più frequenti, una stagione vegetativa.
In una pianta pluriennale, ogni anno deve essere prodotta
una nuova popolazione di foglie.
A questo provvede il meristema apicale del fusto ( o meglio,
tutti i meristemi di tutte le ramificazioni del fusto), la cui
funzione principale è appunto quella di produrre foglie
e, alla loro ascella , gemme che ramificheranno il fusto e
porteranno altre foglie.
Una gemma che cresce, fa diventare foglie le sue bozze,
distende i suoi internodi (nei quali si troveranno i fasci
conduttori derivanti dalle foglie stesse) e prolunga
longitudinalmente il fusto precedente, nel quale sono
presenti i tessuti conduttori che servivano la
generazione precedente di foglie.
In questa zona di fusto dovranno essere inseriti nuovi
tessuti conduttori che, per così dire, colleghino fino alle
radici i fasci conduttori delle nuove foglie.
La forma della foglia è in relazione con la funzione
fotosintetica da svolgere:
Ampia superficie
Spessore ridotto
Per questo hanno sviluppato una struttura laminare: il
lembo fogliare.
Deve sussistere un giusto apporto di acqua ed anidride
carbonica assicurato dalla presenza, al suo interno, dei
tessuti conduttori
Carichi di acqua provenendo dal fusto, si ramificano a
formare un efficace sistema di fasci conduttori: le
nervature fogliari.
L’anidride carbonica invece diffonde grazie alla presenza di
spazi intercellulari presenti nello spessore del lembo, il
mesofillo, in collegamento con gli stomi.
La forma è quindi un adattamento alle varie funzioni svolte
alle quali corrispondono diverse morfologie.
È infatti anche il principale organo traspirante della pianta
che permette la risalita dell’acqua dalle radici fino a tutti
gli organi epigei.
Il lungo e sottile picciolo, le piccole dimensioni e la forma grossolanamente romboidale o triangolare
permettono alle foglie della betulla di vibrare a ogni alito di vento, conferendo a questa pianta latifoglia
una particolare bellezza.
Le foglie sono semplici, a margine dentellato, e disposte in modo alterno sui rami; in alcune specie,
come in Betula pubescens, possono essere ricoperte da una fine peluria
Lo sviluppo delle foglie
I primordi fogliari (abbozzi), si sviluppano ad
intervalli regolari immediatamente al di sotto
dell’apice caulinare in accrescimento.
Prendono origine da divisioni cellulari che
avvengono nella tunica del cono vegetativo
Gli abbozzi si sviluppano dapprima all’apice
(meristema apicale) a somiglianza del cono
vegetativo.
La crescita apicale viene tuttavia inibita presto.
Alla base delle foglie c’è una banda meristematica
(meristema basale)che assicura la formazione di
nuove cellule per l’allungamento di questi
organi.
Lo sviluppo delle foglie
La crescita in larghezza viene affidata a meristemi periferici.
Proprio per l’attività di questi meristemi avremo le diverse
forme fogliari (es. lobata o palmata).
Dietro ai meristemi, si ha l’allungamento delle cellule
neoformate (zona di distensione).
Nelle pteridofite le foglie si sviluppano a partire da una cellula
apicale che dividendosi, forma cellule figlie secondo due
dimensioni (bidirezionale) e rimane attiva fino al completo
sviluppo della foglia.
Nella radice e nel fusto si ha una crescita continua. Nelle foglie,
invece, si ha un rapido raggiungimento della loro taglia
definitiva.
I meristemi fogliari (o la cellula apicale nelle pteridofite) cessano
la loro attività con l’eccezione delle graminacee e di
numerose monocotiledoni, nelle quali i meristemi basali della
foglie restano sempre attivi e producono nuovi tessuti
fogliari.
I tipi di foglie:
generalità
Le piante producono, lungo il fusto, foglie con diversa
morfologia e funzione.
Nelle plantule si riconoscono le foglie
embrionali(cotiledoni).
Sopra queste si possono trovare in alcune piante
piccole foglie ridotte (catafilli).
Al di sopra si formano le foglie normali (nomofilli)che
corrispondono agli organi propri della fotosintesi.
Verso l’apice e nella zona dei fiori si trovano le brattee
(ipsofilli).
I segmenti fiorali, sepali, petali, stami e carpelli
corrispondono a foglie modificate.
Le strutture che per la loro conformazione possono
essere fatte risalire ad un’origine comune sono
dette omologhe.
I differenti tipi di foglie sono organi omologhi benchè la
loro funzione e la loro forma siano talora
profondamente modificate (metamorfosi) tanto da
non essere più riconoscibili come foglie.
Le foglie embrionali e le foglie primarie
Le foglie embrionali (cotiledoni)sono già
presenti nella plantula racchiusa nel
seme.
Nelle monocotiledoni compare un solo
cotiledone, nelle dicotiledoni due mentre
nella maggior parte delle gimnosperme
ne compaiono molti.
In certe piante i cotiledoni rimangono sotto il
terreno, rinchiusi nel seme al momento
della germinazione (germinazione ipogea).
Essi non sono visibili sul fusto aereo; la
loro funzione è quella di organi di
riserva sotterranei, ricchi di lipidi e di
glucidi che saranno utilizzati per
l’alimentazione della plantula sino al
momento in cui questa sarà in grado di
svolgere la fotosintesi, cioè dopo la
formazione delle foglie.
Nel caso della germinazione epigea i cotiledoni vengono sollevati fuori
dal terreno.
In molti casi diventano verdi e possono contribuire per qualche
tempo alla produzione fotosintetica di composti utili alla pianta
I catafilli sono piccoli organi, con
funzione protettiva, a forma di
squama, spesso privi di pigmenti e
perciò di colore brunastro.
Le squame delle gemme(perule), che di
queste rappresentano un organo
protettivo, derivano da foglie
primarie.
In certe piante le foglie primarie si
presentano come appendici
insignificanti del fusto, inserite fra i
cotiledoni e le foglie normali.
Le foglie normali- morfologia esterna
In una giovane foglia, non ancora
differenziata del tutto, si distinguono due
porzioni:
1.
la foglia inferiore, verso la base
2.
la foglia superiore verso l’apice.
La foglia inferiore, dà luogo alla base
allargata della foglia e alle stipole
(espansioni appiattite), caratteristiche
di certe famiglie di dicotiledoni(es.
Rosacee).
Il lembo fogliare (lamina) ed il picciolo si
sviluppano dalla foglia superiore.
Il lembo può essere intero, dentato o
diviso in foglioline(palmato o
pennato).
In certe monocotiledoni, soprattutto nelle
graminacee, le foglie sono prive di
picciolo, ma la lamina si allarga alla
base in una guaina che avvolge il fusto.
Essa prende origine dalla foglia inferiore.
Quando le foglie formate per prime sono
diverse da quelle formate
successivamente si distinguono foglie
primarie e foglie secondarie.
Ad esempio l’Hedera helix produce
inizialmente le caratteristiche foglie
trilobate. Le foglie secondarie sono
invece ovoidali appuntite e compaiono
solamente in condizioni di illuminazione
sufficientemente intensa.
Alcune piante come la Seleginella hanno un
fusto dorso-ventrale su cui sono inserite
sulle due facce foglie di differente taglia.
Questa conformazione viene detta
anisofillia. Questa è indotta anche dal
peso sui rami laterali dell’Acer platanoides e
dell’ Aesculus hippocastanum.
In alcune piante, le foglie secondarie non
sono tutte della medesima forma.
Questo fenomeno viene detto eterofillia.
Il Ranunculus aquatilis produce sotto la
superficie dell’acqua foglie finemente
laciniate. Successivamente, raggiunge
la superficie e produce foglie laminari
galleggianti. In questo caso si tratta di
eterofillia indotta.
La lamina è attraversata da un sistema di
fasci conduttori che corrispondono
alle nervature fogliari.
Le nervature attraversano il lembo in
modo da assicurare il trasporto delle
sostanze mediante i fasci conduttori e
contribuiscono anche in modo
consistente alla funzione di sostegno
meccanico del lembo che spesso è
molto sottile.
La disposizione delle nervature è un carattere
importante per distinguere le monocotiledoni
dalle dicotiledoni.
Nelle monocotiledoni le nervature sono parallele e
collegate fra loro da un reticolo di nervature più
fini.
Nelle dicotiledoni al contrario le nervature formano
un reticolo simile ad una filigrana.
Dalla nervatura mediana si dipartono le nervature
laterali, collegate fra loro da un reticolo di
nervature più fini.
Le superfici comprese fra i fasci conduttori si
chiamano campi intercostali e sono alimentate
da nervature finemente ramificate che
terminano cieche.
Le nervature e perciò anche i vasi conduttori sono
via via più sottili a partire da quella principale
verso le estremità di quelle laterali..
Molte felci e, fra le gimnosperme Ginko biloba,
presentano un tipo di innervazione molto
primitivo.
Le nervature si ramificano biforcandosi (innervazione
dicotomica) e non posseggono collegamenti
trasversali.
Palminervia: Foglia con diverse nervature ugualmente robuste che si irradiano come un
ventaglio dall'estremità del picciolo
angolinervie- le nervature si intersecano a reticolo(molte dicotiledoni)
parallelinervia - Foglia con nervature uguali e parallele
plurinervie - Foglia con nervatura ramificata
uninervia - Foglia con una sola nervatura. Rara, nel pino.
penninervia - Foglia con una nervatura principale mediana da cui partono nervature
minori laterali.
peltinervia - Foglia con nervature ugualmente robuste che si irradiano come i raggi di una
ruota, con picciolo inserito nel centro del lembo.
Struttura interna della lamina
Struttura interna della lamina
La struttura interna della lamina fogliare è
essenzialmente adattata a due
funzioni:
1.
Fotosintesi
2.
limitazione e controllo della perdita di
acqua legata alla traspirazione.
i problemi di adattamento delle piante
terrestri si riferiscono a questi due
punti.
Alla costituzione della lamina prendono
parte soprattutto tre tipi di tessuti:
•
Tessuto di rivestimento (epidermide)
•
Tessuto di assimilazione (mesofillo)
•
Fasci conduttori.
Con riferimento alla disposizione di questi
tessuti nella lamina si distinguono
foglie bifacciali, equifacciali e unifacciali.
L’epidermide
L’epidermide fogliare deve svolgere due
funzioni a prima vista incompatibili:
Proteggere dal disseccamento limitando la
perdita di vapor d’acqua(traspirazione);
Deve permettere gli scambi gassosi fra la pianta e
l’ambiente.
La soluzione del problema risiede nel fatto che
l’epidermide riveste la foglia come una
pellicola impermeabile ai gas, ma è anche
interrotta da pori che permettono gli
scambi gassosi fra la pianta e l’ambiente: gli
STOMI.
Questi possono passare, attraverso vari stadi
dall’apertura massima alla chiusura
completa, in modo da regolare gli scambi
gassosi.
Assolve anche ai compiti di protezione dai
danni fotossidativi, dai danni meccanici, da
infezioni dovute a microrganismi.
Basilico con stomi e goccia di olio
Di solito è monostratificata,
composta da cellule vive che
sono conformate come un
puzzle con tessere
concatenate e prive di spazi
intercellulari.
Vanno a costituire una struttura
resistente alle deformazioni
laterali.
Di norma le cellule epidermiche
non posseggono cloroplasti,
ma possono essere colorate
da sostanze disciolte nel
succo vacuolare come i
derivati flavanici, cui si
devono certi disegni colorati
sulla superficie delle foglie.
Le pareti cellulari esterne sono
frequentemente ispessite ed impregnate
di sostanze lipofile( cutine, cere).
La faccia esterna delle cellule epidermiche è
ricoperta da una pellicola di cutina, la
cuticola a cui si deve la debole permeabilità
al vapore acqueo e ad altri gas.
Questo effetto di barriera è rafforzato in
alcune piante, da depositi di cere, posti
sopra la cuticola.
Le piante a foglie scure(cavolo rosso),
presentano spesso sulla cuticola depositi
biancastri di cera che possono essere
asportati, ed impediscono alle foglie di
bagnarsi.
Peli
PELI VIVI In qualche caso possono secernere dei composti (oli essenziali) e vengono detti peli
secretori.
L’epidermide può formare anche dei peli che
possonosono
essere
uni o pluricellulari,
che famiglia Urticaceae.
I peli urticanti
presenti
soprattuttoma
nella
tuttavia
prendono
origine
da
una
cellula
A causa
della mineralizzazione con silice, la loro parete diventa particolarmente fragile. È per questo
epidermica.
cheSil’apice
del pelo
facilmente per contatto in un punto predeterminato, liberando il
distinguono
peli si
vivispezza
e peli morti.
contenuto di acetilcolina.
I peli morti sono pieni d’aria e a causa della riflessione totale della luce appaiono
bianchi. Sulle foglie formano spesso un fitto feltro.
Si suppone che questo feltro protegga le foglie dalle forti radiazioni ed in particolare da
quelle a lunghezza d’onda corta.
Un tipo particolare sono i peli squamosi o squame assorbenti che compaiono sulle foglie
della famiglia Bromeliaceae (Ananasso). Fra la superficie dell’epidermide e i peli
squamosi si trova uno spazio pieno d’acqua (e di sali nutritivi) che viene attirata per
capillarità verso la base del pelo.
In caso di deficit idrico i peli squamosi si appiattiscono contro l’epidermide, chiudendo
il poro. Le squame assorbenti rappresentano un adattamento di queste piante alla
vita epifitica.
Tricomi al microscopio
Dando una descrizione semplificata del processo di assorbimento attraverso i tricomi,
possiamo dire che fino a che la pianta è in attesa di acqua, l'ala rimane sollevata per l’ingresso di
acqua. Altrimenti, si abbassa.
Tricoma in sezione con ala abbassata. É evidenziato
in blu il percorso dell'acqua che cade sulla foglia.
In questa sezione sono ben visibili anche i
fasci conduttori
tricomi al microscopio ottico
Le emergenze
Sono strutture pluricellulari costituite dall’epidermide e da un
tessuto subepidermico. I “tentacoli” delle piante carnivore come
la Drosera, così come la base del pelo urticante dell’ortica, sono
esempi di emergenze.
stomi
Sono costituiti da un complesso di
cellule epidermiche specializzate.
L’apparato stomatico consta di un paio di
cellule stomatiche affiancato da due o
più cellule annesse. Le cellule
stomatiche assicurano la chiusura e
l’apertura degli stomi. Sono in
genere reniformi e contengono
cloroplasti.
Sono raggruppate a due a due. Le
pareti cellulari che si affrontano
non sono associate dalla lamella
mediana, ma lasciano una fessura al
centro la rima stomatica. Il grado di
apertura aumenta con l’aumento
del grado di turgore delle cellule
stomatiche. Inversamente la rima si
chiude con il diminuire del turgore
e quindi l’afflosciamento delle
cellule.
Gli stomi effettuano i cosiddetti effetti
nastici di turgore e nel movomento
distinguiamo una componente
meccanica ed una fisiologica.
Fisiologica= riguarda le cause
delle variazioni di turgore.
Meccanica= è legata al modo in
cui le variazioni di volume
delle cellule, dovute a
variazioni di turgore, si
traducono in un movimento
orientato. Le pareti delle
cellule sono ispessite solo da
una parte e perciò la
deformazione avviene solo in
un determinato senso.
Struttura di diversi tipi di stomi
In rapporto alla distribuzione degli stomi sulla superficie delle foglie, si possono
distinguere :
Foglie ipostomatiche: gli stomi sono sulla superficie inferiore delle foglie;
Foglie epistomatiche: gli stomi sono distribuiti solo sulla superficie superiore delle
foglie (es. nelle foglie galleggianti di piante acquatiche-ninfea)
Foglie anfistomatiche: gli stomi sono distribuiti su tutta la superficie della foglia, dove
tuttavia il numero può essere diverso nella pagina superiore rispetto a quella
inferiore.
Lo stesso numero degli stomi può variare e va da 15 sino a + di 800 per mm2.
Distribuzione e numero riflettono l’adattamento ecologico in relazione
all’economia d’acqua in determinati habitat.
Le distanze reciproche fra gli stomi sono determinate secondo
modelli specifici detti: ENDONOMA e EZIONOMA
Il mesofillo
Il tessuto fogliare compreso fra l’epidermide superiore e quella inferiore viene
denominato MESOFILLO. Nelle sue cellule si svolge la fotosintesi.
Nelle foglie bifacciali come accade nella maggior parte delle dicotiledoni il mesofillo è
asimmetrico.
Verso la pagina superiore si trova il parenchima a palizzata, formato da cellule cilindriche,
ricche di cloroplasti; le cellule sono orientate perpendicolarmente alla pagina
superiore della foglia e lasciano numerosi spazi intercellulari in modo da assicurare
l’aerazione del tessuto.
Il parenchima a palizzata è di solito monostratificato,
ma può essere pluristratificato in certe piante con
foglie sottoposte ad un forte irraggiamento
solare.
Al di sotto del parenchima a palizzata si trova il
parenchima lacunoso, costituito da cellule irregolari
che lasciano grandi spazi intercellulari per cui il
tessuto assume un aspetto spugnoso.
Nella zona degli stomi gli spazi intercellulari sono
particolarmente ampi e danno luogo a cavità
sottostomatiche. Le cellule del parenchima
lacunoso contengono solamente il 20% dei
cloroplasti di una foglia, mentre il rimanente 80%
è situato nel parenchima a palizzata.
I fasci conduttori della lamina sono disposti nella zona
di confine tra i parenchimi a palizzata e lacunoso.
Sono di tipo collaterale chiuso. Lo xilema si trova
verso la pagina superiore mentre il floema verso
quella inferiore.
Nelle nervature principali i fasci sono accompagnati da
tessuti di sostegno (sclerenchima). Nelle nervature
più sottili i fasci sono costituiti solo da tracheidi
xilematiche che terminano cieche nel tessuto.
I fasci conduttori della foglia sono tuttavia interamente
circondati da una guaina perivascolare che è composta
da cellule parenchimatiche prive di spazi
intercellulari.
La guaina, serve da intermediaria per il trasporto nei
vasi conduttori di sostanze provenienti dal
mesofillo. Nelle foglie C4 le cellule di questa guaina
sono provviste di cloroplasti e svolgono particolari
compiti nell’organicazione della CO2
Disposizione ed orientazione delle foglie
OPPOSTE o DISTICHE:
Foglie inserite a coppie sul ramo, alla stessa altezza ,
l'una di fronte all'altra.
TERNATE:
Come le opposte ma inserite a tre a tre, alla stessa altezza.
VERTICILLATE:
Foglie inserite alla stessa altezza in numero superiore a tre.
ALTERNE O SPARSE:
Foglie inserite in punti diversi alternativamente secondo una spirale
Le foglie sono inserite sul fusto
secondo un modello
geometrico che viene detto
FILLOTASSI.
È un carattere specifico molto
utile nella classificazione delle
piante. La disposizione delle
foglie ha il compito di evitare
che le foglie nuove
ombreggino quelle formate in
precedenza in modo che la
fotosintesi si svolga nel
migliore dei modi.
La diversità dei modelli fillotassici
può essere ricondotta ad alcuni
tipi fondamentali
La posizione delle foglie è già determinata dalla posizione degli abbozzi fogliari
nell’apice del germoglio.
Oltre ad evitare l’ombreggiamento reciproco, le foglie devono essere disposte in
posizione ottimale in relazione all’incidenza della luce.
In effetti le superfici fogliari sono orientate in modo che la luce incidente risulti
perpendicolare quando il sole si trova allo zenit. Nel caso di radiazione molto
forte, come nei luoghi molto assolati, può accadere che le foglie siano sottoposte
ad eccessivo riscaldamento.
Le cosiddette “piante bussola” ad esempio (Lactuca seriola) aggirano il problema
orientando il lembo verticalmente e secondo la direzione nord-sud come l’ago di
una bussola; in tal modo la luce debole del mattino e quella della sera illuminano
le lamine fogliari, che sono per lo più equifacciali, mentre quella forte del
mezzogiorno illumina i margini.
La metamorfosi delle foglie
ciliegio
Come avviene in altri organi della pianta, anche nelle foglie si
osservano metamorfosi.
Vediamo delle metamorfosi in relazione delle funzioni specifiche.
Brattee (bractea =lamina di metallo!)
Le brattee proteggono i fiori e le infiorescenze come un involucro,
oppure presentano colori vivaci in modo da attirare insetti
pronubi (es. le brattee fiorali violacee del rampicante tropicale
Bouganville;
oppure l’Euphorbia pulcherrima
spine
Sono strutture appuntite, formate da
tessuti sclerenchimatici offrono
vantaggi selettivi alle piante
proteggendole per esempio da
morsi di animali.
Non esistono solo spine di origine
caulinare, ma anche di origine
fogliare e cioè foglie o stipole
modificate.
Spine di origine fogliare si osservano nel
crespino, che presenta numerosi
stadi intermedi fra foglie normali e
spine raggruppate per tre, e nelle
cactacee.
Spine di origine stipolare sono presenti
sempre a paia, si osservano nella
robinia e nelle euforbie succulente.
viticci
Le piante possono fissarsi, ed
arrampicare sui sostegni,
mediante strutture filiformi.
Tutte le parti della foglia sono in
grado di originare viticci.
Quelli di Pisum sativum derivano
dalle ultime foglioline della
foglia pennato-composta.
Alcune piante si arrampicano
mediante i piccioli che si
avvolgono attorno a sostegni
(Nepethes-pianta insettivora).
Organi di riserva
In certe piante di luoghi aridi, le foglie sono
modificate in organi di riserva d’acqua.
Sono spesso equifacciali. Al fine di svolgere
questa funzione, possono essere
coinvolti epidermide e tessuti
subepidermici o anche il mesofillo. Le
cellule di riserva posseggono sempre
vacuoli di grandi dimensioni e per
questo le foglie mostrano un aspetto
carnoso e “succulento”.
Le piante provviste di queste foglie, sono
denominate come piante a succulenza
fogliare.
La succulenza fogliare( e quella caulinare)
sono collegate alla comparsa del
metabolismo CAM.
Tipiche piante a succulenza fogliare sono le
agavi, i semprevivi, i generi Sedum…
I bulbi sono strutture sotterranee che consistono
di un fusto estremamente raccorciato, su cui
sono inseriti i catafilli carnosi, squamiformi
ed appressati gli uni sopra gli altri. I catafilli
sorgono dalla base di foglie normali e
fungono da organi di riserva.
I bulbi sono frequenti nelle monocotiledoni (es.
cipolla). Rappresentano le forme biologiche
che permettono a determinate piante di
superare periodi stagionali sfavorevoli.
Alla ripresa di un nuovo periodo vegetativo
gemme ascellari, situate fra le foglie di riserva,
danno luogo a nuove strutture vegetative
consumando le sostanze di riserva,
immagazinate.
Le piante che svernano mediante organi
sotterranei durevoli, siano essi bulbi, tuberi o
rizomi si dicono geofite.
Fillodi
Sono piccioli modificati in
organi adatti alla fotosintesi.
In alcuni casi la lamina fogliare
è fortemente ridotta. Molto
caratteristica è Acacia
heterophylla nella quale
compaiono sulla stessa
pianta forme di passaggio
dalle tipiche foglie pennate a
fillodi del tutto privi di
lamina. Anche gli organi
assimilatori succulenti e
cilindrici di Kalanchoë tubiflora
(CAM)sono fillodi.
Organi di cattura per animali
cephalotus
Nelle piante carnivore le foglie sono trasformate in organi di cattura
delle prede.
Per la verità, le piante carnivore si nutrono di insetti per necessità.
Sono piante, infatti, che in natura vivono in ambienti
particolarmente poveri dei nutrienti necessari alla crescita di tutte
le piante.
Le ritroviamo in paludi, in torbiere acide, su rocce spoglie,
abbarbicate su tronchi di altre piante, sott'acqua... Le piante
carnivore riescono a sopravvivere in questi ambienti sterili e
inospitali ricavando il loro nutrimento dagli insetti. In questo
modo, anche se il terreno e' sterile, queste piante riescono
comunque a crescere e moltiplicarsi.
dionea
Le modalità di
funzionamento delle foglie
La legge del “fattore limitante” di Liebig
I processi vitali svolti dalle foglie sono influenzati dai fattori ambientali e si
possono avere diversi fenomeni adattativi, tra i quali i più importanti sono la
fotosintesi e lo scambio gassoso.
In condizioni naturali, ogni processo fisiologico che avviene nelle piante, è
influenzato da numerosi fattori ambientali. Vige qui la legge di J.Von Liebig
(1803-1873) che dice:
Se un processo dipende da molti fattori esso può incrementare la sua intensità
solo in relazione a quel fattore che di volta in volta è presente al minimo e che
perciò agisce come fattore limitante.
È una legge valida anche per la fotosintesi sulla quale influiscono i fattori luce,
temperatura e concentrazione di CO2.
Fattore luce.
Gli ambienti dove vivono le piante, possono essere diversi fra loro rispetto alle
condizioni di luce, e la vita si ha solo negli organismi che si adattano a quelle
condizioni di luce dominanti in quell’ambiente.
Si distinguono 2 tipi di adattamento e cioè piante
eliofile adattate a forti luminosità
sciafile adattate a deboli luminosità.
In uno stesso albero possono sussistere entrambe le condizioni a seconda
dell’esposizione!
•
•
•
•
affinché si abbia la formazione di tessuti o di organi, la produzione primaria
lorda (PPL), che comprende anche la quota di produzione che verrà spesa per
la respirazione della pianta, deve superare la respirazione stessa (R)
il punto in cui si ha equilibrio fra fotosintesi e respirazione è detto “punto di
compensazione”
al crescere della quantità di luce cresce anche la fotosintesi fino al punto, detto
“punto di saturazione”, in cui rimane praticamente costante
i punti di compensazione e di saturazione possono notevolmente variare da
specie a specie.
L’adattamento alla fotosintesi alle condizioni di luce è evidenziato dalle curve di saturazione
Specie sciafile: hanno un punto di
compensazione basso ed una
respirazione meno intensa (es.:
faggio e leccio giovani, abete rosso,
bosso, carpino, ecc.); non sono in
grado di vegetare in piena luce
Specie eliofile: hanno punti di
compensazione e di saturazione
elevati ed una respirazione più
intensa (es.: roverella, pini, frassino,
ecc.); non possono vegetare in
condizioni di bassa intensità
luminosa
Non è solo una diversità di
tipo fisiologico, ma
anche di tipo
morfologico.
I tipi eliofili sono
caratterizzati da due o più
strati di parenchima a
palizzata, mentre si
osserva un solo strato nei
tipi sciafili( è sufficiente
a catturare i pochi quanti
di luce disponibili).
Anche a livello cloroplastico
ci sono differenze:
Sciafili: per ogni granum si
osserva un numero
maggiore di tilacoidi
appressati ed i complessi
antenna sono più ricchi
di pigmenti antenna.
Eliofili: numero minore di
tilacoidi.
A- cloroplasto di sole; B- cloroplasto d’ombra
In condizione di luce debole i tipi sciafili sono superiori, perché già
a bassa luminosità assicurano un guadagno fotosintetico netto di
carbonio.
La loro fotosintesi massimale, la produttività fotosintetica, resta bassa
rispetto alle eliofile che, per fissare la CO2, sono più valide in
condizioni di luce forte, poiché possono utilizzare una quantità
più elevata di NADPH + H+e di ATP prodotti mediante le
reazioni fotochimiche grazie alla loro capacità elevata di
assimilazione di CO2.
I tipi adattati alla luce forte presentano perciò una produttività
fotosintetica significativamente più elevata.
(le piante coltivate + importanti presentano questo tipo di
adattamento!)