caricato da
giu_acsf
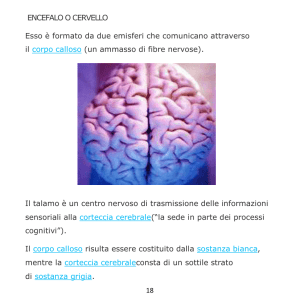
CAP 5

CAP 5: sensazione e percezione Percezione: inizia con uno stimolo nell’ambiente, come un suono o una luce che stimola uno degli organi di senso come l’orecchio o l’occhio. Sensazione: consiste nella prima elaborazione che si verifica. È la rappresentazione mentale dello stimolo originale che deriva dalle varie fasi di elaborazione dunque la percezione può essere considerata anche come il processo di costruzione del percetto. Nella percezione tutti i sensi sono fondamentali. Durante un pasto il tatto renderà più interessante la nostra esperienza così come i segnali visivi renderanno più allettanti i cibi presenti sul piatto. Ogni sistema ha una propria struttura anatomica per recepire, filtrare e amplificare le informazioni provenienti dall’ambiente. ogni sistema possiede delle cellule recettive specializzate che traducono lo stimolo ambientale in segnali neurali. questi segnali vengono poi fatti passare attraverso specifiche vie sensoriali nervose così da proiettarli attraverso il midollo allungato dove ritroviamo il nervo glossofaringeo e il nervo vestibolo cocleare. L’unico nervo a andare subito nella corteccia e il nervo olfattivo. Le cellule ricettive rispondono a una gamma limitata di stimoli. questo tipo di cellule si attiva solo quando viene superato il livello minimo di intensità dello stimolo. le soglie non sono fisse ma si adattano ai cambiamenti ambientali. Ad esempio le cellule ricettive della vista degli esseri umani rispondono solo a una piccola regione dello spettro elettromagnetico. le lunghezze d’onda della luce comprese tra i 400 700 nanometri. Quelle uditive a una frequenza fino a 20 Herz. Lo stimolo minimo in grado di attivare un percetto viene detto valore di soglia dello stimolo. Per adattamento si intende invece il modo in cui sistemi sensoriali rimangono finemente sintonizzati. Questo permette di regolare la sensibilità del sistema sensoriale all’ambiente che lo circonda e ai suoi cambiamenti. l’adattamento nel sistema olfattivo avviene rapidamente così come in quello uditivo. Alcuni neuroni continuano a scaricare durante l’intera durata dello stimolo ma il loro tasso di scarica diminuisce dunque più dura lo stimolo meno frequenti sono i potenziali d’azione. Anche il sistema visivo si adatta ai cambiamenti dell’intensità luminosa dell’ambiente, quando ci spostiamo da una zona luminosa a una più buia ci vorrebbero dei 20/30 minuti affinché le persone riescano a recuperare una buona visione notturna. Acuità: è la capacità di distinguere tra gli stimoli entro una stessa modalità sensoriali. Dipende da due fattori. Il primo il modo in cui è progettato il sistema di raccolta dello stimolo, il secondo riguarda il numero e la distribuzione dei recettori.ad esempio noi abbiamo più recettori sulle nostre dita che sulla schiena quindi possiamo distinguere meglio gli stimoli utilizzando le dita. Quando un sistema sensoriale dedica più recettori a un certo tipo di informazione si riscontrerà un incremento nella zona corticale di quell’informazione. Lo stimolo sensoriale viene tradotto in attività neurale dei recettori e inviata alle regioni Sub corticali e corticali del cervello così da poter essere elaborato. lo stimolo però non è l’unico fattore che contribuisce alla consapevolezza sensoriale entrano in gioco anche i sistemi attentivi, mnemonici ed emozionali. Nonostante questo la maggior parte della stimolazione sensoriale non raggiunge il livello di coscienza. La strada però non è solo dagli organi di senso al cervello. le connessioni neurali si muovono in entrambe le direzioni. Questo si nota soprattutto tra la corteccia e la sotto corteccia dove i segnali provenienti dal sistema visivo, uditivo, somatosensoriale e gustativo effettuano sinapsi entro il talamo prima di proiettare verso le specifiche regioni della corteccia. la via visiva passa attraverso il nucleo genicolato laterale del talamo ( LGN) il sistema uditivo attraverso il nucleo genicolato mediale( MGN), la via somatica attraverso il complesso nucleare ventrale posteriore e la via gustativa attraverso il nucleo ventrale postero mediale.sembrerebbe che i nuclei talamici siano 1 interconnessi fornendo un’integrazione multisensoriale. il talamo riceve anche connessioni discendenti dalle regioni sensoriali primarie della corteccia ed altre aree come i lobi frontali. queste connessioni sembrano fornire una via attraverso cui la corteccia può controllare il flusso di informazioni dei sistemi sensoriali. 2 Udito: percorsi neurali dell’udito Le onde sonore giungono all’orecchio ed entrano nel canale uditivo. all’interno di questo canale le onde vengono amplificate. le onde così viaggiano fino all’estremità più remota del canale dove colpiscono la membrana timpanica o timpano facendola vibrare. Superata la membrana timpanica le vibrazioni a bassa pressione viaggiano fino all’orecchio medio pieno d’aria e mettono in moto tre ossicini (martello, incudine e staffa) che a loro volta provocano la vibrazione di una seconda membrana chiamata finestra ovale. La finestra ovale è la via di accesso alla coclea piena di fluido. che costituisce la struttura principale dell’orecchio interno. All’interno della coclea vi sono delle piccole cellule ciliate posizionate lungo la superficie interna della membrana basilare. queste cellule costituiscono i recettori sensoriali del sistema uditivo. Sono composte da 200 sottili filamenti detti stereociglia che fluttuano nel fluido. Le vibrazioni sulla finestra ovale producono delle piccole onde nel fluido che a loro volta muovono la membrana basilare piegando le stereociglia. La posizione di questi neurorecettori sulla membrana basilare determina la selettività alla frequenza. questo è dato dallo spessore e dunque rigidità della membrana che varia a seconda della lunghezza dalla finestra ovale all’apice della coclea. Vicino alla finestra ovale la membrana è spessa e rigida e le cellule ciliate in questo punto rispondono a vibrazioni ad alta frequenza. all’estremità della coclea la membrana è sottile e meno rigida; le cellule ciliate in questo punto rispondono solo a basse frequenze. la disposizione spaziale dei recettori sonori è detta tonotopica. Essendo presente una mappa tono topica già a questo stadio vi è una distinzione riguardo all’informazione sulla sorgente sonora. le cellule ciliate agiscono ancora da meccanocettori poiché i canali ionici sono aperti in maniera meccanica e permettono l’ingresso degli ioni caricati positivamente di potassio e calcio. Quando la cellula è sufficientemente de polarizzata rilascerà trasmettitori nelle sinapsi tra la base della cellula ciliata è una fibra nervosa afferente. In questo passaggio avviene la conversione da evento meccanico a segnare neurale. Il nostro sistema uditivo risulta più sensibile a suoni che vanno dai 1000 ai 4000 Herz range in cui ritroviamo anche buona parte della comunicazione umana come linguaggio o il pianto dei neonati affamati. Il viaggio delle informazioni uditive contiene numerosi sinapsi tra le cellule ciliate e la corteccia. le informazioni provenienti dalle cellule ciliate vengono incanalate nel nervo cocleare detto anche il nervo acustico, che proietta l’informazione al nucleo cocleare nel bulbo; gli ioni emessi dal nucleo proiettano l’informazione fino al ponte e si dividono innervando i nucleo Olivari di destra e quello di sinistra. In questo punto avviene per la prima volta la condivisione degli impulsi provenienti da entrambe le orecchie. gli assioni derivanti dei nuclei cocleare e Olivari proiettano al collicolo inferiore che si trova più in alto del mesencefalo. In questo punto i segnali uditivi possono accedere alle strutture motorie permettendo di orientare la testa in direzione del suono. A questo punto alcuni segnali uditivi vengono proiettati al nucleo del lemnisco laterale nel mesencefalo dove avviene l’elaborazione del tempo. Dal mesencefalo l’informazione arriva al nucleo genicolato mediale del talamo e che a sua volta proietta alla corteccia uditiva primaria che troviamo nella parte superiore del lobo temporale. I neuroni mantengono per tutta la via una selettività alla frequenza è una disposizione tonotopica che si riscontra anche nella corteccia uditiva primaria. Nella parte rostrale tende a rispondere ai suoni a bassa frequenza mentre nella parte caudale a suoni ad alta frequenza. Le Mappe tonotopiche sono state trovate anche nelle aree uditive secondarie della corteccia. Studi hanno dimostrato che la selettività uditiva umana sia più acuta di molte altre specie ad eccezione del pipistrello. 3 La variazione delle frequenze è fondamentale affinché si possono identificare parole o musica. La percezione uditiva oltre ad individuare il contenuto di uno stimolo ostico riesce a localizzare i suoni nello spazio. Per le ricerche sul “dove” sono stati usati molti animali tra cui il barbagianni.questi animali si affidano due segnali per localizzare i suoni il primo è la differenza di tempo impiegato dal suono per raggiungere le tue orecchie detto anche tempo interaurale. Il secondo segnale è la differenza di intensità del suono alle due orecchie. Infatti a meno che il suono non sia perfettamente parallelo all’orientamento della testa esso raggiungerà prima un orecchio rispetto all’altro. Queste differenze sono aumentate nel barbagianni data la sua anatomia in quanto un orecchio risulta più in alto di un altro. Esperimenti scientifici hanno portato ad insegnare al barbagianni a muovere la testa verso il suono. Il suono veniva proiettato attraverso delle cuffie e si è riscontrato che nel momento in cui una delle orecchie veniva tappata il barbagianni rilevava il suono ma non gli era possibile localizzare la fonte.altri studi hanno ipotizzato un modello neurale specifico che afferma che i neuroni del tronco-encefalico riescono a codificare la differenza di tempo operando come rilevatori di coincidenze ma che per essere attivati devono ricevere in maniera simultanea input da entrambe le orecchie. Un altro modello prende in esame l’intensità del segnale e la differenza interaurale. 4 OLFATTO: indispensabile per i mammiferi per comprendere se la commestibilità del cibo. Le molecole odorose entrano all’interno della cavità nasale sia durante l’atto di respirazione sia quando annusiamo.io doranti possono entrare nel sistema attraverso la bocca viaggiando a ritroso fino alla cavità nasale.i l modo in cui i recettori olfattivi leggono le molecole odorose non è a oggi del tutto conosciuto.una delle ipotesi più accreditate sostiene che i recettori olfattivi traducono le molecole odorose legandole ed incorporandole all’interno della membrana della mucosa nel tetto della cavità nasale chiamato epitelio olfattivo. Nell’epitelio si trovano più di 1000 tipi di recettori che rispondono a un numero limitato di odoranti sebbene un odorante si possa legare a più di un tipo di recettore. un’altra ipotesi mette in evidenza l’importanza della vibrazione molecolare che contribuisce al riconoscimento dell’autore. questa ipotesi spiega come molecole olfattive con vibrazioni simile abbiano risposto olfattive simili e come molecole con una struttura molecolare simile abbiano risposte effettive diverse come sono diverse le loro vibrazioni. I recettori olfattivi sono detti neuroni bipolari poiché le loro estremità si estendono ai lati opposti del corpo cellulare. quando una molecola odorosa innesca il neurone il segnale viene inviato ai neuroni del bulbo olfattivo detti anche glomeruli. Un neurone bipolare può attivare più di 8000 glomeruli e a sua volta un glomerulo può ricevere input da più di 750 recettori. gli assoni continuano la loro corsa formando il nervo olfattivo. la loro destinazione la corteccia olfattiva primaria detta anche corteccia piriforme situata nella giunzione ventrale della corteccia frontale e temporale. Il nervo olfattivo raggiunge la corteccia olfattiva primaria senza passare dal talamo. la corteccia olfattiva primaria proietta un’area olfattiva secondaria entro la corteccia orbitofrontale ampliando la connessione con altre regioni del cervello inclusi il talamo, ipotalamo, ippocampo e amigdala. Si spiega il motivo di come l’olfatto influisca il comportamento, l’attenzione, la memoria e le emozioni. Sebbene gli studi sull’olfatto siano pochi data la complicanza nella valutazione uno studio ha individuato la diversa risposta neurologica tra l’olfatto e l’annusare. La sensazione di un odore produceva l’attivazione nelle aree laterali della corteccia orbitofrontale, regione considerata parte della corteccia olfattiva secondaria e non nella corteccia olfattiva primaria come ci si aspettava. la corteccia olfattiva primaria era legata all’annusare infatti ogni volta che la persona non usava la risposta aumentava indipendentemente dalla presenza o meno dell’odore. Una risposta a tale fenomeno si è data con studi sui ratti affermando che i neuroni della corteccia olfattiva primaria tendono ad adattarsi in maniera molto veloce. La corteccia odorosa primaria può essere considerata fondamentale per l’identificazione di variazione degli odori mentre la corteccia secondaria sia indispensabile per identificare l’odore. Le narici per aumentare la percezione degli odori tendono ad alternarsi. questo cambiamento porta un effetto sul modo in cui l’odore viene elaborato. La presenza di due narici con dimensioni leggermente diverse e fornisce al cervello un’immagine olfattiva leggermente differente. La ricerca ha dimostrato che la molecola odorosa somministrata nella narice più ampia risultava più intensa rispetto a quella presentata nella narice più stretta. 5 IL gusto: come per l’olfatto viene considerato un senso chimico poiché interpreta l’ambiente differenziando diverse sostanze chimiche. Il gusto inizia con la lingua sulla quale si possono trovare le pupille (piccole protuberanze) le papille hanno diverse funzioni legati al gusto, alla sensazione. E alla lipasi linguale (enzima che aiuta a demolire i grassi) le papille presenti nelle regioni laterali della lingua e nella regione anteriore contengono molti bottoni gustative mentre quelle centrali non li contengono. i pori gustative rappresentano i condotti che portano l’informazione dalla superficie della lingua fino ai bottoni gustative. ciascun bottone gustativo contiene molte cellule gustative. questo tipo di cellule sono state trovate anche sulle guance e sul palato. I gusti di base sono cinque: salato, aspro, amaro, dolce e umami ( saporito). Nelle papille gustative le cellule gustative danno luogo a sinapsi con i neuroni bipolari i cui assoni formano il nervo della corda del timpano. il nervo si unisce ad altre fibre formando il nervo facciale che proietta al nucleo gustativo posizionato nella regione rostrale del nucleo del tratto solitario nel tronco-encefalico. in contemporanea la regione caudale dello stesso nucleo riceve informazioni sensoriali dal tratto gastrointestinale. Questa interazione di informazioni può dare luogo a reazioni rapide come ad esempio può essere il conato di vomito si si avverte un sapore non gradito. La sinapsi successiva avviene nel nucleo ventrale posteriore mediale del talamo. gli assoni in uscita fanno poi sinapsi nella corteccia gustativa primaria che si trova a livello dell’insula e dell’opercolo. Struttura tra i lobi temporali e frontali. La corteccia gustativa primaria si connette con le aree elaborative secondarie della corteccia orbitofrontale fornendo un substrato anatomico per l’integrazione tra gusto ed olfatto. La lingua oltre ad assaggiare il cibo permette grazie alle papille gustative che contengono recettori nocicettivi di attivarsi con sostanze irritanti come quelle contenute nel peperoncino, nelle bibite gassate o nell’aceto. I segnali di questi recettori seguono un percorso diverso formando il nervo trigemino che porta informazioni dolorifica e segnali riguardo la posizione la temperatura. La percezione del gusto varia a seconda di quante e di quale tipo di papille e di bottoni gustative hanno gli individui. negli esseri umani i bottoni gustative si aggirano tra i 120 e 668 per centimetro quadrato di norma le donne ne presentano di più rispetto agli uomini. La differenziazione dei gusti di base fornisce al corpo informazioni circa il cibo che è stato consumato ad esempio la sensazione di umami informa il corpo che è stato ingerito un alimento ricco di proteine, il dolce di carboidrati, il salato informazioni importanti sull’equilibrio tra minerali o elettroliti ed acqua, l’amaro e l’acido sono segnali di allarme. La corteccia orbitofrontale gioca un ruolo importante nell’elaborazione della gradevolezza del valore di ricompensa legato all’alimentazione. Degli studi sono stati fatti con l’assunzione del cioccolato e la gratificazione derivante da questo. è stato visto che la porzione posteromediale della corteccia orbitofrontale veniva attivata quando il cioccolato era indicato come molto gratificante e vi era un forte desiderio di mangiarne ancora. 6 Somatosensazione: È la percezione somatosensoriale di tutti gli stimoli meccanici che riguardano il corpo. è probabilmente il sistema sensoriale che include maggiori altri sistemi sensoriali. in quanto è formato da un’intricata serie di recettori specializzati in vaste proiezioni e in diverse regioni del sistema nervoso centrale. I recettori si trovano sotto la pelle ed includono diversi tipi di recettori che si differiscono per velocità con cui si adattano per la propria sensibilità a vari tipi di tatto. sono: - Corpuscoli di Meissner; - Cellule di Merkel; - Corpuscoli di pacini; - Corpuscoli di Ruffini. La sensazione di dolore viene segnalata dai nocicettori che sono di tre tipi: - Recettori termici: rispondo al caldo o al freddo; - Recettori meccanici: rispondono a intense stimolazioni meccaniche; - Recettori polimodali: rispondono ad una vasta gamma di stimoli nocivi tra cui traumi meccanici e sostanze chimiche. I nocicettori sono localizzati nella pelle, sotto la pelle nei muscoli e nelle giunture. i neuroni dolorifica afferenti possono essere sia mielinizzati sia deminilizzati. Le prime conducono velocemente le informazioni di dolore producendo un’azione immediata. le seconde sono responsabili del dolore sordo e prolungato che segue di solito il dolore acuto delle prime. Alcune cellule nervose specializzate permettono di ricavare informazioni riguardo alla posizione del corpo (propriocezione) queste cellule permettono al sistema motorio e a quello sensoriale di rappresentare l’informazione sullo stato dei muscoli e degli arti ad esempio se sono allungati o usati per qualche movimento. I recettori somatosensoriali hanno i corpi cellulari nelle radici dorsali dei gangli ed entrano nel midollo spinale tramite le radici dorsali. Altri entrano in contatto sinaptico con i motoneuroni del midollo spinale per formare archi riflessi. Altri ancora fanno sinapsi con neuroni che inviano gli assoni super le colonne dorsali del midollo spinale fino a raggiungere il bulbo (midollo allungato) punto in cui l’informazione si incrocia verso il nucleo ventrale posteriore del talamo E poi verso la corteccia cerebrale. oltre alle proiezioni corticali le informazioni propriocettive e somatosensoriali vengono proiettate anche in altre strutture sottocorticali come il cervelletto. 7 L’aria in cui le informazioni arrivano inizialmente e detta corteccia somatosensoriale primaria (S1) E contiene le aree di Brodmann 1,2,3. S1 contiene una rappresentazione somatotopica del corpo detta anche homunculus sensoriale. Questa rappresentazione è sensibile all’informazioni che arrivano dalle aree del corpo. Maggiori saranno i recettori in una zona del corpo maggiore sarà l’aria in S1.per esempio le mani coprono una grande parte della corteccia rispetto al tronco. le mappe somatotopiche variano tra le specie in quanto le diverse parti del corpo rivestono un ruolo maggiore in alcune specie piuttosto che in altre. Subito dopo S1 troviamo la corteccia somatosensoriale secondaria (S2).in tale corteccia si trovano rappresentazioni più complesse ad esempio per il tatto le informazioni sono codificate e possono dare informazioni riguardanti la trama o la dimensione dell’oggetto. Ciascun emisfero in S2 riceve informazioni da entrambi i lati del corpo grazie alle proiezioni che attraversano il corpo calloso. Nel 1949 Donald Hebb propose una teoria che suggeriva una riorganizzazione funzionale detta plasticità corticale del cervello. Studi successivi hanno scoperto che a seguito di un’amputazione la parte della corteccia relativa alla parte del corpo smetteva inizialmente di funzionare ma subito dopo riprendeva rispondendo a stimolazioni provenienti dalle aree adiacenti a quella amputata. In uno studio fatto su un soggetto a cui era stato amputato un braccio si riscontrò che nel momento in cui veniva passato un tampone di cotone sul volto esso sentiva qualcosa che gli toccava il braccio amputato. questo fenomeno è noto ed è chiamato sensazione dell’arto fantasma. Questo fenomeno si spiega con il fatto che una parte del corpo si è appropriata della vecchia superficie corticale dedita all’arto mancante. a questo punto i ricercatori hanno indagato se una pratica intensa potesse portare a una riorganizzazione del cervello adulto. Tale ipotesi è stata confermata in uno studio fatto con dei musicisti che suonavano il violino osservando che le aree relative alla mano con cui suonavano erano più intense rispetto al gruppo di controllo. Un altro studio ha preso in esame i giocolieri e avvisto che dopo una pausa di allenamento le aree del cervello tendevano a tornare alla loro dimensione iniziale. Un ruolo importante sulla riorganizzazione della corteccia motoria dipende dal livello di acido gamma aminobutirrico (GABA) il principale neurotrasmettitore inibitorio. Si è visto che quando i livelli di GABA sono alti l’attività di singoli neuroni corticali è stabile ma se il livello di GABA e basso i neuroni tenderanno a rispondere a una gamma di stimoli più ampia. Si è osservato che quando viene bloccato temporaneamente il flusso sanguigno di una mano si riscontrano livelli di GABA più bassi a livello cerebrale. Questo ha portato a pensare che il rilascio di un’inibizione tonica possa avvenire anche da fonti remote. E dunque predisporre a una plasticità cerebrale. 8 Visione: uno dei motivi per cui la visione è un senso così importante e dato dal fatto che ci permette di percepire informazioni anche a una certa distanza e dunque di impegnarci in quello che viene detto “telerilevamento”. Questo ci permette di evitare i predatori e di identificarli ad una certa distanza. L’informazione visiva è contenuta nella luce riflessa dagli oggetti per questo motivo i nostri recettori sensoriali rispondono alla luce riflessa. La luce attraversa il cristallino dell’occhio e l’immagine viene invertita e indirizzata sulla superficie posteriore dell’occhio detta retina. La retina è spessa 0,5 mm ed è formata da 10 strati di neuroni molto condensati. Negli strati più profondi troviamo milioni di fotorecettori che si suddividono in coni e i bastoncelli. I fotorecettori contengono foto pigmenti ovvero delle molecole proteiche che sono sensibili alla luce. I coni e bastoncelli non producono potenziali d’azione ma la decomposizione dei fotopigmenti altera il potenziale di membrana dei fotorecettori e innesca così potenziali d’azione nel neurone a valle. - I bastoncelli: contengono il pigmento rodopsina che si destabilizza da bassi livelli di luce. Per questo motivo questo tipo di fotorecettore è molto utile di notte. in realtà rispondono anche a luce molto brillante ma in questo caso il pigmento si esaurisce rapidamente e il bastoncello cessa di funzionare finché non viene rifornito tale processo richiede diversi minuti. - I coni: contengono un pigmento detto Fotopsina. Rispondono a livelli di luce più alti ma si rigenerano molto rapidamente per questo sono utili durante la visione diurna. Ci sono tre tipologie di coni suddivisi n base alla sensibilità alla lunghezza d’onda. alcuni sono più sensibili alle onde corte (blu) altri a quelle medie (verde) altri ancora a quelle lunghe (Rosse). I coni sono più addensati nella fovea mentre i bastoncelli sono distribuiti in tutta la retina. I coni e bastoncelli si connettono ai neuroni bipolari che creano sinapsi con le cellule gangliari. Gli assoni delle cellule tutte insieme formano il nervo ottico che trasmette l’informazione al sistema nervoso centrale. Prima di arrivare al nervo ottico l’informazione viene elaborata nel momento in cui da milioni di fotorecettori passa a soli 2 milioni di cellule gangliari sommando i loro input. questo avviene per i bastoncelli . La cellula gangliare innervata dei coni viene innervata da un numero minore per questo l’informazione e più specifica permettendo un’immagine più nitida. Ciascun nervo ottico si suddivide in due parti: - 1) ramo temporale (laterale) continua verso il lato ipsilaterale - 2) ramo nasale (mediale) degusta e proietta al lato controlaterale questo punto di incrocio si chiama chiasma ottico. Le fibre di ciascuna semiretina (parte interna e quella vicino alle tempie) si incrociano tutta l’informazione di un campo visivo destro o sinistro viene proiettata all’emisfero opposto. La via retinogenicolata È la proiezione che avviene dalla retina al nucleo genicolato laterale del talamo. Il nucleo genicolato laterale è formato da sei strati: - Un tipo di cellula gangliare; - La cellula M ( proietta ai due livelli più bassi); - Un altro tipo di cellula gangliare; - La cellula P (proietta ai primi quattro livelli). Il 10% delle fibre del nervo ottico va ad innervare strutture sottocorticali come il nucleo di pulvinar del talamo e il collicolo superiore del mesencefalo. Queste strutture giocano un ruolo importante nell’attenzione visiva. 9 La parte finale della proiezione alla corteccia visiva avviene attraverso la via Gianicolocorticale Ovvero la via che parte dal nucleo genicolato laterale e arriva alla corteccia visiva primaria del lobo occipitale. Ricapitolando l’informazione della corteccia è stata elaborata da almeno quattro neuroni diversi: fotorecettori, cellule bipolari, cellule gangliari, cellule del nucleo genicolato laterale. - Campo recettivo: I neuroni rispondono a degli stimoli visivi solo quando lo stimolo viene presentato in una regione specifica dello spazio che viene detto appunto campo recettivo. I campi recettivi di cellule vicine sono in relazione ordinata. - Mappe retinotopiche: sono rappresentazioni topografiche che prendono in considerazione le Mappe che si vengono a formare dei campi recettivi delle cellule che sono l’insieme del l’attributo esterno (localizzazione spaziale) e la rappresentazione neurale di quell’attributo. Una mappa retinotopica completa rappresenta l’intero emicampo controlaterale. I campi recettivi hanno una grandezza variabile normalmente sono più ristretti all’inizio del sistema visivo come ad esempio nel nucleo genicolato laterale dove rispondono solo se lo stimolo cade entro una regione dello spazio molto limitata; risultano leggermente più ampi ad esempio nella. Le cellule del lobo temporale ad esempio potrebbero includere un intero emicampo. 10 Le aree corticali visive: la corteccia visiva primaria detta anche corteccia striata e la regione iniziale di proiezione degli assoni genicolati. La presenza di numerose aree visive nella corteccia porta a pensare che elaborazione visiva sia di tipo gerarchico dunque ogni aria elabora l’informazione partendo dalle elaborazioni fatte nelle aree precedenti. Le cellule nella corteccia visiva primaria identificano i margini, le cellule più complesse delle altre aree visive identificano gli angoli e immagini poi le forme fino ad arrivare al culmine strutturando la rappresentazione dello stimolo così che esso possa corrispondere alle informazioni depositate in memoria.ma questa ipotesi di semplice gerarchia non permette una spiegazione completa infatti diversi partner di convergenza e divergenza dando luogo a una molteplicità di vie. Una seconda ipotesi è quella di un processo analitico ovvero ogni area visiva produce una mappa dello spazio esterno ma ogni aria è altamente sensibile a differenti tipi di informazioni come ad esempio il colore, il movimento. Le informazioni sarebbero il frutto di una strategia del tipo di video e conquista poiché invece di avere tutti gli attributi di un oggetto rappresentati in tutte le aree visive ogni aria produrrebbe una sua analisi specializzata. man mano che l’informazione avanza lungo il sistema visivo le diverse aree integrano le informazioni così da poter formare percetti riconoscibili. Ipotesi di aree specializzate è stata ulteriormente dimostrata da diversi esperimenti come ad esempio dagli esperimenti effettuati nell’aria V5 detta anche (MT poiché si trova nel lobo temporale mediale) le registrazioni sul cervello della scimmia in questa regione non hanno portato specifici riferimenti al colore dello stimolo le cellule rispondevano in maniera simile sia se lo stimolo era rosso o verde; rispondevano in modo molto sensibile invece al movimento e alla direzione. Prendendo in esame una singola cellula è stato dimostrato che questa rispondeva in modo maggiore quando lo stimolo veniva mosso verso il basso verso sinistra piuttosto che quando lo stimolo veniva mosso verso l’alto e verso destra. questo ha dimostrato che la cellula si attiva solo quando lo stimolo ricade all’interno del suo campo ricettivo e che la sua risposta sarà massima quando lo stimolo si muove in una direzione particolare. L’attività delle cellule in Q5 è correlata anche alla velocità di movimento questo è stato dimostrato osservando che la risposta maggiore si aveva quando lo stimolo veniva mosso in maniera rapida. I primi studi fatti nella corteccia visiva avevano dimostrato che le cellule in quest’area a differenza di quelle del nucleo genicolato laterale che avevano campi recettivi di forma circolare, erano sensibili ai margini. Ad esempio nelle cellule del nucleo genicolato laterale quando uno stimolo luminoso ricade entro la regione centrale la cellula si attiva, se cade nella periferia la cellula viene inibita e ancora se lo stimolo coinvolge sia il centro e la periferia non si riscontrerà una risposta cellulare poiché le due regioni si annullano a vicenda. Questo viene spiegato con il fatto che i nostri organi di senso sono interessati soprattutto al cambiamento. Le cellule della corteccia visiva primaria non possiedono un campo ricettivo tondo ma un centro in tre posizioni dello spazio leggermente diverso per questo motivo la cellula e selettiva per margini orientati in una certa direzione. Qualora lo stimolo non colpisse il campo recettivo con la giusta angolazione e dunque colpisse sia parte del campo che della periferia la cellula sarebbe inibita. Queste cellule sono state definite “cellule semplici” altri studi hanno visto che man mano che veniva spostato l’elettrodo lungo la corteccia l’orientamento preferenziale delle cellule variava in modo continuo. La selettività per l’orientamento si è rilevata una proprietà distintiva dei neuroni della corteccia visiva primaria. Prendendo in esame un pezzetto di corteccia i neuroni hanno campi recettivi incentrati in regioni dello spazio molto simili. I neuroni si dispongono in colonne alternate di cellule sensibili agli input provenienti dall’occhio destro e sinistro. in questo modo è possibile una rappresentazione dello spazio esterno definendone i margini. - Cellule Interblob: semplici e complesse, selettive all’orientamento, alla direzione; tendenzialmente non sensibili alla lunghezza d’onda. 11 - Cellule blob: campo recettivo circolare con diversi tipi di opponente acromatica; monoculari e sensibile alle lunghezze d’onda contengono la maggior parte dei neuroni sensibili al colore non sono selettive all’orientamento e alla direzione. Sono dei siti di convergenza degli imput ( magno; parvo; Konio) Utilizzando tecniche di neuro immagine I ricercatori con i primi studi tentarono di descrivere l’architettura funzionale del cervello umano.1 primo studio fatto da Zeki utilizzo la PET per analizzare il sistema visivo umano. Il suo obiettivo era quello di identificare le aree coinvolte nell’elaborazione dell colore e del movimento. utilizzò il metodo sottrattiva andando a sottrarre l’attivazione registrata nella condizione di controllo a quella sperimentale. Nella sperimentazione sul colore gli osservanti osservavano dei rettangoli acromatici che possedevano un diverso tono di grigio e un’ampia gamma di luminanza. Nella condizione sperimentale le macchie grigie venivano sostituite da una varietà di colori. Ogni aria colorata era abbinata alla stessa aria grigia con luminanza uguale.i n questo modo i neuroni specifici per la luminanza si dovevano attivare in ugual misura. Stimolo colorato portava ad un momento di attività nella regione neurale sensibile all’informazione cromatica. Una volta avuti i risultati di entrambi i gruppi si procedeva con la sottrazione. un altro esperimento venne fatto sul movimento. i risultati di questi studi portarono ad osservare che i due compiti attivavano diverse aree cerebrali. I quando analizzavano i sette colori notavano aree bilaterali situate nelle regioni più anteriori e inferiori del lobo occipitale. durante la sperimentazione sul movimento invece si evidenziavano aree residue bilaterali localizzate vicino alla giunzione tra la corteccia temporale, parietale e occipitale. Così area visiva quattro l’area attivata dal colore e aria visiva cinque lare attivata dal movimento. l’aria V5 viene detta anche Mt umana anche se nel cervello umano non si trova nel lobo temporale. Le Mappe che uscirono dagli studi con la PET erano piuttosto grossolane per questo motivo di recente lo studio sull’organizzazione della corteccia visiva umana si è basato su tecniche più sofisticate come l’ FRMI in cui un partner semicircolare a scacchiera viene fatto ruotare lentamente intorno al centro del video. I dati che vennero estrapolati vennero poi elaborati su una mappa cerebrale bidimensionale utilizzando dei codici a colori in cui colori simili indicano area attivate negli stessi erano presenti negli studi fatti sulle scimmie. E queste aree sembrano attivarsi quando segnali di movimento definiscono i margini degli oggetti come ad esempio quando uno stimolo in 12 movimento copre lo sfondo. Dalle mappe si vede anche in che modo l’eccentricità: la distanza dal punto di fissazione, viene rappresentata nelle aree visive. Eccentricità: distanza radiale dal centro della visione (Fovea) alla periferia. L’acuità visiva è maggiore nella fovea a causa della sproporzionata quantità di corteccia che codifica l’informazione proveniente da questa parte dello spazio. Dalla sensazione alla percezione: la prima cosa che gli studiosi hanno cercato di capire e dove si trovano i percetti. Per capirlo hanno utilizzato le illusioni in modo da capire quando e dove l’elaborazione degli stimoli porti a fuorviare i segnali.ad esempio quando vediamo giallo un disco che in realtà cambia colore ad intermittenza da rosso a verde. Questo fenomeno è detto fusione del lampeggio intermittente. quello che si chiesero gli scienziati e a che livello questa fusione avviene. confrontarono le risposte BOLD nelle diverse arie della corteccia visiva V1; V4 e VO modificando il livello di intermittenza e il contrasto. L’elaborazione sottocorticale di alcune aree di ordine inferiore come V14 erano in grado di distinguere l’intermittenza a cinque Herz e a 30 Herz contrariamente le risposte bald nell’aria visiva adiacente a V4 VO non era in grado di differenziare tra lo stimolo ad intermittenza maggiore e lo stimolo di controllo statico. Dunque l’illusione viene creata nelle aree superiori conosciute come VO e V8 questo porta a pensare che la percezione cosciente avvenga in arie superiori. Un altro studio che prese in esame la velocità con cui uno stimolo veniva presentato cerco di indagare la percezione non cosciente si dimostrò che le aree di Bruno erano in grado di rappresentarlo ed identificarne l’orientamento ma tale informazione veniva persa già in V2 e V3. Questo dimostra ulteriormente che le regioni sensoriali primarie sono strettamente legate allo stimolo fisico mentre la nostra esperienza percettiva dipende 13 maggiormente dalle regioni sensoriali secondarie e associative. Un altro esperimento è stato quello del partner dell’enigma in cui illusione è data dal movimento non vero dei cerchi blu.noi sappiamo che è un movimento produce una risposta in Q5 quello che si sono chiesti i scienziati e se anche il movimento illusorio attivi quest’aria. Studi con la PET e con la risonanza magnetica hanno dimostrato un’effettiva attivazione di V5. Differenze individuali nella percezione: quando si guarda un’illusione insieme ad un’altra persona non si avrà la stessa reazione. Una delle illusioni classiche e quella presentata nella figura in cui nonostante le persone percepiscono il cerchio centrale di destra maggiore di quello di sinistra, la percezione di grandezza varia dal 10% fino a un 50%. queste differenze hanno portato i ricercatori ad indagare il perché.si è constatato che tanto più l’area visiva primaria era piccola tanto più era maggiore illusione percepita. questa correlazione non è stata invece trovata in V2 e V3. Un’ipotesi è data dal fatto che nelle persone con V1 più grande vengano influenzate meno dai cerchi i vicini a quello centrale e in quanto hanno una rappresentazione ad alta risoluzione dello spazio. Le differenze di percezione possono essere date anche da dei deficit come avviene ad esempio nelle persone daltoniche. queste persone non hanno uno dei tre fotorecettori. Vengono definiti “dicromati” può essere anche che abbia tutti e tre foto pigmenti ma che funzionino in maniera anomala in questo caso vengono definiti “tricromati anomali” I deficit di percezione dei colori dovuti a disturbi del sistema nervoso centrale sono molto rari e vengono detti (acromatopsia). 14 Questo deficit è associato a lesioni che comprendono V4 e la regione anteriore di V4. I neuroni sensibili ai colori sono anche selettivi per l’orientamento per questo motivo molti hanno difficoltà a percepire la forma. Uno dei test più utilizzati per riscontrare questo fu il test della percezione della forma.dei risultati che si ottennero appare chiaro come V4 non è solo l’area del colore ma un’aria in cui si possono ricavare informazioni che aiutano a definire i margini che separano gli oggetti dal nostro ambiente visivo. Achinectopsia: perdita selettiva della percezione del movimento. Gli studi partirono da un soggetto a cui vennero fatte delle scansioni con la tac che rilevarono lesioni bilaterali che coinvolgevano le cortecce tempo parietali. le lesioni riguardavano porzioni posteriori e laterali del giro temporale medio. Negli anni si è riscontrato che lesioni unilaterali portano a deficit più lievi. questo potrebbe far pensare che il movimento viene percepito finché V5 rimane intatta almeno in uno degli emisferi. Questo portò a studiare come la corteccia in Q5 reagiva quando veniva presentata una serie di stimoli in movimento. per rendere il tutto più complesso solo alcuni stimoli si muovevano nella direzione del target mentre gli altri in direzioni casuali inoltre il target era preceduto e seguito da stimoli di mascheramento. la direzione dello stimolo era visibile solo in una breve finestra di tempo. I scienziati utilizzarono la stimolazione magnetica transcranica (TMS) E notarono che le prestazioni venivano compromesse in due distinti intervalli di tempo. il primo si verificava quando lo stimolo veniva applicato circa 100 ms prima dell’inizio il secondo si verificava quando lo stimolo è rappresentato 150 ms dopo lo stimolo. Per quanto riguarda il secondo. I ricercatori lo spiegarono affermando che l’impulso aggiungeva rumore alla rappresentazione in V5. Il secondo deterioramento sarebbe provocato da un effetto indiretto dell’impulso sull’attività in V1. Percezione senza corteccia visiva: la maggior parte degli assoni a scende dal nucleo genetico lato laterale e termina nella corteccia visiva primaria. Per questo motivo si aspetta che un individuo con lesione alla corteccia primaria presenti cecità. La cecità può essere incompleta nel caso in cui la lesione è strettamente collegata a una parte del campo visivo. Questo deficit è detto Emianopsia. Le regioni di cecità vengono dette scotomi. Ma poiché le vie della visione oltre alle vie corticali presentano anche delle vie sottocorticali gli esperti sono riusciti ad ottenere dei risultati sorprendenti dimostrando che le capacità visive possono persistere anche in assenza della corteccia visiva primaria. Il 10% delle proiezioni infatti va verso i nuclei sottocorticali, la proiezione 15 più importante va verso il collicolo superiore. Il collicolo superiore svolge un ruolo fondamentale nella produzione dei movimenti oculari, la simulazione in questo punto può provocare dei movimenti oculari, la direzione dei quali dipende dal sito della stimolazione. È interessante capire come il collicolo superiore rappresenti il mondo visivo.uno degli studi più famosi fu quello su alcuni criceti che vennero addestrati a seguire due compiti. Nel primo (orientamento) i criceti venivano addestrati a raccogliere semi di girasole disposti in diverse porzioni dello spazio; nel secondo compito (discriminazione) i criceti erano istruiti a percorrere uno dei corridoi verso una porta che aveva strisce o verticali od orizzontali. Si è dimostrato che lesioni al collicolo influivano sulla prestazione del compito di localizzazione mentre lesioni alla corteccia visiva influivano Sulla prestazione del compito di discriminazione. Percezione multimodale: l’esperienza che ci circonda è una costruzione multisensoriale unitaria. Molto di quello che proviamo risulta da ciò che ci aspettiamo dall’esperienza. La percezione non è un processo frammentario ma è il risultato di una rappresentazione coerente del mondo che proviene da tutti gli organi di senso. Come avviene nell’effetto “ MCGurk” Dove ciò che si sente è influenzato dal movimento delle labbra. O durante gli esperimenti sulla mano di gomma che viene interpretata come propria. gli scienziati hanno cercato di capire come il cervello riesca ad integrare l’informazione proveniente da diversi sensi. Per molto tempo si è pensato che alcuni sensi dominassero su altri recentemente invece si pensa che il cervello riesca a combinare gli input provenienti dai diversi sensi e in base a proprietà esterne dell’informazione (localizzazione di un suono o di un’esperienza tattile). Pesi l’affidabilità di ogni senso, faccia una stima e prendo una decisione basata su quest’informazione e sulla proprietà esterna in questione. Nella maggior parte dei casi il cervello giudica l’informazione proveniente dalla vista più affidabile ma non è sempre così.ad esempio se un lampo visivo viene proiettato insieme due segnali acustici i partecipanti riportano che il lampo a lampeggiato due volte. questa illusione e detta guida uditiva. I neuroni che rispondono a più di un senso sono detti multisensoriali e si trovano in diverse regioni del cervello sia sottocorticali e corticali. - Sottocorticale: collicolo superiore del mesencefalo molti studi hanno dimostrato che in questa zona sono presenti diversi neuroni che si attivano maggiormente quando gli input derivano da molteplici sensi rispetto a quando l’input proviene da una singola modalità. In quest’aria sono presenti anche diverse mappe topografiche ordinate visive, uditive e somatosensoriali. I segnali multisensoriali vengono considerati dal cervello come più affidabili rispetto ai segnali provenienti da un singolo canale sensoriale. L’integrazione richiede che i diversi stimoli coincidano nello spazio nel tempo. - Corticale: l’attività multisensoriale viene osservata anche in molte regioni corticali come ad esempio nel solco temporale superiore. i ricercatori hanno dimostrato che nel solco temporale superiore delle scimmie vi erano una risposta agli stimoli visivi, uditivi e somatosensoriali. Attraverso indagine con la risonanza magnetica funzionale è stato visto come alcune aree del solco temporale superiore sinistro degli esseri umani si attivino quando i soggetti sono impegnati a leggere le labbra ma non quando i suoni non corrispondono ai movimenti delle labbra. altre regioni multisensoriali sono i lobi parietali e occipitali (ippocampo) in diversi studi si e cercato di comprendere come avvenisse l’interazione visiva ed uditiva, capendo se i due sistemi si trovano in competizione l’uno con l’altro oppure riescono ad integrare le informazioni. L’idea di quale sia la via neurale che permette una sensazione multisensoriale ha generato molto clamore tra i scienziati producendo diverse ipotesi. - La prima è la più radicale e crede che l’intera neuro corteccia sia multisensoriale e che l’integrazione avvenga nella zona Sub corticale. Esistono degli imputo e multisensoriali 16 diretti alla corteccia che provengono dal talamo. Ma sappiamo che è una lesione alla corteccia visiva primaria produce una grave cecità ma non ha un effetto sugli altri sensi. - Un’altra versione e che la corteccia possiede delle aree sensoriali specifiche che contengono alcuni interneuroni multisensoriali. - Un’altra versione ancora afferma che l’integrazione multisensoriale coinvolge proiezioni che si originano in aree corticali specifiche. Le proiezioni multisensoriali andrebbero da una regione sensoriale all’altra permettendo così una modulazione veloce nelle regioni primarie secondarie. - Un’altra versione ancora afferma che le sensazioni multisensoriali provengono da aree corticali specializzate per l’elaborazione multisensoriale come la corteccia parietale posteriore E solco temporale superiore. Sinestesia: fenomeno nel quale i sensi sono mescolati.si caratterizza per l’unione idiosincrasica tra le modalità sensoriali. le parole possiedono un gusto o la musica possiede dei colori. Essendo ricorrente all’interno di famiglie si pensa abbia una base genetica. La forma di sinestesie è più studiata e quella di grafemi colorati in cui le persone potrebbero affermare di vedere la lettera a come rossa e così via. Per cercare di studiare tale patologia si utilizza il compito di Stroop osservando se i colori delle lettere sono congruenti oppure no con la gamma sin estetica del soggetto. Studi con immagini cerebrali hanno dimostrato che l’esperienza multisensoriale Cina estetica si verifica lungo vari stadi sulla via visiva. Ad esempio quando ascoltano delle parole si vede un aumento dell’attività di V4 o ancora nel solco temporale superiore. Delle ricerche si sono domandati se i soggetti sinestetici presentino maggiori connessioni rispetto ai non sinestetici.usando immagini con tensore di diffusione è stato osservato che vi è un’aumentata connettività della sostanza bianca nei soggetti sinestetici. Riorganizzazione percettiva: in studi con la PET si è dimostrato che si verifica un notevole grado di riorganizzazione funzionale delle aree cerebrali.in un esperimento considerazione due gruppi uno formale e uno con una mancanza congenita della vista e vennero sottoposti a due condizioni sperimentali.la prima veniva chiesto loro di strofinare le dita su una superficie ruvida e coperta con puntini, nella seconda con dizione venivano somministrati dei compiti di discriminazione tattile (decidere se due solchi sulla superficie fossero uguali o diversi). Venne poi osservato il flusso sanguigno nella corteccia visiva durante le diverse prove. Nei partecipanti vedenti fu trovato un calo dell’attività nella corteccia visiva primaria, nei non vedenti l’attivazione della corteccia visiva primaria aumentava durante i compiti di discriminazione ma solo quando utilizzavano attivamente l’informazione tattile. Un altro studio ha studiato gli effetti della plasticità corticale in persone vedenti che erano state deprivate delle informazioni visive per un lungo periodo. i partecipanti rimasero cinque giorni bendati e furono sottoposti ad un addestramento intensivo sul Braille. Al quinto giorno riuscirono a discriminare in modo migliore rispetto a coloro che non erano stati bendati. immagini con Fm RI mostravano un’attivazione della corteccia visiva durante la stimolazione tattile dei polpastrelli. Dopo 20 ore dalla rimozione della benda però l’attivazione della corteccia visiva durante la stimolazione tattile scomparve questo dimostra che vi è una riorganizzazione rapida della corteccia. - Emiacromatopsia: centro del senso cromatico (giri linguale e fusiforme) - Achinetopsia: grave lesione MTG bilaterale - Regioni multisensoriali della corteccia: solco temporale superiore e posteriore, parietale posteriore, corteccia premotoria, corteccia prefrontale inferiore. 17 Cap 7 L’ATTENZIONE L’attenzione permette una percezione integrata. senza attenzione la percezione si dividerebbe e non ci sarebbe nessuna percezione è coerente del mondo. L’attenzione rappresenta un processo per cui la mente sceglie tra vari stimoli sensoriali in un dato momento consentendo solo ad alcune informazioni di arrivare alla coscienza. Attenzione selettiva: è il processo secondo il quale si processano informazioni rilevanti e se ne ignorano altri. l’attenzione selettiva può essere dall’alto al basso. Prestare attenzione alla pagine da studiare(top Down) o dal basso verso l’alto, diretto allo stimolo ad esempio rumore improvviso (Bottom Up). Modelli dell’attenzione: attenzione volontaria (prestare attenzione intenzionalmente); attenzione riflessa-autonoma (uno stimolo cattura la nostra attenzione dal basso verso l’alto). Sindrome di Balint: danno bilaterale regioni posteriori corteccia parietale e occipitale. Il soggetto può prestare attenzione solo ad un oggetto o un o un piccolo sottogruppo di oggetti. non è in grado di vederli tutti insieme e dunque di descrivere la loro posizione rispetto a un altro oggetto. William James al termine del XIX secolo sottolineò il concetto che l’attenzione ha una capacità limitata. L’attenzione influenza il modo in cui le persone codificano gli input sensoriali, li immagazzinano in memoria e li elaborano a livello semantico. I meccanismi che determinano dove e su cosa venga focalizzato la nostra attenzione sono detti i meccanismi di controllo attentivi, comprendono reti celebrale diffuse ma altamente specifiche. Anatomia: - Strutture sottocorticali: - collicolo superiore del mesencefalo – pulvinar - Strutture corticali: porzioni della corteccia frontale, parietale posteriore, corteccia temporale superiore e posteriore. - Strutture cerebrali mediali: corteccia cingolata anteriore, corteccia ci colata posteriore, insula. - Regioni sensoriali corticali, coinvolte poiché l’attenzione influenza in modo in cui l’informazione sensoriale viene elaborata dal cervello. Molto di ciò che i neuroscienziati conoscono è stato ottenuto esaminando pazienti che presentavano dei danni cerebrali che portavano a disturbi dell’attenzione. 18 Negletct: ha origine da un danno alla rete attentivi a celebrare di un solo emisfero. Spesso provocato da un ictus. nonostante ciascun emisfero possa essere danneggiato gli effetti più gravi si evidenziano quando ad essere interessato è quello destro. Pazienti con neglette all’emisfero destro possono comportarsi come se gli oggetti a sinistra non esistessero. in gergo tecnico si tende ad affermare che i pazienti tendono ad omettere l’informazione. Si possono osservare gli effetti del neglette su dipinti di un pittore tedesco che fu colpito a 67 anni da un grave ictus. I quadri dimostrano come egli era incapace di rappresentare la porzione dello spazio controlaterale. La sindrome non porta a problemi di vista ma comporta deficit dell’orientamento dell’attenzione e dell’azione in direzione opposta al lato del danno cerebrale unilaterale. un modo per osservare questo fenomeno è quello di osservare i movimenti oculari nei pazienti affetti da questa sindrome. Tali pazienti tendono a effettuare movimenti oculari nella direzione del campo visivo destro mentre gli altri pazienti mettevano in atto dei movimenti oculari dirigendoli nell’intero spazio sia a destra che a sinistra. Per porre una diagnosi di neglette si possono utilizzare il test della bisezione the line: i pazienti ricevono un foglio di carta sul quale sono presenti molte linee orizzontali e viene chiesto loro di dividerle esattamente a metà segnando una linea verticale. I pazienti con neglette tenderanno a segnare la linea verticale più spostata verso destra rispetto alla linea mediana. Un altro test potrebbe essere quello di far copiare loro un disegno come un fiore o di un orologio. tenderanno a disegnarne solo metà. Gli scienziati fecero degli esperimenti per capire anche se il neglette coinvolgesse la memoria. chiesero ai loro pazienti di ricordare e di descrivere la piazza del Duomo immaginandosi di trovarsi sui gradini. notarono che i partecipanti tendevano ad omettere le informazioni che si trovavano sul lato della piazza controlaterale alla loro lesione. Quando poi venne chiesto loro di immaginarsi nella piazza e di descrivere il Duomo riferivano prima avevano tralasciato. questo dimostrò che il neglette spaziale si ritrova anche nella memoria visiva durante la rievocazione. L’esame dei campi visivi dimostra che i pazienti affetti da neglette rilevano gli stimoli in modo normale quando gli stimoli sono rilevanti e vengono presentati singolarmente. quando vengono invece presentati più stimoli in maniera simultanea questi entrano in competizione nell’emicampo ipsilaterale impedendo così al paziente di rilevare quello controlesionale. Questo fenomeno è noto come estinzione. Questa condizione viene descritta con una tendenza pilotata piuttosto che come una perdita delle capacità di focalizzare l’attenzione in direzioni contro lesionali. Sindrome di Balint: questi pazienti mostrano tre deficit caratteristici di tale disturbo: 1) simultanagnosia: difficoltà a percepire il campo visivo come una scena unitaria.vi è una cecità per stimoli presentati simultaneamente; 2) Aprassia oculare: vi è un deficit nell’esecuzione dei movimenti oculari (saccadi) che non permette al soggetto di prestare attenzione e dunque di scansionare il campo visivo. 3) Atassia ottica: difficoltà nei movimenti manuali diretti visivamente. questi pazienti qualora gli venisse chiesto di prendere l’oggetto avrebbero molte difficoltà. Sebbene in entrambe le patologie vi siano dei disturbi della percezione e anche vero che ciascuno dei due disturbi presentare aree danneggiate del cervello diverse.nel neglette si trovano lesioni unilaterali della corteccia parietale, temporale posteriore e di quella frontale. oppure danni sottocorticali ai gangli della base, al talamo e al mesencefalo. I pazienti che invece soffrono della sindrome di Balint presentano lesioni occipitoparietalo bilaterali. 19 Modelli dell’attenzione: quando le persone rivolgono l’attenzione su un oggetto viene definito orientamento. Nell’attenzione selettiva l’orientamento può essere volontario o riflesso. L’orientamento attentivo può essere esplicito quando si volge lo sguardo verso lo stimolo. Oppure implicito quando non è accompagnato da movimenti orientati del corpo. Attraverso l’esperimento di Helmotz (tabellone lettere) si dimostrò che mediante una decisione volontaria anche in assenza di movimenti oculari e di cambiamenti di accomodazione è possibile concentrare l’attenzione sulle sensazioni provenienti da una parte particolare del nostro sistema nervoso periferico e allo stesso tempo escludere l’attenzione da tutte le altre parti. Effetto cocktail party: la capacità di attenzione uditiva selettiva che consente di partecipare ad una conversazione in un bar o in un party affollati mentre signora il resto dei suoni intorno. Cherry mise in atto degli esperimenti in cui chiedeva ai partecipanti di indossare delle cuffie. Nelle cuffie venivano trasmessi segnali differenti (ascolto dicotico) . Chiese ai partecipanti di prestare attenzione e di ripetere l’enunciato verbale di uno dei due orecchi ignorando contemporaneamente l’input che arrivava nell’altro orecchio. Quando il messaggio era diverso notò che i partecipanti non riuscivano a riferire nessun dettaglio sull’input ignorato. Questo portò Cherry ad affermare che l’attenzione volontaria influenza ciò che viene elaborato in quanto da una parte focalizza l’attenzione codificandone in maniera migliore gli input che arrivano ad un orecchio dall’altra però vi è una perdita o degradazione degli impulsi inviati all’altro orecchio. Ciò venne spiegato da James come una limitazione a collo di bottiglia dell’elaborazione dell’informazione agli stadi di analisi percettiva a capacità limitata. Dunque sarebbero soltanto input ad alta priorità che si è selezionato ad essere elaborati. Gli effetti sull’attenzione nell’elaborazione sensoriale hanno portato i neuroscienziati a chiedersi se questi passaggi avvengono in maniera precoce o in maniera tardiva. Ovvero chiedersi se il cervello elabora fedelmente tutti gli input sensoriali in entrata per creare una rappresentazione del mondo esterno oppure processi come la tensione possono influenzare l’elaborazione sensoriale. Lo psicologo inglese Donald Broadbent ipotizzò che il sistema di elaborazione delle informazioni sia caratterizzato da uno o più Stati di capacità limitata attraverso il quale può passare solo una certa quantità di informazioni. e dunque gli input che arrivano ai livelli superiori di elaborazione del cervello devono subire una selezione a livello inferiore. verranno fatti passare solamente quelli più importanti sui quali si è focalizzata l’attenzione. lo psicologo definì questo tipo di meccanismo simile ad un filtro che poteva lasciar passare l’informazione è considerata rilevante dei meccanismi attentivi e bloccare l’informazione ignorata. Era convinto anche della selezione precoce. Altri modelli che seguono la selezione tardiva ipotizzano invece che tutti gli stimoli siano elaborati in modo uguale dal sistema percettivo e che la selezione avvenga con la funzione di determinare quali di essi verranno sottoposti a un’elaborazione ulteriore e quali arriveranno a essere rappresentati nella consapevolezza cosciente. tali processi indicano dunque che I processi attentivi non influiscono sulla nostra analisi percettiva degli stimoli, in quanto la selezione avverrebbe in stadi più alti dell’elaborazione dell’informazione. I modelli di selezione precoce (tutto o niente) si scontrarono un problema in quanto si osservò che in alcuni esperimenti sul cocktail party informazioni salienti dell’orecchio ignorato venivano percepite ad esempio quando si utilizzava il nome dell’ascoltatore. per questo si arrivò a pensare che l’informazione del canale ignorato non venisse del tutto escluso ma soltanto degradato o attenuato questo permette ad alcuni input di arrivare ad essere elaborati a livelli superiori per quanto il segnale sia attenuato. per accertarsi di questo vennero messi in atto metodi che includevano l’analisi cronometrica ovvero l’analisi del decorso del tempo dell’elaborazione dell’informazione con livello di risoluzione del millisecondo. Un altro modo per studiare l’effetto dell’attenzione su l’elaborazione dell’informazione e quello di esaminare i soggetti sottoposti a stimoli-bersaglio in diverse condizioni di attenzione.uno dei 20 metodi più popolari consiste nel fornire segnali che indirizzano l’attenzione del partecipante verso una particolare posizione dello spazio o una caratteristica del target prima di presentare lo stimolo target è rilevante per il compito. questi compiti vengono detti “compiti di cuing” in uno degli esperimenti più importanti i partecipanti ricevono istruzioni relative al fatto che nonostante un suggerimento esempio una freccia indichi la posizione più probabile di un target devono rispondere al target ovunque esso appaia. questo è noto come cuing endogeno. In questa condizione l’attenzione è orientata verso il suggerimento dal rispetto volontario da parte del partecipante e dal significato del suggerimento piuttosto che Dalle semplici caratteristiche fisiche. Quando il suggerimento suggerisce la posizione del Target questi casi vengono dette prove valide; quando non suggerisce la posizione veritiera si parla di prove invalide; quando non suggerisce alcuna posizione del Target viene detta prova neutra. Nelle prove valide i partecipanti rispondono più velocemente di quando vengono somministrati suggerimenti neutri. questo dimostra i benefici dell’attenzione sulla velocità. inoltre ci sono riscontrati tempi più lenti quando il suggerimento non predice la posizione del target nelle risposte invalide sottolineando i costi dell’attenzione. La differenza dei tempi di risposta viene spiegata dal Cunnie considerando l’attenzione spaziale come un fascio di luce che viene indirizzata in maniera cosciente a piacimento della persona o che può essere richiamata in maniera riflessa dei venti sensoriali salienti. Bisogna considerare che sebbene la maggior parte degli esperimenti ricada su un’attenzione visiva in verità essa opera in tutte le modalità sensoriali. dopo aver studiato le conseguenze sulle modalità uditive si vuole studiare quelle sull’elaborazione visiva capendo se l’attenzione influenzasse l’elaborazione visiva, quando e dove e se durante l’elaborazione avvenisse l’influenza sul sistema visivo. Attenzione spaziale volontaria: - Effetti dell’attenzione corticale: i meccanismi neurali dell’attenzione selettiva visiva sono stati indagati utilizzando i metodi dei paradigmi di cuing in un esperimento i partecipanti devono prestare attenzione in modo non esplicito ovvero non togliendo lo sguardo dal punto di fissazione centrale, a stimoli presentati in una posizione dello spazio ignorando altri stimoli presentati in un’altra posizione dello spazio. durante l’esperimento si procede con la registrazione dei potenziali correlati agli eventi (ERP) La tensione spaziale è un effetto più precoce sull’elaborazione dello stimolo, questo può essere dato anche dal fatto che la mappa retino topica del sistema visivo comporta che il cervello codifichi lo spazio molto precocemente già a livello di retina. Si considera che +1 rappresenta l’onda sensoriale generata dall’attività neurale nella corteccia visiva e conseguentemente la sua sensibilità all’attenzione spaziale sostiene in modelli della selezione precoce dell’attenzione. Come sappiamo però gli ERP registrano un insieme di scariche alcuni neuroscienziati hanno indagato invece in che modo l’attenzione selettiva visiva spaziale influisse sui tassi di scarica di singoli neuroni nella corteccia visiva delle scimmie. Prima registrarono e catalogarono le risposte dei singoli neuroni nell’area visiva 21 quattro per capire a quali stimoli rispondessero in maniera più vigorosa. Successivamente presentarono in maniera simultanea lo stimolo preferito e quello non preferito, gli stimoli erano presentati vicini nello spazio così da farli rientrare in regioni del campo visivo che definiva il campo recettivo del neurone. I per mesi hanno istruito la scimmia a fissare un punto specifico del monitor così da indirizzare l’attenzione in modo non esplicito. Vennero così registrate e confrontate le risposte dei singoli neuroni nelle due condizioni e si osservò che quando veniva presentata attenzione allo stimolo preferito questo generava una risposta più forte rispetto a quando lo stimolo preferito veniva ignorato poiché veniva presentato quello meno preferito. questo dimostrò che l’attenzione influisce sui tassi di scarica dei neuroni in V4. Questi studi sono stati fatti anche in altre aree visive incluse aree più tardive della via ventrale nella regione infero temporale. altri lavori sulla via dorsale ha dimostrato effetti dell’attenzione anche nelle aree dell’elaborazione del movimento MT e MST. Altri studi si sono concentrati nel capire se l’attenzione potesse influenzare anche l’area visiva primaria. Alcuni scienziati hanno insegnato a delle scimmie a fissare un punto centrale di uno schermo e a prestare attenzione non esplicita a un input di… Rumorosi bianchi e neri sfarfallamenti. Per rilevare un piccolo puntino colorato che poteva comparire in un punto qualsiasi dello schermo. Dopo l’apparizione del puntino dovevano indicare il punto con un rapido movimento oculare. la posizione attesa poteva essere posizionata nel campo ricettivo del neurone di uno o nel campo visivo opposto. gli studiosi registravano un singolo neurone. Ottennero come risposta che l’attenzione spaziale accresceva le risposte delle cellule semplici ma non influenzava l’organizzazione spaziale dei loro campi recettivi. Sugli esseri umani sono state condotte delle ricerche mettendo in relazione i risultati degli ERP con la neuro anatomia celebrale funzionale E combinando le immagini con la PET. Hanno dimostrato che l’attenzione viso spaziale modula il flusso sanguigno della corteccia visiva. Studi successivi che hanno utilizzato FMRI portarono a vedere attivazione della corteccia visiva controlaterale all’emicampo attenzionato. Vennero create delle mappe retino topica ed usate in un compito di attenzione spaziale in cui i partecipanti dovevano prestare in maniera selettiva attenzione agli stimoli localizzati in un determinato quadrante del campo visivo e ignorare quelli presentati negli altri quadranti. I ricercatori hanno trovato che l’attenzione spaziale produceva modulazioni dell’attività in molteplici aree visive della corteccia extra striata e modulazioni più piccole nella corteccia striata V1. dunque la tensione spaziale influenza della bora azione di input visivi in quanto gli stimoli attesi producono risposte neurali maggiori di quelle degli stimoli ignorati. tale differenza è osservabile In diverse aree visive corticali. Per capire i diversi effetti dell’attenzione sulle diverse aree corticali si sono utilizzate immagini provenienti dalla risonanza magnetica funzionale in cui si osservato che l’attività in buono sembra essere meno robusta di quella della corteccia extra striata. Bisogna capire quali siano i meccanismi che potrebbero spiegare l’organizzazione gerarchica degli effetti dell’attenzione. Per comprendere questo si è postulato il modello della competizione pilotata dell’attenzione selettiva: tale modello potrebbe spiegare perché gli effetti dell’attenzione risultino maggiori quando all’interno del campo recettivo di un neurone rientrino molti stimoli in competizione E come l’attenzione influisca in modi diversi nella gerarchia visiva in base al variare delle proprietà dei campi recettivi neurali. Il modello dunque afferma che nel momento in cui all’interno del campo ricettivo di un neurone si presentino due stimoli l’attenzione può aiutare a risolvere la contesa a favore dello stimolo atteso. più si sale nella via visiva maggiore sarà il campo recettivo dei neuroni e dunque maggiore sarà la competizione tra stimoli diversi questo porterà ad aver bisogno di maggiore attenzione per aiutare a risolvere la contesa. Dunque l’attenzione è focalizzata su uno stimolo attenua l’influenza dello stimolo in competizione. Le differenze che si riscontrano in V1 rispetto che in V4 possono dipendere sia dalla differenza di 22 ampiezza del campo ricettivo sia dalla distanza dell’input dall’osservatore. Alcuni ricercatori unendo i dati provenienti dagli ERP, dalla magnetoencefalografia e dalla risonanza magnetica funzionale hanno proposto ad alcuni soggetti dei bersagli; alcuni bersagli erano composti da un quadrato e altri da un gruppo di quadrati di piccole o grandi dimensioni di colore rosso e verde che si muovevano sopra e sotto. L’indicazione era quella di prestare attenzione a un target di un certo colore e al suo spostamento. Lo studio ha rilevato che l’attenzione agiva a livelli precoci del sistema visivo per i Target più piccoli rispetto ai target più grandi dunque questo porta a pensare che vi sia un’ottimizzazione per adeguare l’estensione spaziale degli stimoli visivi del compito. - Effetti dell’attenzione sottocorticale: noi sappiamo che parte del nervo attico entra in contatto sinaptico con i neuroni del nucleo reticolare talamico (porzione del nucleo reticolare che circonda il corpo genicolato laterale). I risultati negli studi mostrarono che l’attivazione del nucleo genicolato laterale nella corteccia visiva controlaterale era maggiore per i target attesi rispetto all’attività misurata in risposta ai target ignorati. Questo mostra che un’attenzione “intensamente focalizzata” può modulare l’attività del talamo. Altri studi hanno osservato che quando lo stimolo cadeva il campo recettivo del neurone del nucleo genicolato laterale il tasso di scarica del neurone aumentava di conseguenza la scarica dei neuroni circostanti del nucleo reticolato talamico diminuiva. Questo può essere spiegato con l’esistenza di segnali attivanti e inibitori del nucleo genicolato laterale alla corteccia visiva attraverso i circuiti del nucleo reticolato talamico. quando un segnale neurale discende la corteccia o va verso i neuroni del nucleo reticolato talamico questi possono eccitare i neuroni di questo nucleo inibendo quelli del nucleo genicolato laterale oppure sopprimere l’attività dei neuroni di questo nucleo aumentando di conseguenza la trasmissione di quelli del nucleo genicolato laterale. Questi studi dimostrano che l’attenzione spaziale intensamente focalizzata può modulare l’attività agli stadi più precoci del sistema visivo, nei nuclei di relais sottocorticali del talamo. Attenzione spaziale riflessa: avviene quando viene attivata da stimoli che risultano in qualche modo vistosi.più è saliente è lo stimolo più la nostra attenzione verrà catturata. Molte volte capita di girare la testa ancora prima di riuscire a fermare il movimento perché la nostra attenzione riflessa ci induce a orientarci in modo manifesto verso lo stimolo sensoriale (esplicito) ma può 23 succedere anche che non sia esplicita ma comunque venga attratta verso stimoli sensoriali. Quello che gli scienziati si chiesero e se l’attenzione riflessa e quella volontaria siano elaborate nel medesimo modo. Gli effetti dell’attenzione riflessa possono essere misurati mostrando in che modo un lampo di luce irrilevante ai fini del compito influenzi la velocità delle risposte a stimoli bersaglio rilevanti per il compito presentati successivamente. Questo metodo è indicato con il termine cuing riflesso o esogeno perché l’attenzione è diretta le caratteristiche di basso livello degli stimoli esterni e non da un controllo volontario endogeno. nonostante i segnali luminosi non indicassero la posizione dei target le risposte furono più veloci quando questi venivano proiettati nelle vicinanze della posizione del lampo, anche se per un breve intervallo di tempo dopo il quale non vi era più distinzione. Quando tra il lampo di luce irrilevanti e la prestazione dei target trascorrevano più di 300 ms gli effetti sul tempo di reazione risultava invertito ovvero i partecipanti rispondevano più lentamente agli stimoli che comparivano nelle vicinanze del punto in cui si era verificato il lampo di luce. Questo effetto è conosciuto come “effetto inibitorio retroattivo o inibizione di ritorno”. Questi effetti possono essere spiegati con le teorie dell’adattamento ovvero se così non fosse le persone sarebbero continuamente distratti dalle cose che accadono intorno a loro e non sarebbero capaci di prestare attenzione a un fine. Il nostro sistema di orientamento riflesso dell’attenzione è costituito da meccanismi che impediscono che l’attenzione diretta in modo riflesso rimanga incollata su una porzione per più di un paio di centinaia di millisecondi. Per mantenere l’attenzione su un qualcosa di saliente possiamo rapidamente invocare i nostri meccanismi volontari che ci permetteranno di sostenere l’attenzione più allungo e di eliminare l’inibizione di ritorno. Sebbene i suggerimenti volontari e quelli involontari siano simili ovvero aumentano degli stimoli sensoriali attesi riducono quelli degli stimoli ignorati la differenza si trova nella velocità e nella durata dell’attenzione che veloce bassa nell’attenzione riflessa mentre l’effetto è più lento e sostenuto nel tempo in quella volontaria. La ricerca visiva: nella vita quotidiana l’attenzione involontaria quella riflessa interagiscono tra loro e competono per il controllo del focus dell’attenzione. Per comprendere come l’attenzione volontarie quella riflessa sono collegate alla ricerca visiva si potrebbe esaminare una serie di esperimenti effettuati, che andavano ad osservare come gli stimoli bersaglio vengano localizzati più velocemente quando sono identificabili con una singola caratteristica che differisce dalla caratteristica tipica dei distrattori. non importerà quanti distrattori siano presenti il tempo per ricercare tale Target sarà rapido questo effetto è conosciuto come “fenomeno pop-out”; se invece lo stimolo bersaglio presenta delle caratteristiche simili ai distrattori, il numero dei distrattori risulterà rilevante. il tempo impiegato per individualizzare il bersaglio sarà influenzato dal numero di distrattori presenti nell’insieme. Qualora dovessimo rappresentare su un grafico la funzione di ricerca essa sarà una retta orientata obliquamente. il tipo di ricerca che si metterà in atto e nota con il termine “ricerca di congiunzioni di caratteristiche” poiché il target bersaglio è costituito da un insieme di due o più caratteristiche (colore rosso e lettera O”. Teoria dell’integrazione delle caratteristiche dell’attenzione: l’teorizza che l’attenzione possa essere paragonata al fascio di luce che si muove in modo sequenziale da un item all’altro dell’insieme per ricercare il target obiettivo. Ricerche hanno dimostrato che i soggetti tendevano ad impiegare più tempo quando veniva chiesto loro di mettere in atto movimenti deliberati dell’attenzione in un compito di ricerca visiva in confronto a quando tali movimenti non erano richiesti e veniva consentito loro di esplorare in modo automatico. in altre parole è più facile trovare un target bersaglio permettendo all’attenzione di pagare in modo casuale piuttosto che indirizzandola in maniera ordinata. Questo modello suggerisce che la ricerca visiva possa essere simile al paradigma di cuing. Per questo motivo effettuare degli esperimenti in cui durante la ricerca visiva appariva uno stimolo sonda in una posizione sulla quale era focalizzata l’attenzione spaziale in questo modo lo stimolo sonda doveva indurre gli herpes visivi di maggiore grandezza 24 quando appariva nella parte del bersaglio. doveva invece scatenare stimoli inferiori quando appariva non nel bersaglio. Ai soggetti venivano presentati stimoli a forma di “the” di colore blu, verde e rosso in posizione sia verticale che rovesciata. in ciascuna prova il soggetto doveva cercare una tesi di uno specifico colore di cui viene era solo un esemplare ciò portava all’effetto pop-aut. Dopo un breve intervallo di tempo nella posizione del bersaglio o nella posizione di un altro dei distrattori nel lato opposto dell’insieme veniva presentato uno stimolo sonda dei contorni di un quadrato bianco che compariva attorno alla T di colore blu o a quella di colore verde ma mai attorno a quella russa. lo stimolo sonda dava risposte visive precoci più ampie. Cioè a P1 più grandi dove appariva nel Target designato in confronto alle regioni dove erano presentati solo i distrattori. Dunque l’attenzione spaziale influisce sull’elaborazione precoce della ricerca visiva.ma ancora non si è compreso come l’attenzione spaziale precede in modo gerarchico o meno l’attenzione per le caratteristiche o per gli oggetti. Attenzione per le caratteristiche: in un lavoro in cui misero a confronto l’attenzione spaziale con l’attenzione per le caratteristiche utilizzando il paradigma di cuiing volontario notarono che l’indicare tramite un suggerimento migliorava la prestazione e dunque che l’attenzione può essere diretta in anticipo sia sulle posizioni spaziali sia sulle caratteristiche non spaziali degli stimoli target. Mettendo a confronto gli arti si notò che l’attenzione per le caratteristiche e quella spaziale differiscono. Alcuni scienziati studiarono quali sistemi neurali erano coinvolti nella discriminazione delle caratteristiche in due diverse condizioni: la prima nell’attenzione divisa (i candidati dovevano prestare attenzione a colore, forma e movimento) nell’altra condizione studiarono l’attenzione selettiva (i candidati dovevano prestare attenzione a una sola caratteristica dello stimolo osservare se questo cambiasse forma) attraverso le immagini della PET dimostrò che vi era una maggiore attività nell’attenzione selettiva rispetto a quando veniva richiesta un’attenzione divisa. Attenzione selettiva: attiva regioni della corteccia extra striata distinte e ampiamente non sovrapposte e. Studi successivi con risonanza magnetica funzionale hanno identificato delle aree specializzate della corteccia visiva umana che le caratteristiche, come il movimento o il colore di uno stimolo. queste aree vengono modulate dall’attenzione visiva selettiva. Sebbene gli studi con risonanza magnetica funzionale abbiano localizzato le regioni sensibili dell’attenzione selettiva in base alle caratteristiche dell’oggetto come colore e movimento, si è riscontrato comunque che rispetto all’attenzione spaziale, l’attenzione selettiva avvenga a stadi successivi per quanto precoci. Come nella corteccia extra striata piuttosto che nella corteccia striata o i relais sotto corticali visive del talamo. 25 Per comprendere come l’attenzione spaziale e l’attenzione le caratteristiche sicuro ordinarono o si ostacolano sono stati fatti degli studi utilizzando gli Harp e la risonanza magnetica funzionale. è stato osservato attraverso N2pc degli ERP dove N2 È una seconda deflessione negativa, pc la posizione dell’elettrodo posteriore controllante lo stimolo atteso ovvero la porzione dove la componente compare., che questa seconda deflessione segnava l’attenzione e non implicita durante la ricerca visiva. Durante i compiti di ricerca un Erp per la selezione delle caratteristiche veniva generato nella corteccia occipitale ventrale, questo era seguito molto velocemente da una risposta N2pc proveniente dalle regioni più anteriori della corteccia occipito-temporale. questi risultati dimostrarono che l’attenzione per le caratteristiche può precedere quella spaziale quando la posizione del Target non è nata in anticipo. L’attenzione per gli oggetti: La proprietà dell’oggetto è l’insieme delle caratteristiche elementari dello stimolo che vengono combinate in un particolare modo così da identificare l’oggetto o una persona. Prestare attenzione a più oggetti comporta dei costi di tempo per questo motivo sono stati fatti degli studi utilizzando la risonanza magnetica funzionale per determinare quale effetto gli oggetti avessero sull’attenzione spaziale. Attraverso degli studi si scoprì che le aree corticali di V uno e Vv quattro aumentavano la loro attività in risposta a posizioni non presegnalate localizzati sullo stesso oggetto rispetto a posizioni non presegnalate che non si trovavano sullo stesso oggetto che era stato presegnalato. Questo portò a dimostrare che la rappresentazione degli oggetti può modulare l’attenzione spaziale. per capire se l’attenzione degli oggetti possa operare in maniera indipendente dall’attenzione spaziale è stato messo in pratica un altro studio basandosi sulle immagini della risonanza magnetica funzionale. In questo studio si sovrapposero le immagini del volto (che attivano l’area dei volti nel giro fusiforme, meno attiva in risposta a immagini di altri oggetti come ad esempio le case) e l’immagine di una casa (maggior attività nella corteccia paraippocampale che è più attiva in risposta immagini di case rispetto ai volti) inoltre veniva mosso , fatto oscillare uno dei due stimoli così da attivare anche l’area corticale del movimento MT/V5 si è riscontrato che le attività in queste tre aree aumentavano quando veniva chiesto ai soggetti di prestare attenzione a una delle due immagini piuttosto che l’altra e anche l’aria del movimento aumentava oltre che l’aria in relazione all’immagine che si muoveva. Dunque quando un compito non coinvolge l’attenzione spaziale il livello dell’analisi percettiva su cui agisce il controllo attentivi può essere quello delle rappresentazioni degli oggetti. Dunque l’attenzione visiva spaziale può influenzare l’elaborazione di uno stimolo in maniera molto precoce nelle vie sensoriali ascendenti e nella corteccia. inoltre l’attenzione aumenta l’elaborazione delle caratteristiche di uno stimolo. E ancora l’attenzione può essere diretta sugli oggetti e quindi può velocizzare l’elaborazione di tutte le caratteristiche . Le reti del controllo attentivi: il controllo dell’attenzione può essere sia finalizzato, sia diretto da uno stimolo. I modelli del controllo attentivi suggeriscono che per permettere il compimento delle azioni che ci permettono di prestare attenzione siano attivi due sistemi corticali fronto parietali separati: 1) Sistema dell’attenzione dorsale: interessato all’attenzione spaziale; 2) Sistema dell’attenzione e ventrale: interessato ad aspetti non spaziali dell’attenzione stessa. Il modello suggerisce che questi due sistemi cooperino così da produrre un comportamento normale.questa cooperazione però è compromessa nei pazienti affetti da negligenza unilaterale Rete dorsale: si attiva quando prestiamo attenzione finalizzata il circuito prende il nome di “ rete dell’attenzione frontoparietale dorsale” ed è composta da: 26 1) Campi oculari frontali, situati nella giunzione del solco precentrale e superiore di ciascun emisfero; 2) Campi oculari supplementari nella corteccia frontale; 3) Solco intraparietale; 4) Lobulo parietale superiore; 5) Precuneo, nel lobo parietale posteriore. 6) Regioni collegate. Durante questi studi si osservava una risposta prima della presenza del Tirget quando veniva chiesto ai soggetti di prestare attenzione ad una determinata parte dello schermo. Studi su pazienti con lesioni prefrontali hanno trovato che i pazienti che presentavano un danno alla corteccia frontale dovuta ad esempio un ictus mostravano una riduzione delle risposte visive evocate negli Erp registrati dalla corteccia visiva. Questo fa pensare che la corteccia frontale abbia un’influenza modulatori a sulla corteccia visiva. Altri studi hanno dimostrato che i neuroni dei campi oculari frontali sono coinvolti nel controllo attentivi o finalizzato specifico del compito. Dunque una delle componenti della rete di controllo dell’attenzione dorsale influisce sulla . Corteccia parietale: costituisce l’altra principale regione corticale che fa parte del sistema dell’attenzione fronto parietale. In quest’area possiamo trovare due regioni distinte: le aree dorsali lungo il solco intraparietale e il lobulo parietale superiore. queste parti fanno parte della rete dorsale mentre le arie ventrali che fanno parte della giunzione tempo parietale sono una parte della rete d’attenzione ventrale. Il lobo parietale a connessioni estese sottocorticali con il pulvinar e la corteccia frontale. Contiene molteplici rappresentazioni dello spazio. Gli spostamenti dell’attenzione sono collegati con variazioni dell’attività dei neuroni parietali; dunque quando l’attenzione viene diretta su uno stimolo il tasso di scarica dei neuroni parietali aumenta sia quando si utilizza lo stimolo come bersaglio o in un movimento di inseguimento o ancora quando si discriminano le sue caratteristiche in modo non esplicito. Molti studi si sono focalizzati sull’aria intraparietale specialmente sul solco intraparietale delle scimmie. Questa regione è coinvolta nei movimenti oculari e fa parte della rete dell’attenzione fronto parietale dorsale. LiP interessata alla posizione e alla salienza degli oggetti. 27 Rete dell’attenzione destra ventrale: È una rete che rimane a vigilare per un qualsiasi stimolo significativo in tutte le modalità sensoriali. Questa rete ventrale è fortemente lateralizzata nell’emisfero destro e include: 1) Giunzione tempo parietale, della corteccia parietale posteriore, localizzata nella giuntura del lobo temporale posteriore e il lobo parietale inferiore; 2) Corteccia frontale ventrale e, include anche la corteccia frontale ventrale costituita dei giri inferiore e mediale frontali. Gli studi hanno osservato che la giunzione tempo parietale della corteccia parietale posteriore non si attivava per compiti di attenzione visiva ma era impegnata sulla rilevanza del Target specialmente quando questo si verificava in una posizione inaspettata. Questo fa presupporre che questa parte sia impegnata da stimoli inaspettati o stimoli che variano in maniera inaspettata.si potrebbe considerare che la giunzione tempo parietale agisca come un interruttore che interrompe il focus attentivi stabilendo una rete dorsale finalizzata, infatti il risultato È un deficit dello sganciamento dell’attenzione spaziale. Le due reti sono interconnesse quella dorsale e quella ventrale e negli studi è stato evidenziato che la rete dorsale, più nello specifico il solco intraparietale fornisca alla giunzione tempo parietale della rete ventrale informazioni rilevanti dal punto di vista comportamentale degli stimoli cioè sulla loro salienza. Proprio poiché la rete ventrale non è interessata allo spazio non è stata trovata in tale rete nessuna mappa topografica. Componenti sottocorticale delle reti di controllo dell’attenzione: - Collicoli superiori: l’strutture mesencefaliche che ricevono input da molte fonti come: la retina, sistemi sensoriali, gangli della base e corteccia cerebrale. i collicoli superiori mandano anche molti output al talamo e il sistema motorio. Altri input dei campi oculari frontali aiutano a generare movimenti intenzionale, mentre altri input dei campi oculari parietali aiutano innescare saccadi riflesse. Questi neuroni non partecipano in maniera volontaria all’attenzione visiva di per sé ma fanno parte del sistema che controlla i movimenti oculari e sembrano avere un ruolo negli aspetti espliciti piuttosto che non espliciti dell’attenzione in quanto sono sensibili alla salienza di uno stimolo e guidano i movimenti oculari verso di essi. - Il pulvinar del talamo: riceve gli output derivanti dal collicolo superiore. È costituita da un gruppo di nuclei caratterizzati da connessioni con molte parti del cervello. possiede neuroni che rispondono a segnali visivi caratterizzati da una selettività per colore, movimento e orientamento; possiede aree che annoverano mappa retino topica del mondo visivo e interconnessioni con le aree corticali frontali, parietali, occipitali e temporali. Questa struttura implicata sia nell’attenzione volontaria che in quella riflessa in quanto i suoi neuroni si attivano quando lo stimolo è il bersaglio di un movimento oculare 28 o quando lo stimolo viene fatto oggetto di attenzione senza movimenti oculari bersaglio. Per comprendere come funziona ricercatori lo hanno disattivato nelle scimmie per capire in che modo l’attenzione variasse. Le scimmie trovavano difficoltà a orientare l’attenzione in modo non esplicito e a filtrare le informazioni distraenti. Avevano anche difficoltà con i distrattori in competizione e a discriminare il colore o la forma degli oggetti. Altri studi hanno notato che con l’aumentare del numero degli stimoli l’attività in quest’aria aumenta dunque quest’aria è importante della funzione spaziale non esplicita e per il filtraggio degli stimoli. Pazienti con lesioni mostrano difficoltà ad agganciare l’attenzione su una posizione dello spazio segnalata da un suggeritore.i loro tempi risultano molto dilatati sia per bersagli segnalati da un suggeritore valido sia per quelli segnalati da un suggeritore non valido. L’attenzione e quindi è controllata da tre reti interagenti tra loro: - Rete dell’attenzione dorsale: finalizzata ed è interessata al controllo dell’attenzione spaziale e alla salienza degli oggetti. Questa rete ci consente di mantenere l’attenzione su un fine contingente. - Sistema fronto parietale ventrale: interessato a disimpegnare e riorientare la nostra attenzione. È in grado di rilevare stimoli inaspettati, infrequenti o variabili. Il sistema rimane sempre in guardia pronto ad entrare in azione. Questi due sistemi interagiscono per permettere agli esseri umani di rimanere focalizzati su un fine rimanendo allo stesso tempo in allerta nei confronti di qualsiasi cosa potrebbe accadere. - Rete sottocorticale: contribuiscono all’aerosol, ai movimenti oculari, al filtraggio degli input e allo spostamento e orientamento dell’attenzione. Attività di barrage : capacità di direzionare l’attenzione sullo stimolo target in presenza di altri stimoli distrattori.gli stimoli possono essere di vario tipo (colori, figure di animali o di cose) la presentazione può avvenire per gruppi omogenei o eterogenei, ordinata o causale, spaziale o sequenziale. Un esercizio per l’attenzione selettiva è quella di individuare dei target in mezzo a dei distrattori mantenendo la concentrazione sullo scopo. Dual task: esercizio per l’attenzione divisa. Ad esempio affiancando parole e movimenti e con 29 Cap 9 la memoria L’apprendimento è il processo di acquisizione di nuove informazioni ed il risultato dell’apprendimento e la memoria. Quindi la capacità di acquisire, ritenere e richiamare esperienze ed informazioni.questa capacità è essenziale in quasi tutti i processi cognitivi.e frazionata in una serie di sistemi e sottosistemi tra loro interagenti; prestazioni circuiti neurali sotto stanti cambiano con lo sviluppo. I vari modelli distinguono i tipi di memoria che possono essere di breve vita come “la memoria sensoriale” che hanno una durata di millisecondi o secondi, memoria di breve e media vita come possono essere “memoria breve termine” e “memoria di lavoro” che durano da secondi a minuti. La memoria lungo termine può durare invece per decenni. I ricercatori dividono l’apprendimento e la memoria in tre principali stadi dell’operazione: 1) Codifica: si suddivide in acquisizione I sistemi sensoriali sono circondati da tantissimi stimoli, la maggior parte di questi stimoli produce solo una breve risposta sensoriale transitoria che svanisce molto rapidamente e non raggiunge mai la memoria breve termine. Tuttavia gli stimoli durante questo breve periodo sono disponibili per essere elaborati questa condizione conosciuta come buffer sensoriale. il secondo stadio è il consolidamento nel quale Vi sono dei cambiamenti nel cervello che stabilizzano una traccia nel tempo che risulti memoria lungo termine. 2) Immagazzinamento: È il risultato dell’acquisizione e del consolidamento e rappresenta la registrazione dell’informazione. 3) Recupero: permette l’accesso all’informazione immagazzinata usandola per creare una rappresentazione cosciente o eseguire un comportamento appreso come un atto motorio. Anatomia della memoria : il cervello alla capacità di modificarsi attraverso l’esperienza. Esistono diverse aree che sono coinvolte nell’apprendimento e che apprendono in diverso modo. Ad esempio l’amigdala è coinvolta nell’apprendimento della paura. Molte regioni sono coinvolte in uno o più aspetti della memoria. Sistema di memoria del lobo temporale mediale è formato da: - Ippocampo, un ripiegamento della corteccia temporale mediale che alla forma di un cavalluccio marino. - Corteccia entorinale - Corteccia peririnale - Corteccia paraippocanpale - Strutture Sab corticali che includono i corpi mammillari e i nuclei talamici anteriori. - Corteccia prefrontale 30 Le amnesie: È una forma di deterioramento della memoria che colpisce tutti i sensi. di solito si manifestano deficit specifici in specifici tipi di memoria. La perdita di memoria per eventi che si verificano dopo una lesione è conosciuto come amnesia anterograda ovvero l’incapacità di acquisire nuove informazioni. le amnesie che fanno riferimento a conoscenze che si sono verificati prima della lesione viene detta amnesia retrograda alcuni di questi amnesie retrograde è tempo realmente limitata cioè si può estendere a minuti a ore, giorni o all’intero arco della vita. L’essa tende a essere più violenta per i fatti recenti. Questo effetto è conosciuto come gradiente temporale o legge di Ribot. Prendendo in esame i casi di persone a cui erano stati praticati degli interventi chirurgici è stato possibile osservare come alcune parti della corteccia siano importanti per la memoria. Studi recenti hanno osservato che i lobi temporali mediali sono indispensabili per formare nuove tracce a lungo termine e per trasferire le informazioni sì fatti o persone dalla memoria a breve termine a quella lungo termine. Memoria a breve termine: 1) Memoria sensoriale: prende il nome di memoria ecoica quando fa riferimento all’udito e memoria iconica quando fa riferimento alla vista. per studiare la memoria ecoica si sono presi in esame i potenziali correlati a eventi ERP conosciuto come negatività da discordanza (MMN) elettrica, o Campo da discordanza (MMF) magnetica. Quando viene presentato l’input vengono generate delle onde. l’ampiezza di queste risposte certe e liberali a differenti intervalli di tempo tra i toni devianti e quelli standard può essere usata per indicare quanto allungo persiste la traccia della memoria econica. Che dura dai 9 ai 10 secondi. Studi invece sulla memoria iconica dimostrano che duri solamente dai 300 ai 500 ms. Questo tipo di memoria può ottenere molte informazioni ma solo per un periodo di tempo molto breve.. 2) Memoria breve termine: a un tempo più lungo che vada secondi a minuti è una capacità però più limitata. Gli studi partono da diversi modelli che prendono in considerazione i vari stadi di elaborazione dell’informazione durante l’apprendimento e la memoria. - Modello modale: propone che l’informazione è prima immagazzinata nella memoria sensoriale e poi spostata nel magazzino a breve termine grazie ad elementi selezionati dai processi attentivi. Una volta che l’elemento è nel magazzino della memoria breve termine se questo viene ripetuto può essere spostato nella memoria lungo termine. questo modello afferma anche che le informazioni possono essere perse a causa del decadimento (le informazioni si degrada e si perde nel tempo) , l’inferenza (una nuova informazione sostituisce la vecchia) o dalla combinazione di entrambi fattori. Questo modello richiede una elaborazione seriale ovvero essenziale la presenza della memoria breve termine per poter inserire le informazioni nella memoria lungo termine. alcuni casi di persone in cui la memoria breve termine era lesionata hanno però messo in evidenza che acquisire informazioni a lungo termine in assenza della memoria a breve termine era in qualche modo fattibile. Anche un altro paziente che presentava un tumore nel lobo temporale dopo l’operazione presenta problemi alla memoria a breve termine ma non riferiva a problemi in quella a lungo termine queste scoperte possono far pensare che l’informazione potrebbe entrare nella memoria lungo termine bypassando quella breve termine e provenire solo ed esclusivamente dai magazzini della memoria sensoriale. 31 Memoria di lavoro: rappresenta un magazzino che mantiene una capacità limitata di informazioni per un breve periodo da qualche secondo a qualche minuto. Consente anche di eseguire operazioni mentali i contenuti di questo magazzino come ad esempio avviene nelle somme aritmetiche. Le informazioni possono arrivare dalla memoria sensoriale oppure essere recuperate dalla memoria lungo termine. Si propose una suddivisione in tre parti: 1) Meccanismo esecutivo centrale: consiste in un centro di comando e controllo che coordina le interazioni tra due sistemi subordinati che sono in magazzino di memoria breve termine (circuito fonologico e taccuino viso spaziale) e la memoria a lungo termine. 2) Circuito fonologico: meccanismo che codifica l’informazione acustica nella memoria di lavoro. La sua esistenza bene intuita poiché nel lavoro di presentazione di consonanti gli errori erano basati su consonantico il suono simile piuttosto che su consonanti visibilmente simili. Altri studi notarono che le difficoltà del numero di input che potevano essere elaborati contemporaneamente aumentavano quando avevano un suono simile piuttosto che semanticamente simili. Il circuito fonologico si suddivide ulteriormente in magazzino acustico di breve vita (per gli imputo sonori) e una componente articolatoria (che partecipa alla ripetizione subito vocali dica degli item da ritenere sul breve termine). 3) Taccuino visuospaziale: È un magazzino che funziona in maniera parallela al circuito fonologico che però elabora input visivi. Deficit nella memoria di lavoro sono stati studiati soggetti che presentavano lesioni in aree specifiche del cervello. Ricerche hanno suggerito che il circuito fonologico e il taccuino viso spaziale potrebbero corrispondere alle funzioni della memoria di lavoro rispettivamente degli emisferi sinistro e destro. Questo si è ritrovato in pazienti con lesioni nel giro sopramarginale sinistro che presentavano un deficit nella memoria di lavoro fonologica. Il processo della ripetizione del circuito fonologico coinvolge una regione nell’aria premotoria sinistra (aria 44). Nel lavoro fonologico il network dell’emisfero sinistro coinvolge i lobi frontale laterale e parietale inferiore. Grafici di natura uditivo verbale non hanno dimostrato associazioni con deficit nella percezione o nella produzione del linguaggio. 32 Il taccuino viso spaziale viene compromesso si sono presenti dei danni nella regione parietooccipitale di entrambi gli emisferi; quelli più gravi però si riscontrano con danni all’emisfero destro. Questi soggetti presentano delle difficoltà in compiti non verbali che coinvolgono la memoria di lavoro come ad esempio tenere a mente i blocchi toccati da un’altra persona. Danni analoghi all’emisfero sinistro portano deficit nella memoria breve termine di materiali linguistici presentati visivamente. Attraverso studi con neuroimmagini si è dimostrata questa divisione nella memoria di lavoro. In uno studio ai soggetti venivano presentate una serie di lettere o in forma visiva o in forma fonologica. Dopo una pausa di tre secondi veniva chiesto loro di ricordare la posizione o le lettere durante la prova E se queste coincidevano con i nuovi input proposti. Per il compito verbale si trova un’attivazione nelle regioni dell’emisfero sinistro che corrispondeva la corteccia frontale laterale inferiore, per i compiti di memoria di lavoro spaziale le attivazioni Siri trovavano principalmente nella regione dell’emisfero destro frontale inferiore parietale posteriore, corteccia extra striata del lobo occipitale. In altri studi si trova all’attivazione della corteccia prefrontale ventro laterale sinistra quando si utilizzava la memoria di lavoro Visual spaziali. La tendenza ad una accensione bilaterale delle cortecce in compiti di memoria di lavoro spaziale può essere spiegata con il fatto che quando tentiamo di recuperare l’informazione possiamo utilizzare una Rick codifica verbale degli stimoli non verbali ad esempio “sopra sinistra”. Memorie a lungo termine: 1) Memoria dichiarativa: È la memoria per gli eventi e fatti personali e generali ai quali abbiamo un accesso cosciente e che sono riportati verbalmente. Questa memoria viene considerata come “memoria esplicita”. Essa dipende dal lobo temporale mediale. Viene poi suddivisa ulteriormente in memoria episodica e memoria semantica. La memoria episodica contiene tracce delle esperienze personali sulle nostre vite e cose con chi dove quando sono accadute. Costituisce quindi il nostro personale ricordo autobiografico e differisce dalla conoscenza personale. ( ad esempio la conoscenza del giorno in cui si è nati ma non vi è una ricordo). La memoria episodica è il risultato dell’apprendimento associativo rapido in cui il cosa, il dove, il quando e il perché di un singolo episodio e il suo contesto sono associati e legati assieme e possono essere revocati dalla memoria dopo un singolo episodio. La memoria semantica invece è la coscienza oggettiva di natura fattuale che però non include il contesto in cui è stata presa. Riflette la conoscenza di fatti e concetti come ad esempio il saper dire che ore sono e la capitale della Francia. Noi conosciamo che si è fatti ma non possiamo risalire a quando . La memoria episodica e quella semantica appaiono in età diverse ad esempio bambini di due anni sono stati in grado di dimostrare la rievocazione di cose alle quali avevano assistito a 13 mesi. Tuttavia finché non viene compiuto il 18º mese di età sembra che non riescano a includere loro stessi come parte del ricordo questa abilità sembra invece essere presente ed affidabile intorno ai tre-quattro anni. 2) Memoria non dichiarativa: viene detta anche memoria implicita poiché della sua esistenza non abbiamo un accesso cosciente. Questa memoria è importante quando esperienze precedenti facilitano la prestazione di un compito che non richiede intenzionalmente la rievocazione dell’esperienza essa coinvolge i gangli della base, il cervelletto, l’amigdala e la neocorteccia. Ad esempio fa riferimento all’apprendimento di un’abilità motoria o cognitiva. 33 - - - 3) Memoria procedurale: È un tipo di memoria non dichiarativa che dipende da vaste ripetute esperienze. Comprendono l’apprendimento di abilità motorie come andare in bicicletta o nuotare o l’apprendimento di abilità cognitive come saper leggere. Contesto di apprendimento procedurale è il compito dei tempi di reazione seriali. Che consiste nel premere con i polpastrelli dei tasti seguendo la sequenza delle luci date. Soggetti sani tendevano a rispondere più velocemente a sequenze ripetute rispetto a quelle casuali. Il grado di miglioramento permetteva di capire l’acquisizione della sequenza. Quando veniva chiesto loro qual era la sequenza sembravano non riconoscerla questo dà prova che esiste una memoria procedurale in assenza di una dichiarativa. Esperimenti su soggetti aventi amnesia anterograda dimostrarono che l’apprendimento procedurale è indipendente dalla memoria episodica. Le aree che vengono attivate durante l’apprendimento di abilità motorie sono: i gangli della base; pazienti con deficit ai gangli della base o alle afferenze a queste strutture sottocorticali mostrano prestazioni molto basse in una varietà di compiti con apprendimento procedurale. Questi deficienti si trovano in pazienti con il morbo di Parkinson in cui la morte delle cellule della Substanzia nigra interrompe le proiezioni dopaminergica e ai gangli della base. Tali deficit si riscontrano anche nei pazienti con la corea di Huntington che si caratterizza con una degenerazione dei neuroni nei gangli della base. 4) Priming: consiste in un cambiamento nella risposta uno stimolo o nella capacità di identificarlo. tale cambiamento è dovuto a una precedente esposizione a quello stimolo. Il priming può essere percettivo, concettuale o semantico. (Riconoscere il manubrio di una bicicletta in una determinata angolazione poiché se ne già fatto esperienza). Il priming percettivo si verifica all’interno del sistema di rappresentazione percettiva. la struttura e la forma degli oggetti e delle parole possono essere innescate dalle nostre precedenti esperienze e gli effetti persistono per mesi. Per valutarlo si usano dei compiti di completamento di frammenti di parole “ p_ens_eri” la prestazione sarà più veloce nelle parole già presentate questo è un tipico effetto di priming. Il priming funziona meglio quando la presentazione del target e la compilazione utilizzano gli stessi sensi. ad esempio se la parola viene presentata uditivamente il completamento del frammento della parola fatto visivamente l’effetto del priming sarà ridotto. Alcune prove confermano che alcune lesioni del cervello possono interessare il sistema di rappresentazione percettiva e mantenere intatta la memoria dichiarativa.si studiò un paziente che presentava una lesione al lobo occipitale destro, venne trattato con un intervento chirurgico in cui vennero rimosse parte delle aree di Brodmann 17,18 19 del lobo occipitale destro. Ciò provocò la cecità nel campo visivo sinistro. sebbene questo paziente dimostrava di avere una buona memoria esplicita le sue prestazioni non furono sufficienti nel test di priming percettivo implicito. Il priming concettuale dura di meno rispetto a quello percettivo. i soggetti saranno più veloci a rispondere a domande di cultura generale se il concetto è stato presentato in precedenza. Anche questo priming non è influenzato da lesioni al lobo temporale mediale ma da lesioni alle regioni temporali laterali e prefrontali. Il priming semantico: qui troviamo lo stimolo facilitante e il Target differenti ma collegati semanticamente.ad esempio lo stimolo facilitante potrebbe essere la parola martello ma la parola target è chiave inglese. questo tipo di priming è breve e dura solo pochi secondi. 34 Altri domini della memoria non dichiarativa sono: - il condizionamento classico: detto anche condizionamento Pavloviano Dove uno stimolo condizionato viene accoppiato a uno stimolo in condizionato che evoca una certa risposta da parte dell’organismo.dopo l’associazione lo stimolo condizionato è in grado di evocare una risposta condizionata simile a quella tipicamente provocata dallo stimolo incondizionato. Il condizionamento classico si divide in due forme: da una parte quello ritardato e dall’altra il condizionamento della traccia.nel condizionamento ritardato lo stimolo in condizionato viene presentato quando è ancora presente lo stimolo condizionato nel condizionamento della traccia c’è un gap temporale e dunque è necessario una traccia mnemonica. In alcuni studi è stato visto che è un danno all’ippocampo danneggia il condizionamento ritardato ma non quello della traccia. Ne deriva che alcuni tipi di apprendimento dipendono dall’ ippocampo e altri no. - L’apprendimento non associativo: non implica nessun tipo di associazione di due stimoli per evocare un cambiamento comportamentale. coinvolge forme di apprendimento semplici come l’abituazione ( la risposta di uno stimolo ripetuto nel tempo richiede meno tempo). Un altro tipo di apprendimento non associativo e la sensibilizzazione (la risposta aumenta in seguito a ripetute presentazioni dello stimolo). L’apprendimento non associativo attiva le vie sensoriali e sensomotoria riflesse. Sistema di memoria del lobo temporale mediale: Supporta la memoria a lungo termine (episodica e semantica) per poter comprendere si fa ricorso a casi di pazienti con deficit di memoria. Il lobo temporale mediale comprende: l’amigdala, l’ippocampo e le circostanti aree corticali paraippocampale, entorinale e peririnale. Inoltre sappiamo che i meccanismi di memoria si possono suddividere in: acquisizione, consolidamento, immagazzinamento e recupero. In un paziente che aveva perso la memoria in seguito a un episodio ischemico si riscontrava una grave amnesia anterograda è una lieve amnesia retrograda. dopo la sua morte l’autopsia rivelò che vi erano lesioni seppur limitate a una particolare regione dell’ippocampo ovvero alle cellule piramidali CA1. Questo caso conferma che l’ippocampo copre un ruolo importante nella formazione di nuovi e memorie. altre informazioni riguardante l’importanza dell’ippocampo nella memoria lungo termine provengono da pazienti con amnesia globale transitoria. In questi pazienti il flusso sanguigno del cervello è alterato soprattutto nella parte del lobo temporale mediale e al diencefalo. Questa situazione può portare ad un’ischemia transitoria che torna in seguito alla normalità. immagini ad alta risoluzione suggeriscono che le lesioni causate da questo evento siano localizzate nel sotto campo CA1 dell’ippocampo e che tali neuroni siano vulnerabili allo stress metabolico. questa disfunzione del flusso sanguigno causa un’improvvisa amnesia transitoria anterograda e l’amnesia retrograda si estende settimane, mesi e a volte anche anni. Entro 24-48 ore torna alla normalità anche se persistono lievi deficit per qualche giorno o settimana. L’importanza dell’ippocampo nella formazione della memoria lungo termine può essere riscontrata anche da pazienti con amnesia causata da lesioni nelle regioni connesse con il lobo temporale mediale ma al di fuori da esso come ad esempio un danno al diencefalo. Un ulteriore evidenza si riscontra nei pazienti malati di Alzheimer in cui vi è un deterioramento neurale che provoca danni gravi alle strutture del lobo parietale della corteccia retrospleniale, del cigolato posteriore, del precuneo e del giro angolare. Ma scienziati credono che anche l’ippocampo si deteriori prima rispetto a persone sane. le placche amiloide e gli ammassi neurofribrillari sì concentrano nell’area temporale mediale.misurazione del volume del cervello con la risonanza magnetica hanno mostrato che le dimensioni dell’ippocampo cambiano con la progressione dell’Alzheimer e che persone con un ippocampo più spesso tendono a sviluppare la demenza in misura minore. Nei pazienti con Alzheimer si verifica anche una perdita importante di cellule 35 acetilcolinergiche che si connettono all’ippocampo e la corteccia prefrontale. Questo fattore sembra giocare un ruolo importante nella perdita progressiva della capacità di formare nuove memorie episodiche. Per comprendere l’importanza dell’amigdala sulla memoria, sono state prodotte chirurgicamente lesioni nella boh temporale e nell’amigdala di scimmie. Dopo aver prodotto delle lesioni chirurgiche o all’ippocampo o all’amigdala o entrambe le strutture le scimmie venivano sottoposte al “compito di discriminazione ritardata di un oggetto non corrispondente a un campione”. La scimmia viene chiusa all’interno di una gabbia in cui vi è uno sportello che non permette di vedere al di fuori.a sportello chiuso viene posta una ricompensa sotto a un segnale la scimmia una volta che lo sportello verrà aperto comprenderà che la ricompensa del cibo si trova sotto al segnale indicato.a questo punto lo sportello viene Ri abbassato e viene dato un nuovo segnale non corrispondente al vecchio che andrà a coprire la ciotola con il cibo. Dopo un periodo di apprendimento la scimmia riuscirà a discriminare il nuovo segnale. Il deficit di memoria risulta presente soltanto se la lesione include l’ippocampo e la amigdala. questo portò a pensare che l’amigdala sia una struttura chiave nel sistema di memoria. Tali dati però non concordavano con i dati provenienti da soggetti che presentavano amnesia anterograda causata da lesioni soltanto ai neuroni CA1 dell’ippocampo e non dell’amigdala. Per questo motivo altri studi produssero nel cervello delle scimmie lesioni più selettive che interessavano l’amigdala o l’ippocampo o la corteccia che circonda ciascuna di queste strutture. le lesioni prodotte coinvolgevano l’amigdala, la corteccia entorinale ( area 28-34 di brodmann) e la neocorteccia del giro paraippocampale ( 3536 area di brodmann) si notò che quando le lesioni riguardavano solamente l’ippocampo o l’amigdala senza toccare le aree della corteccia circostante una lesione o l’assenza di una lesione dell’amigdala non influenzava la memoria delle scimmie.dunque l’amigdala non poteva far parte del sistema a supporto dell’acquisizione della memoria lungo termine. Altri studi hanno generato lesioni selettive alla corteccia circostante nelle regioni perirenale, interinale e paraippocampale. le aree paraippocampali e perirenale ricevono informazioni dalla corteccia associativa visiva, uditiva e somatosensoriale e trasmettono questi input all’ippocampo. Lesioni in queste aree provocano un peggioramento della prestazione di memoria nei test di discriminazione ritardata di un oggetto non corrispondente a un campione. Dunque lesioni dirette all’ippocampo o lesioni che danneggiano le connessioni input e output tra ippocampo e nella corteccia provocano deficit di memoria gravi. l’amigdala non è una componente cruciale del sistema memoria episodica ma lo è per la memoria emozionale. Inoltre gli studi hanno dimostrato che il lobo temporale mediale non è essenziale per i processi a breve termine o per la memoria di lavoro. il lobo temporale mediale non è la sede del magazzino della memoria lungo termine poiché si è danneggiato non porta a un’amnesia retrograda totale ma rimane comunque una componente cruciale nell’organizzare e consolidare la memoria lungo termine che sono immagazzinate in modo permanente in forma distribuita in tutta la neocorteccia. 36 Studi sui roditori: i primi studi effettuati sui roditori hanno riscontrato che le lesioni ippocampali non producevano un’alterazione dell’apprendimento stimolo-risposta. per questo proposero che l’ippocampo fosse coinvolto nell’immagazzinamento e nel recupero della memoria contestuale. questo è dato anche dal fatto che quando venivano inseriti degli elettrodi nell’ippocampo del ratto si attivavano delle cellule conosciute come “cellule per i luoghi”; queste cellule si attivavano solo quando il ratto si trovava in una posizione precisa e di fronte a una particolare direzione.si inibivano quando l’animale si spostava in un ambiente differente per poi scaricare in maniera specifica per una posizione in quella nuova area. Dunque come l’animale si sposta nell’ambiente l’attività di specifici neuroni ippocampali di CA1 e CA3 correla con specifiche posizioni. questi risultati portarono a pensare che l’ippocampo rappresenti contesti spaziali in altre parole il dove della memoria contestuale. L’ippocampo è coinvolto nell’apprendimento della navigazione spaziale. La navigazione spaziale nei ratti e testato utilizzando il labirinto acquatico di Morris. è costituito da una vasca circolare con acqua scura. sopra l’acqua sono presenti diversi segnali visivi identificabili e da qualche parte sotto la superficie dell’acqua c’è una piattaforma invisibile. i ratti vengono immersi nella vasca da differenti punti in differenti prove. il tempo da essi impiegato per raggiungere la piattaforma si riduce nel corso del tempo, indicando così di aver appreso dove fosse la piattaforma in relazione ai riferimenti visivi al di sopra dell’acqua. I ratti con lesioni all’ippocampo non riescono ad imparare ad associare i riferimenti visivi alla posizione della piattaforma.i ratti tendono a nuotare a caso in ogni prova alla ricerca della piattaforma.tuttavia se vengono immersi nello stesso identico. Tendono ad apprendere dove è localizzata la piattaforma sottostante.questo dimostra che sebbene siano presenti lesioni all’ippocampo sono in grado di apprendere un compito ripetuto di cui hanno fatto pratica ma non sono in grado di mettere in relazione le informazioni sullo spazio con le differenti informazioni contestuali. Altri studi hanno osservato che alcuni neuroni dell’ippocampo del ratto scaricano in risposta a specifici odori o a specifiche combinazioni di odori e posizioni, ad alcuni stimoli visivi e uditivi o alla combinazione di entrambi, e ad altre caratteristiche non spaziali incluso il comportamento questo ha portato a pensare che l’ippocampo possa essere cruciale per mettere insieme le diverse informazioni contestuali per formare una complessa forma di memoria di esse. In recenti studi è stato osservato anche che l’ippocampo danneggiato provochi dei deficit di navigazione spaziale sia sulle memorie recenti sia su quelle remote. 37 Cv la rievocazioni di memoria contestuali nei ratti e studiata usando l’apprendimento contestuale alla paura.consiste nel posizionare in una piccola camera con caratteristiche visive specifiche i ratti e a sottoporli ad una scossa ai piedi.i ratti successivamente mostrano una risposta di paura che viene percepita attraverso il loro comportamento di immobilizzazione che avviene quando vengono riposizionate nella stessa camera.ad alcuni ratti poi venivano ricavati dei danni all’ippocampo parziali o totali.si osservato che dopo una distruzione dell’ippocampo i ratti non si immobilizzavano mentre i ratti con un danno parziale mostravano qualche forma di immobilizzazione ma in misura minore rispetto ai reati di controllo. La gravità dell’amnesia retrograda era legata all’estensione del danno dell’ippocampo l’amnesia era presente anche per la memoria contestuale e retrograda remota.questi studi dimostrano che l’ippocampo svolge un ruolo sia nella rievocazione della memoria lungo termine contestuale “ed episodica”. Si è notato anche che col tempo i topi che riuscivano a discriminare camere simili tendevano dopo lasso di tempo a non distinguere più generalizzando la reazione di paura dunque si può affermare che le memorie contestuali diventano meno dettagliate e molto generiche con Los trascorrere del tempo così che la reazione di paura si attiva in contesti nuovi ma simili. Imaging nella memoria umana: Studi con risonanza magnetica funzionale hanno dimostrato che l’ippocampo nell’uomo è attivo quando vengono codificate nuove informazioni. Queste immagini vengono raccolte mentre viene chiesto ai partecipanti di ricordare degli item “paradigma del ricordo seguente”. L’attività celebrale viene misurata sia con la risonanza magnetica funzionale che con gli ERP sia quando immagazzinano le informazioni sia quando le riportano in memoria. Da queste immagini si è rilevato che vi era un’attività sia nelle regioni dell’ippocampo sia nella corteccia paraippocampale posteriore durante la codifica. Dunque così come negli studi degli animali anche in quelli degli umani l’ippocampo è importante per la formazione di nuovi ricordi a lungo termine (codifica) È stato osservato anche che durante la codifica si attivavano anche regioni della corteccia frontale. Altri studi hanno scoperto che l’ippocampo è coinvolto anche nel recupero delle informazioni della memoria lungo termine. attraverso immagini con risonanza magnetica funzionale è stato visto che le arie dell’ippocampo si accendevano quando i partecipanti memorizzava no una lista di parole e quando 20 minuti più tardi dovevano recuperarli. Fu interessante osservare che l’ippocampo si attivava solo per gli item che venivano veramente correttamente ricordati indicando così una memoria episodica mentre non si riscontrava un’accensione per la memoria basata sulla familiarità, ovvero quando l’item sembrava ricordare qualcosa ma in realtà non era stato presentato in precedenza. 38 Le diverse aree del lobo temporale mediale della memoria lungo termine: alcuni studi si sono soffermati su quali parti del lobo temporale mediale fossero coinvolti nella memoria episodica e invece in quella basata sulla familiarità. Nella memoria episodica l’evento viene codificato insieme al momento specifico e ad uno spazio specifico. Diversi tipi di informazioni provenienti da tutta la corteccia convergono nelle regioni del lobo temporale mediale che circondano l’ippocampo, non tutti però effettuano la stessa strada. Sappiamo che le informazioni sulle caratteristiche degli item “cosa” che provengono dalle regioni sensoriali unimodali della neocorteccia decorrono attraverso le porzioni anteriori della regione paraippocampale conosciuta anche come corteccia peririnale. Le informazioni sugli item relative al “dove” provenienti dalle aree della neocorteccia Polimodale attraversano le porzioni più arretrate della corteccia paraippocampale. Entrambe le informazioni proiettano entro la corteccia entorinale ma non convergono fino a quando non si trovano all’interno dell’ippocampo. Un modello “ modello della dimensione degli item con i loro contesti” afferma che la corteccia perirenale rappresenta le informazioni relative agli item specifici sul chi e che cosa mentre la corteccia paraippocampale rappresenti informazioni degli item relativi al contesto in cui sono stati incontrati “il dove il quando” l’ippocampo alla funzione di unire le rappresentazioni degli item con quelle dei loro contesti. Da questa Unione si avrà quella che viene definita memoria relazionale. dunque per riconoscere un item familiare è sufficiente unicamente la corteccia perirenale ma per riconoscere l’intero episodio e qualsiasi cosa esso collegato è necessario l’ippocampo. Dunque durante il riconoscimento fondato sulla familiarità erano attive le regioni del giro paraippocampale mediale anteriore nell’ambito della corteccia perirenale e attorno adesso.mentre quando il ricordo si basava su informazioni episodiche si attivano l’ippocampo e la corteccia paraippocampale posteriore. 39 La memoria relazionale viene definita così in quanto recupera informazioni episodiche che conducono ad esperienze che siamo in grado di indicare di aver incontrato recuperando le fonti e comprendendo che è un’esperienza diversa da quella incontrata precedentemente. Per studiare questo tipo di memoria vengono utilizzati metodi che utilizzano la fissazione oculare. Ai partecipanti viene chiesto di guardare delle scene complesse che vengono manipolate così da modificare le relazioni spaziali degli oggetti. I partecipanti sani dimostrano di essere sensibili a tali manipolazioni mentre coloro che soffrono di amnesia temporale mediale non sono altrettanto sensibili. questo dimostra che la memoria relazionale è distinta dalla memoria episodica che richiede consapevolezza cosciente. Le prestazioni nei money che sono state valutate in tre diversi tipi di compiti relazionali: spaziale, associativo sequenziale. Inoltre sono stati messi a confronto soggetti sani e singoli ricordi di soggetti con amnesia. I soggetti con un solo danno all’ippocampo mostravano una compromissione in tutte e tre i tipi di compiti relazionali ma non nel compito del ricordo di singoli item. Pazienti che invece presentavano un danno più esteso al lobo temporale mediale presentavano una compromissione in entrambi tipi di compiti. Questo perché quando si valuta la relazione tra i diversi item si nota un’attivazione dell’ippocampo mentre quando si osserva e si codifica solo item non si riscontra attivazione dell’ippocampo ma in altre regioni del lobo temporale mediale specialmente nella corteccia perirenale. Recupero e riattivazione della memoria lungo termine: da una parte vi sono le proiezioni che partono dalla neocorteccia e arrivano all’ippocampo che danno informazioni sulla cosa e sul dove degli item; dall’altra parte abbiamo proiezioni che partono dall’ippocampo e che viaggiano a ritroso prima verso la corteccia entorinale poi verso la corteccia pererinale e paraippocampale per arrivare infine alle aree neocorticali che hanno fornito l’input primario verso la neocorteccia. Durante studi sul recupero della memoria ai partecipanti vennero somministrati degli item sia in versione uditiva che visiva.giorni dopo utilizzando la risonanza magnetica funzionale venivano presentati degli item e chiesto loro se erano stimoli uditivi o visivi.le immagini dimostrarono che le regioni della neocorteccia che si erano attivate durante la percezione si riattivavano in maniera simile anche durante il recupero. durante il recupero della memoria le regioni di attività delle specifiche modalità della neocorteccia risultavano sotto insiemi delle aree attivate dalla presentazione delle sue informazioni percettive quando non veniva richiesto alcun compito di memoria. Altri studi hanno dimostrato che regioni dell’elaborazione visiva nella corteccia infero temporale erano coinvolte nella rappresentazione del recupero dell’informazione visiva mentre l’attività parietale più dorsale e quella occipitale superiore erano collegate al processo della ricerca mnemonica per gli item rilevanti. Corteccia frontale: Studi su pazienti con amnesia hanno affermato che la corteccia frontale è coinvolta nei processi di memoria sia lungo termine sia a breve termine. Si è riscontrato che la corteccia frontale sinistra e spesso coinvolta nella codifica episodica delle informazioni mentre la corteccia frontale destra e spesso coinvolta durante l’attività di recupero dell’episodio. Si è osservato anche che la codifica semantica e il suo recupero coinvolgono la corteccia frontale sinistra compresa l’area di Broca (area 44 fino alla 46) e la regione laterale ventrale (45 e 44). Altri studi hanno ipotizzato che la lateralizzazione dell’attività della corteccia frontale durante recupero a lungo termine è più legata alla natura del materiale che si deve recuperare piuttosto che a una distinzione tra codifica e recupero. L’emisfero sinistro sembra infatti più coinvolto nei processi di codifica linguistica mentre la corteccia frontale destra sembrerebbe più coinvolta nella memoria per informazioni spaziali collegate all’oggetto. Corteccia parietale: Si è osservato che lesioni retrospleniali possono portare ad un’amnesia retrograda e anterograda. questi risultati prendono il nome di (effetto vecchio-nuovo) E dimostrano che quando un elemento viene riconosciuto correttamente come un elemento precedentemente incontrato vi sono delle reazioni nella corteccia parietale posteriore, inclusi i lobuli parietali inferiori e superiori e le 40 strutture mediali che si estendono dal precuneo alla corteccia cingolata posteriore E alla corteccia retrospleniale. rispetto a quando l’individuo riconosce in maniera corretta che un elemento non era mai stato incontrato. Nuovi studi hanno evidenziato che il recupero della memoria in particolare per informazioni contestuali e associato all’attività della corteccia parietale posteriore e del la corteccia retrospleniale. Durante la codifica queste arie sono meno attive rispetto ai livelli di base a meno che gli item non siano codificati in modo autoreferenziale o evochino informazioni autoreferenziali o ancora vi sia un’elaborazione di tipo emotiva. Questi risultati portano a pensare che la corteccia retrospleniale sia più in sintonia con fonti informative interne. queste regioni parietali sono attive anche durante lo stato conscio di riposo quando si attiva una rete di aree corticali associative altamente interconnesse detta anche rete della modalità di default. questa rete di default è attiva quando il soggetto pensa a scenari passati o futuri relativi al sé. Anche la connessione anatomica della corteccia parietale suggerisce il suo coinvolgimento nella memoria poiché le cortecce parietali laterali, retro spinali e ci angolate posteriori sono connesse al lobo temporale mediale sia direttamente sia indirettamente.in particolare la corteccia retro spegnare e estesamente interconnessa con la corteccia paraippocampale ed entrambe si interfacciano con regioni sìmili nell’ippocampo posteriore nel subiculum, nei corpi mammillari e nel talamo anteriore e nella rete di default. La corteccia perirenale mostra invece un partner di connettività diverso non con l’ippocampo posteriore ma con l’ippocampo anteriore, l’amigdala, la corteccia temporopolare ventrale E la corteccia orbito frontale laterale. Considerando questa anatomia è stato proposto un modello della memoria composta da due sistemi: - Il sistema temporale anteriore, che include la corteccia perirenale e le sue connessioni; - Il sistema mediale posteriore è composto dai corpi mammillari, dei nuclei talamici anteriori, dal subiculum e dalla rete della modalità di default. Questi sistemi sono coinvolti oltre che nella memoria anche in altri aspetti della cognizione.la corteccia perirenale del sistema anteriore sostiene la memoria per gli item ed è coinvolta nel riconoscimento basato sulla familiarità, nell’associazione delle caratteristiche degli oggetti e nell’esecuzione di discriminazioni percettive e semantiche di fine risoluzione. La corteccia paraippocampale e la corteccia retrospleniale sono coinvolte nella memoria basata sul ricordo, le memorie per le scene, per le disposizioni spaziali e per i contesti. Alcuni ricercatori sostengono che queste arie assieme ad altre strutture del sistema mediale posteriore possano costituire rappresentazioni delle relazioni tra l’entità. La corteccia parietale ben nota per il ruolo che ha nell’attenzione è implicata anche nella memoria. L’ipotesi è che sostenga il mantenimento nella memoria di lavoro. 41 Il consolidamento della memoria: il consolidamento è il processo che stabilizza una memoria nel tempo dopo essere stata acquisita.vi è un processo di consolidamento inizialmente rapido che è seguito da uno permanente e lento. Durante studi che utilizzano una terapia elettroconvulsivante si osservato che le informazioni che tendevano ad essere perse erano quelle che avevano subito un consolidamento rapido iniziale ma che non erano arrivate a un consolidamento permanente più lento.i lobi temporali mediali soprattutto l’ippocampo sono importanti per il consolidamento rapido e l’immagazzinamento iniziale delle informazioni per la memoria episodica e semantica. Le aree del consolidamento lungo sono ancora incerte la teoria del consolidamento standard propone un’importanza cruciale della neocorteccia mentre l’ippocampo giocherebbe solo un ruolo temporaneo. Poi passano alla neocorteccia dove si arriva a una memoria permanente. il consolidamento avviene dopo ripetute riattivazione della memoria che creano connessioni dirette all’interno della corteccia tra le varie rappresentazioni. questo modello propone lo stesso schema sia per la memoria episodica e semantica. Un altro modello detto teoria multi traccia suggerisce che immagazzini a lungo termine per informazione semantica dipendano dalla neocorteccia mentre alcuni aspetti della memoria episodica continuino a dipendere dall’ippocampo anche durante il consolidamento. Traccia viene rievocata maggiore saranno il numero di tracce delineate. Le tracce non sono simili ma possono differire per gli attributi.questa teoria afferma anche che le tracce episodiche tendono a degradarsi nel tempo e siano lentamente conservate nella memoria semantica. Lobo temporale anteriore laterale: la neocorteccia temporale al di fuori della regione mediale importante per il consolidamento permanente dell’informazione semantica.qualora si presentassero delle lesioni si potrebbe arrivare a un AN amnesia retrograda grave che potrebbe coinvolgere addirittura l’intera vita del paziente. (Corteccia laterale del lobo temporale anteriore, vicino al polo anteriore, aree associate alla demenza semantica) e.è stato osservato però che in alcune condizioni alcuni pazienti possono formare nuovi ricordi episodici a lungo termine. Questo è possibile spiegarlo se le regioni laterali e anteriori del lobo temporale rappresentino la sede dove le memorie semantiche a lungo termine sono immagazzinate altrimenti un’altra alternativa è che queste regioni possono essere importanti per il recupero delle informazioni dei magazzini a lungo termine.questa ultima teoria si riscontra anche in studi di visualizzazione neuro funzionali che suggeriscono che le tracce vengono immagazzinate sottoforma di rappresentazioni distribuite in tutta la neocorteccia compresa e le regioni della codifica iniziale dell’informazione percettiva e le regioni della rappresentazione delle informazioni associate ai dati in ingresso. il lobo temporale mediale può coordinare il consolidamento nel tempo di queste informazioni. 42 Basi cellulari dell’apprendimento e della memoria: L’apprendimento Hebbiano Afferma che sei una sinapsi è attiva mentre attivo neurone post sinaptico, quella sinapsi risulterà rafforzata. Principali componenti neurali dell’ippocampo: le aree neocorticali associative proiettano alla corteccia entorinale attraverso la corteccia paraippocampale o la corteccia perirenale.la corteccia antorinale proietta attraverso la via perforante sulle cellule granulari del giro dentato, con input ecitatori. Assoni non mielinizzati conosciuti come fibre muscolo ID collegano le cellule granulari al giro dentato alle spine di Riddick delle cellule piramidali CA3. I collaterali di shaffer collegano tra loro le cellule piramidali CA1 e CA3. Questo sistema viene utilizzato per studiare la plasticità sinaptica come meccanismo dell’apprendimento a livello cellulare. Si scoprì che il potenziamento a lungo termine si otteneva stimolando gli assoni della via perforante che portarono ad un aumento a lungo termine dell’ampiezza dei potenziali post sinaptici eccitatori. Questo effetto si evidenzia anche nelle altri due proiezioni eccitatoria dell’ippocampo. In animali vivi il potenziamento a lungo termine può durare giorni o settimane.è stato trovato che il potenziamento a lungo termine differisce dalle tre vie e si può trovare anche in altre regioni cerebrali inclusa l’amigdala e i gangli della base, il cervelletto e la corteccia tutte le aree coinvolte nell’apprendimento. Per studiarlo viene inviato un singolo impulso e si misura l’EPsP che ne deriva. L’ampiezza di questa prima registrazione rappresenta la forza della connessione prima dell’introduzione del potenziamento a lungo termine. Dopo che potenziamento a lungo termine è stato indotto si invia un nuovo impulso e si misura la grandezza del’EPSP nella cellula post sinaptica si aumenta il segno di una forza dell’effetto sinaptico. Si afferma anche che se un input debole è uno forte agiscono contemporaneamente su una cellula entrambe le vie mostrano un fenomeno di potenziamento a lungo termine e dunque la sinapsi debole diventa più forte. Dei risultati se questi studi si possono enunciare tre regole per il potenziamento a lungo termine associativo: 1) Cooperatività: più input devono essere attivi nello stesso momento; 2) Associatività: input più deboli risultano rafforzati quando si verificano insieme con input più forti; 3) Specificità soltanto la sinapsi stimolata esibisce il potenziamento Il potenziamento a lungo termine avviene solo quando le cellule post sinaptiche ricevono gli impulsi eccitatori e sono de polarizzata. il potenziamento a lungo termine risulta infatti ridotto dagli input che inibiscono le cellule postina psiche, questo è quello che succede quando si verifica l’abituazione. il potenziamento post sinaptico È impedito anche quando le cellule post sinaptiche sono iper polarizzata. Il recettore NMDA: il potenziamento a lungo termine dipende dal neurotrasmettitore glutammato. Il glutammato si lega due tipi di recettori glutammatergici. Il recettore NMDA sono localizzate sulle spine dendritiche dei neuroni post sinaptici e sono centrali per produrre il potenziamento a lungo termine ma non per mantenerlo.. 43 Il mantenimento è dato dai recettori AMPA. I recettori NMDA possono essere bloccati anche dagli ioni magnesio e possono essere espulsi solo quando la cellula è de polarizzata.i canali ionici si aprono solo se si verificano due condizioni: 1) Neurotrasmettitore glutammato si lega ai recettori; 2) La membrana è depolarizzata Dunque questi recettori sono trasmettitori e voltaggio dipendenti. l’apertura del canale ionico consente agli ioni CA di entrare nella cellula post sinaptica. Il CA funge da messaggero intracellulare del segnale che modifica le attività enzimatiche dalle quali dipende il cambiamento della forza sinaptica. Studi sui topi ai quali era stato bloccato l’apprendimento a lungo termine hanno portato a constatare che vi erano inibizioni dei processi normali della memoria spaziale.questo ha portato ad affermare che il blocco dei recettori NMDA E l’inibizione del potenziamento a lungo termine danneggino la memoria spaziale. Questi recettori nella regione CA1 dell’ippocampo sono necessari per molte forme di plasticità sinaptica e si attivano per l’apprendimento spaziale del contesto. Una volta che si è verificato l’apprendimento possono essere formate nuove tracce di memoria senza la loro attivazione. 44 Cap 11 Linguaggio Anomia: non riuscire a trovare le parole per etichettare le cose. I soggetti sono in grado di comprendere le parole ma vi è una difficoltà nella produzione. Questo dimostra che i processi di comprensione e produzione del linguaggio sono diversi. Gran parte dell’elaborazione linguistica e lateralizzata nelle regioni emisferiche sinistre che circondano la scissura di Silvio. Le aree linguistiche includono la corteccia temporale sinistra (in cui appartiene anche l’area di Wernicke) porzioni della corteccia temporale anteriore sinistra, il lobulo parietale inferiore (di cui fanno parte il giro sovramarginale e il giro angolare), la corteccia frontale inferiore sinistra (che include l’area di brocca), la corteccia insulare sinistra. Queste regioni creano “la rete perisilviana sinistra per il linguaggio” poiché circondano la scissura di Silvio. Sebbene l’emisfero sinistro sia l’attore principale per il linguaggio l’emisfero destro dà il suo contributo con il solco temporale superiore destro che si attiva nell’elaborazione del ritmo del linguaggio (prosodia), la corteccia prefrontale, il giro temporale medio e il cingolato posteriore di destra si attivano quando le frasi hanno un significato metaforico. Il linguaggio coinvolge anche tutte quelle strutture corticali e sottocorticali coinvolte nella tempo realizzazione motoria e nel movimento (corteccia premotoria, corteccia motoria e aria supplementare motoria, talamo, gangli della base e cervelletto). Afasia: termine generico che si riferisce a un deficit della comprensione e produzione del linguaggio, può essere accompagnata da problemi linguistici causati dalla perdita del controllo dei muscoli articolatori (disartria) o da deficit nella pianificazione motoria delle articolazioni (aprassia per il linguaggio). È molto comune in seguito a un danno cerebrale. 45 Afasia di Broca: è conosciuta anche come afasia anteriore ed è la forma di afasia più antica e meglio studiata. Broca nel XIX secolo effettuare un’autopsia su un paziente che per diversi anni aveva potuto esprimere solo la parola “tan”. Dall’autopsia emerse che il paziente aveva una lesione nella porzione posteriore del giro frontale inferiore sinistro, questa lesione è conosciuta anche come parte triangolare e parte opercolare oggi viene definita come area di Broca. Dopo diversi studi Broca affermò che le aree celebrale deputate alla produzione del linguaggio erano localizzate nel lobo frontale inferiore dell’emisfero sinistro. Nei pazienti affetti da afasia di Broca e l’acqua è spesso telegrafico, faticoso e prodotto in modo irregolare. E a fasici di Broca sono consapevoli dei loro errori e hanno un basso livello di tolleranza alla frustrazione. questi pazienti possono anche manifestare deficit di comprensione relativi alla sintassi E spesso possono comprendere solo le forme grammaticali più Elementali e iper apprese questo deficit è noto come afasia con agrammatismo. Tuttavia brocca non sezione ormai il cervello del suo paziente. Studi recenti utilizzando le immagini provenienti da risonanza magnetica dimostrano che i danni sono più profondi e includono la corteccia insulare e porzioni dei gangli della base. 46 Afasia di Wernicke: E nota anche come afasia posteriore o ricettiva. Riguarda la comprensione del linguaggio. L’il pazienti affetti da questa sindrome hanno problemi a comprendere il linguaggio parlato o scritto, la loro rosolia e grammatica sono normali ma quello che dicono risulta privo di senso. Questi pazienti presentavano lesioni nelle regioni posteriori del giro temporale superiore nota come area di Wernicke.poiché l’elaborazione uditiva avviene una regione vicina alla corteccia temporale superiore vernice ne dedusse che questa regione più posteriore partecipava al magazzino uditivo delle parole. Oggi grazie lo studio di immagini abbiamo prove che la fasina di Wernicke grave e persistente soltanto se la lesione riguarda l’area di Wernicke e la corteccia circostante del lobo temporale posteriore, o la sottostante sostanza bianca che collega le aree del linguaggio nel lobo temporale alle altre regioni cerebrali. Lesioni limitate alla sola area di Wernicke portano ad un afasia temporaneo. Afasia di conduzione. Nel 1880 si introdusse l’idea di una terza regione che conservava le informazioni concettuali sulle parole. Si pensava che una volta che la parola fosse stata recuperata dal magazzino tale informazione veniva inviata all’area dei concetti così da arricchirla con tutto ciò che fosse a essa associato.venne così descritto il modello localizzazionista classico. Modello localizzazionista classico: informazione linguistica, il magazzino delle parole (a uguale area di Wernicke), la programmazione del linguaggio (M uguale area di brocca) e il magazzino delle informazioni concettuali (B) sono posizionate in regioni cerebrali interconnesse da tratti di materia bianca. Il tratto di materia bianca che fluisce dall’area di Wernicke a quella di Broca È il fascicolo arcuato. Qualora questo fascicolo venisse danneggiato si presenta una sindrome conosciuta come afasia di conduzione. questi pazienti hanno problemi nella produzione verbale spontanea e nella ripetizione del discorso. Delle volte usano anche le parole in modo scorretto. possono anche capire le parole che sentono e vedono scritte e riescono a percepire i propri errori ma non sono in grado di correggerli. Danni simili si presentano anche quando vi è un danno all’insula. Tali somiglianze di deficit possono essere spiegate con danni ad altre fibre nervose che ancora non si conoscono tra l’area di Wernicke e quella di Broca. 47 Afasia motoriatrans corticale: deriva da una disconnessione tra il Centro dei concetti (B) e l’area di brocca (M). Questa condizione produce sintomi simili a quelli dell’afasia di brocca ad eccezione dell’abilità di ripetere le parole udite che non è compromessa. Afasia globale: vi è una incapacità di produrre e comprendere il linguaggio.è associata ad un danno molto esteso dell’emisfero sinistro che include l’area di Broca, l’area di Wernicke e le regioni intermedie. Sebbene il modello localizzazionista classico spieghi alcuni deficit del linguaggio a oggi utilizzando nuovi strumenti si pensa che il linguaggio sia qualcosa di più rispetto a deficit localizzati ma che nasca da interconnessioni di diverse parti della corteccia. Basi del linguaggio del cervello: Il lessico mentale rappresenta il magazzino mentale delle informazioni riguardanti le parole, nel quale sono contenute l’informazione semantica, l’informazione sintattica e l’informazione relativa la forma della parola. Alcuni modelli teorizzano che il lessico mentale sia unico sia per la comprensione che per la produzione del linguaggio altri invece teorizzano una distinzione fra il lessico in input e quello in output.comunque sia ogni modello deve considerare sia la rappresentazione ortografica basata sulla visione, sia quella fonologica basata sul suono delle parole. Si pensa che nel cervello esistano dei magazzini che già sappiano quale debba essere l’organizzazione concettuale della parola.una volta che le parole sono state analizzate dal punto di vista percettivo si pensa che seguono tre percorsi fondamentali: 1) L’accesso lessicale: elaborazione nel quale l’output dell’analisi percettiva attiva le rappresentazioni sulla forma delle parole nel lessico mentale inclusi i loro attributi semantici e sintattici. 2) Selezione lessicale: è lo stadio successivo dove si identifica la rappresentazione mentale che si adatta meglio all’input. 3) Integrazione lessicale: integra le parole in una frase completa, in un discorso o in un contesto più ampio. rappresenta le regole grammaticali presenti in una determinata lingua da permettere così una comprensione più estesa. Una persona adulta conosce circa 50.000 parole riesce a comprendere senza difficoltà tre parole al secondo, vado a sé che bisogna avere un’organizzazione del lessico mentale molto efficiente. Dunque sicuramente non può essere organizzato in modo alfabetico in quanto andare alla ricerca della parola costerebbe molto tempo. Il lessico mentale è costituito da altri principi organizzativi che ci aiutano a raggiungere la rappresentazione delle parole rapidamente alcuni di essi sono: Morfema: rappresenta la più piccola unità linguistica dotata di significato in una determinata lingua. Il secondo principio organizzativo e che le parole usate con maggior frequenza sono raggiungibili più rapidamente. Un terzo principio organizzativo è l’effetto di vicinanza determinato dal numero di parole che differiscono da un solo fonema. Fonema: la più piccola unità sonora che comporta una differenza di significato. Gli studi hanno evidenziato che parole che presentano più vicini rispetto ad altre vengono identificate più lentamente durante la comprensione linguistica. l’ipotesi è che durante il riconoscimento del discorso si attivino più parole diverse che danno origine a fonemi di competizione. Un quarto fattore organizzativo del lessico mentale e la relazione semantica tra le parole. Questo principio deriva da studi sul priming semantico in cui le persone dovevano seguire un compito di decisione lessicale. Dove si è visto che i tempi di latenza nel pronunciare le parole sono più brevi con parole correlate con l’innesco rispetto a quelle non correlate.(auto-camion) o (tulipano-camion). Dunque le parole associative a livello di significato sono organizzato insieme all’interno del cervello. 48 Modelli del lessico mentale: - Rete semantica: Le parole che costituiscono dei nodi concettuali sono collegate fra loro.la forza della connessione e la distanza fra i nodi sono determinate dalle relazioni semantiche o associative fra le parole. L’attivazione avviene da nodo concettuale ad altro nodo ma i nodi vicini risentono di questa attivazione. Ad esempio se sentiamo la parola automobile si attiveranno delle connessioni anche per le parole camion e automobile. Dunque la parola automobile dovrebbe facilitare il riconoscimento della parola camion ma non di Rosa. Altri modelli affermano che i concetti vengono organizzati tramite le loro caratteristiche o proprietà semantiche. Questi concetti però non riescono a dare una spiegazione convincente su come si attivi l’attivazione.non sono chiari su quante caratteristiche devono essere attivate perché una persona possa riconoscere un determinato oggetto inoltre ad esempio la parola tavolo può essere fatto di legno di vetro in entrambi i casi verrà riconosciuto come tavolo. Un altro problema consiste nel fatto che alcune parole risultano più prototipi rispetto ad altre come ad esempio avviene quando pensiamo alla parola uccello-pettirosso invece che pinguino. Su come sono rappresentati significativamente le parole risulta ancora materia di discussione. Substrati neurali dell’lessico mentale: I pazienti con afasia di Wernicke commettono errori nella produzione del discorso che vengono definiti parafrasie semantiche come ad esempio usare la parola cavallo quando si intende mucca. Pazienti con demenza semantica progressiva mostrano iniziali deficit nel sistema concettuale mentre le altre abilità linguistiche mentali sono risparmiate questo deficit è stato associato a un danno progressivo del lobo temporale e soprattutto nell’emisfero sinistro le regioni superiori del lobo che sono importanti per l’udito e per l’elaborazione del discorso sono però risparmiate. Questi pazienti hanno difficoltà a catalogare gli oggetti a una particolare categoria semantica e tendono a categorizzarle in categorie molto generali ad esempio cavallo animale. Si osservato che si presentavano dei deficit semantici categoria-specifici che avevano una corrispondenza nei siti lesionati.ad esempio i pazienti che avevano danneggiato il riconoscimento degli oggetti animati presentavano lesioni alla corteccia temporale inferiore e mediale spesso nelle regioni anteriori. la corteccia temporale antero-inferiore e vicina alle aree del cervello essenziali per la percezione visiva degli oggetti e al lobo temporale mediale contenente importanti stazioni lungo le proiezioni che connettono la corteccia associativa all’ippocampo che svolge una funzione importante nella codifica delle informazioni nei magazzini di memoria lungo termine. Inoltre il lobo temporale è la stazione terminale della via del “cosa” o del riconoscimento degli oggetti della vista. Sebbene non si sappia molto dei pazienti che presentano deficit riguardanti riconoscimento dei manufatti, sembrerebbe che in questo caso siano coinvolte regioni frontali e parietali sinistre.queste arie sono vicine alle funzioni senso motorie e dunque probabilmente alle rappresentazioni delle azioni che si possono compiere quando si usano oggetti creati dall’uomo.. 49 Sebbene la rete semantica risulti organizzata con una modalità-specifica sono state osservate diverse critiche. per questo Caramazza a proposto una teoria alternativa affermando che la rete semantica è organizzata secondo le categorie concettuali di entità animate ed entità inanimate. Studi su immagini hanno rilevato che i soggetti quando rispondevano a domande su animali o nel leggevano i nomi oppure ne dicevano il nome dopo averne visto l’immagine evidenziavano attivazioni nella porzione più laterale del giro fusiforme (sulla superficie ventrale del cervello) e nel solco temporale superiore. Il denominare un animale attivava anche l’area del cervello associata alle prime fasi dell’elaborazione visiva (lobo occipitale mediale sinistro). Identificare oggetti, utensili attivava invece una regione più mediale del giro fusiforme, nel giro temporale medio sinistro e nell’aria premotoria sinistra. Un’area che diviene attiva anche quando il soggetto immagina movimenti delle mani. Questi risultati dimostrano che effettivamente le rappresentazioni concettuali dell’entità animate e di quelle inanimate come gli utensili sono sostenuti da circuiti neuronali diversi. Studi recenti dimostrano l’esistenza di una rete che connette la parte posteriore del giro fusiforme del lobo temporale inferiore con il lobo temporale anteriore sinistro. Studi recenti hanno osservato che per identificare un essere vivente sono necessarie molte caratteristiche rispetto a un utensile. questo può portare ad affermare che i deficit categoria-specifici nei pazienti possa essere dovuto alla complessità delle caratteristiche che ci aiutano a distinguere le cose dalle altre. Pazienti con lesioni e lobi temporali anteriori non erano in grado di denominare in modo attendibile l’entità viventi a un livello specifico. Studi con immagini derivanti da la risonanza magnetica funzionale su partecipanti sani hanno dimostrato un’attivazione maggiore del lobo temporale anteriore durante la denominazione di entità viventi a livello specifico piuttosto che generale. Inoltre l’attivazione di caratteristiche percettive si verifica nelle cortecce occipitali primarie entro i primi 100 ms dopo la presentazione di una figura. L’attivazione di una rappresentazione semantica più dettagliata si verifica invece nella corteccia ventro laterale posteriore e anteriore tra i 150 e 250 ms. A partire dai 300 ms i partecipanti erano in grado di denominare l’oggetto specifico raffigurato nell’immagine. 50 Comprensione del linguaggio: la comprensione del linguaggio orale e quella scritta condividono alcuni processi ma vi sono anche delle differenze nel modo in cui vengono analizzati i due tipi di input. L’analisi uditiva viene tradotta in un codice fonologico poiché in questo modo sono immagazzinate le rappresentazioni delle forme acustiche delle parole.. Dopo che l’input acustico è stato tradotto in un formato fonologico all’interno del lessico mentale può avvenire l’accesso alla rappresentazione lessicale che meglio corrisponde all’input uditivo (accesso lessicale) in questo modo la parola che meglio corrisponde viene selezionata (selezione lessicale).la forma della parola selezionata attiva poi il livello del Lemma (deposito delle informazioni grammaticali) e poi quello del significato. Infine il significato della parola attiva le informazioni concettuali.la lettura condivide con la comprensione della parola udita almeno gli ultimi due stadi dell’analisi linguistica ovvero l’attivazione del Lemma e significato semantico, differisce però per via della diversa modalità sensoriale in cui viene percepito l’input. Se l’incontro avviene tramite lettura si richiede prima di tutto di identificare l’input visivo e le unità ortografiche. Le unità ortografiche potrebbero essere mappate direttamente sulle forme ortografiche delle parole nel lessico mentale oppure una volta identificate potrebbero essere tradotte in unità fonologiche che a loro volta potrebbero attivare la forma fonologica delle parole nel lessico mentale come avviene nella comprensione uditiva. Imput orale: la percezione dei fonemi varia tra parlanti di lingue diverse. bambini piccoli hanno la capacità percettiva e di distinguere tra ogni possibile fonema entro il loro primo anno di vita. Oltre a comprendere i fonemi il cervello è sensibile ad altre sfumature dei segnali verbali come ad esempio la variabilità del segnale (parlanti di sesso maschile o femminile) dal fatto che i fonemi non rappresentano piccoli pezzi separati di informazioni e inoltre i segnali uditivi a differenza di quelli scritti non sono chiaramente segmentati e questo può portare delle difficoltà a comprendere quando finisci una parola e l’inizio è un altra.quando si parla si usano circa 15 fonemi al secondo.pronunciamo questi fonemi senza discontinuità dunque il segnale di input nel linguaggio parlato e diverso da quello del linguaggio scritto che è costituito da lettere e fonemi nettamente separati. Come il cervello riesca a comprendere il linguaggio in parole separate è stato definito il problema della segmentazione. Il cervello utilizza altri indizi oltre al suono del linguaggio come le informazioni prosodica (ritmo del discorso e intonazione della voce). Le parti del cervello che sono coinvolte nei processi di comprensione del discorso sono la corteccia temporale superiore che ricopre un ruolo importante nella percezione dei suoni. Pazienti con lesioni in quest’aria dimostrano difficoltà nel riconoscere i suoni verbali ma non nell’elaborare gli altri tipi 51 di suoni. Il problema sembra essere limitato al deficit uditivo o fonetico. Oggi grazie alle nuove tecnologie possiamo esplorare le aree specifiche per il linguaggio. I giri di Heschl localizzati nel piano sovratemporale, in posizione superiore e mediale rispetto al giro temporale superiore in ciascun emisfero, contengono la corteccia uditiva primaria ovvero l’area della corteccia che elabora per primo l’input uditivo. le aree che si estendono dai giri di Heshl nel solco temporale superiore costituiscono la corteccia uditiva di associazione. I giri di Heschl di entrambi gli emisferi sono attivati nella stessa misura sia dei suoni verbali sia da quelli non verbali ma l’attivazione del solco temporale superiore di entrambi gli emisferi è modulata dal fatto che il segnale uditivo in ingresso sia o non sia un suono verbale. Più ci allontaniamo dei giri di Heschl verso le porzioni anteriori e posteriori del solco temporale superiore più il cervello diventa meno sensibile a variazioni dei suoni non linguistici e più sensibile a suoni linguistici. dunque le porzioni posteriori del solco temporale superiore sembrano particolarmente rilevanti per l’elaborazione dell’informazione fonologica ma l’elaborazione del linguaggio si estende oltre. Possiamo dunque affermare che il solco temporale superiore da solo non costituisce la sede della comprensione verbale. In uno studio del 2000 alcuni partecipanti ascoltavano suoni linguistici e suoni non verbali. i suoni consistevano in rumori che non contenevano alcuna modulazione sistematica, oppure intoni con frequenze modulate. I suoni verbali invece consistevano in vere parole invertite, pseudo parole e vere parole. Si evidenziò che i toni con frequenza modulata attivavano le porzioni posteriori del giro temporale superiore in entrambi i lobi. Le aree più sensibili ai suoni verbali e non ai toni puri si trovavano in regioni più ventro laterali all’interno o nei pressi del solco temporale superiore. Si ipotizzò anche che queste aree non siano importanti per l’elaborazione semantica in quanto l’attivazione delle parole, delle pseudo parole e delle parole invertite le attivava in ugual misura. Dai risultati di queste di altri studi nasce il modello gerarchico di riconoscimento delle parole. L’elaborazione procede in senso anteriore nel giro temporale superiore. Il flusso dell’informazione uditiva procede dalla corteccia uditiva nei giri di Heschl verso il giro temporale superiore. In quest’aria non viene fatta alcuna distinzione tra suoni linguistici e suoni non linguistici. La prima evidenza di tale distinzione si osserva nella limitrofa porzione mediale del solco temporale superiore anche se in questa regione non vi è ancora un’elaborazione dell’informazione semantica lessicale. Studi neurofisiologici ci dicono che già dai primi 50-80 ms siamo in grado di distinguere se un suono linguistico è una parola o una pseudo parola. Questa elaborazione si presenta più lateralizzata verso l’emisfero sinistro dove vengono combinate le diverse caratteristiche del linguaggio. Dal solco temporale superiore l’informazione procede verso lo stadio finale del riconoscimento delle parole nel giro temporale mediale e nel giro temporale inferiore e infine nel giro angolare dove si trova posteriormente alle aree temporali e più anteriormente verso il polo temporale. 52 53 Import scritto: leggere le parole In più c’è scritto prevede da parte del lettore un riconoscimento del pattern visivo. Il nostro sistema di lettura e di tipo alfabetico in cui si utilizzano dei simboli per rappresentare dei fonemi. Questo processo implica l’analisi visiva di linee orizzontali, vendicati, curve chiuse, curve aperte intersezioni e altre forme elementari. - Nel modello del pandemonium l’input sensoriale viene registrato nella memoria iconica dal così detto demone dell’immagine.28 demoni dei tratti sensibili a caratteristiche elementari come le curve, le linee orizzontali e così via iniziano poi a decodificare i tratti nella rappresentazione iconica dell’input sensoriale. Il secondo passaggio prevede i demoni cognitivi e infine il demone della decisione che seleziona l’input che meglio corrisponde. Questo modello è stato criticato perché prevede solo l’elaborazione guidata dei dati (botto un’app) e non l’elaborazione di un feedback (top Down) come nell’effetto di superiorità della parola.infatti gli esseri umani sono più bravi a identificare lettere che fanno parte di parole più che parole prive di senso. - Modello computazionale a tre livelli: un livello per i tratti delle lettere che compongono le parole, uno per le lettere e uno per la rappresentazione della parola.questo modello presuppone un flusso di informazioni diretto dall’alto in basso che va a influenzare i processi più precoci che avvengono a livelli più bassi.questo modello è differente rispetto all’altro in quanto prevede una via top Down. I due modelli sopra citati rispecchiano due visioni teoriche che sono quello della modularità e dell’interattività. Un’altra differenza fra i due modelli è che nel secondo possono essere elaborate parecchie lettere nello stesso momento in quanto i processi avvengono in parallelo rispetto al primo modello dove ogni lettera viene processata una alla volta. Il secondo modello spiega anche l’effetto di superiorità della parola in quanto presupponendo un’informazione di tipo top Down può essere che tale informazione inibisca le attivazioni delle lettere contribuendo così al riconoscimento della parola piuttosto che della somma delle lettere. 54 Substrati neurali: l’unità ortografica viene elaborata nelle regioni occipito-temporali dell’emisfero sinistro.dando in quest’aria provocano infatti l’Alessia pura ovvero l’incapacità di leggere le parole.mettendo in relazione parole con non parole si trova nella regione della corteccia occipitale una regione che preferiva stringhe di lettere (parole) E venne chiamata area visiva di forma parola. Successivi studi con risonanza magnetica funzionale hanno confrontato i quadri di attivazione cerebrale in risposta a lettere e in risposta a volti o a immagini casuali. Hanno trovato che le regioni occipito temporali della corteccia si attivano preferenzialmente in risposta a stringhe di lettere piuttosto che ad altri input fotografici. Studi successivi hanno osservato che vi era un’attivazione consistente nel solco occipito-temporale sinistro che parte dell’area visiva di forma parola. Si osservava un’analisi playlist cicale che non variava in funzione della posizione spaziale dello stimolo e non era sensibile ad aspetti tipografici. I volti attivavano il giro fusiforme laterale mentre le stringhe di lettere attivano una regione del solco occipito-temporale. Di solito le parole non sono elaborate in modo isolato ma piuttosto nel contesto di altre parole.per comprendere le parole nel loro contesto dobbiamo integrare le proprietà sintattiche e semantiche della parola riconosciuta e generare una rappresentazione complessiva dell’intero enunciato. L’integrazione semantica del significato della parola finale nel contesto della frase ci permette di interpretare l’input nella maniera corretta. Le rappresentazioni contestuali sono cruciali per determinare il senso in cui una parola viene usata.ma bisogna avere anche un’analisi sensoriale altrimenti non ci potrebbe essere rappresentazione del messaggio dovrebbe quindi esiste un momento in cui i due tipi di informazioni si integrano questo momento però non è lo stesso nei diversi modelli. Per spiegare come funziona la comprensione della parola sono stati postulati tre diversi modelli principali: - Modelli modulari: affermano che la comprensione del linguaggio avvenga dall’azione di moduli separati e indipendenti.le rappresentazioni a livello più alto non possono influenzare quella livello più basso e il flusso è di tipo bottom-up. - Modelli interattivi tutti i tipi di informazioni partecipano al riconoscimento della parola. Il contesto può influenzare ancora prima che siano disponibili le informazioni sensoriali modificandone lo stato di attivazione. - Modelli ibridi: le informazioni riguardano le forme delle parole che sono possibili dato il contesto che le precede. Così facendo si riduce il numero dei candidati attivati. In questo modello l’accesso lessicale è autonomo e non influenzato dall’informazioni di livello superiore mentre la selezione lessicale può essere influenzata dall’informazione sensoriale e da quella di livello superiore contestuale. Studi di immagine sul riconoscimento delle parole 55 hanno mostrato che i processi di accesso lessicale e di selezione lessicale riguardo una rete che includa il giro temporale mediale, il giro temporale superiore e la parte ventrale inferiore e bilaterale dorsale del giro frontale inferiore. Hanno osservato che il giro temporale mediale e il giro temporale superiore sono importanti per la traduzione dei suoni del linguaggio parlato in significato. le regioni frontali della corteccia sono importanti nei processi di selezione che il coinvolgimento più rilevante della parte dorsale del giro frontale inferiore si osserva quando la selezione richiedeva di scegliere una parola tra molti possibili candidati (competizione lessicale). Capire una parola nell’ambito della rappresentazione d’ordine superiore del significato della frase implica l’intervento di processi di integrazione semantica e sintattica. Elaborazione semantica è importante per determinare il corretto senso o significato della parola nel contesto della frase. L’informazione semantica tuttavia veicolato da solo dalle parole non è sufficiente per comprendere il messaggio semantica continua nonostante il significato non sia plausibile.. Secondo i principi della psicolinguistica elaborazione della struttura del periodo avviene seguendo delle tappe. Quando sentiamo leggiamo una frase attiviamo le forme delle parole che a loro volta attivano le informazioni semantiche e grammaticali nel lessico mentale. le rappresentazioni delle frasi intere non sono immagazzinate nel cervello come avviene per le parole sintattiche. il cervello dovrà assegnare di volta in volta una struttura sintattica alle parole in entrata. Questo processo viene definito analisi grammaticale e rappresenta un processo di costruzione che non si fonda sul recupero di rappresentazioni delle frasi. Per investigare le basi neurali dell’analisi semantica e sintattica delle frasi sono state utilizzate diversi metodi tra cui quelli elettrofisiologici. Elaborazione semantica e onda N400: È un’onda cerebrale correlata ai processi linguistici. Si tratta di un picco di voltaggio a polarità negativa che di solito raggiunge l’ampiezza massima di 400 ms dopo l’insorgere dello stimolo verbale. Questa onda è sensibile agli aspetti semantici dell’input linguistico. L’il ricercatori misero a confronto l’elaborazione dell’ultima parola di una frase in tre diverse condizioni: 1) Frasi normali che terminano con una parola congruente con il precedente contesto (era il suo primo giorno di lavoro). 2) Frasi che terminano con una parola anomala rispetto al contesto precedente (spalmo sul pane caldo i calzini) 3) Frasi che terminano con una parola congruente al contesto dal punto di vista semantico ma fisicamente deviante (ad esempio con lettere più grandi in caratteri diversi) (lei si mise le scarpe col tacco). Si registra che l’onda N400 avevano ampiezza maggiore in risposta a parole anomale alla fine della frase. Questa differenza di ampiezza dell’onda viene detto effetto N400. Quando la parola era congruente semanticamente ma con un aspetto fisico diverso veniva registrato un potenziale positivo anziché un’onda N400. Tale onda è sensibile anche ad aspetti di comprensione del linguaggio che vanno aldilà delle singole frasi, si trovava infatti anche ad esempio alla fine di una storia. Elaborazione sintattica e onde P600: È conosciuta anche come variazione sintattica positiva.è stata individuata circa 600.000 secondi dopo la presentazione di parole incongrue con le aspettative indotte dalla struttura sintattica. Evocata essenzialmente da frasi che si trovano come nei titoli di notizie scandalistiche ovvero nelle proposizioni note come frasi ingannevoli. Viene definita in inglese come sentiero del giardino, in quanto il giardino è un luogo molto piacevole e colui che viene condotto può essere imbrogliato senza rendersene conto. Quest’onda appare come risposta a molte violazioni sintattiche, si verifica quando i soggetti devono leggere le frasi sia quando le sentono pronunciare. La P600 viene evocata anche da violazioni semantiche in assenza di qualunque violazione sintattica. come ad esempio “Le uova mangerebbero toast e marmellata per colazione) questa frase non contiene errori grammaticali e non è neanche ambigua ma contiene una violazione tematica. l’in questo tipo di frase l’onda 56 risponde in quanto l’analisi grammaticale della struttura della frase viene resa più difficile dalle forti relazioni semantiche. Elaborazione sintattica e riflessa anche da altre onde cerebrali come dall’onda negativa sopra le aree frontali di sinistra detta LAN ( left anterior negativity) che si attiva quando le parole violano la categoria richiesta della frase o quando sono violati i requisiti Moore fossi in tattici. È simile alla N400 ma varia per la distribuzione del voltaggio sul cuoio capelluto. Esistono delle prove che vedono la corteccia frontale inferiore sinistra (area di Broca e zone limitrofe) coinvolta nell’elaborazione sintattica. Altri studi hanno dimostrato che strutture sintattiche più complesse ma si manifestavano con un aumento dell’attività dell’area di brocca. Mettendo in atto studi più complessi che prevedevano periodi più complessi si osservato che oltre all’area di Broca vi erano attivazioni anche nell’area di Wernicke e nelle aree omologhe dell’emisfero destro. È stato osservato anche un’attivazione nella porzione anteriore del Giro temporale superiore. Elaborazione sintattica: corteccia frontale inferiore sinistra e temporale superiore che risultano attive durante l’elaborazione linguistica. Modelli neurali di comprensione del linguaggio: il modello di Hagort di elaborazione linguistica si suddivide in tre parti funzionali che sono: memoria, integrazione e controllo. - La memoria: utilizzata per archiviare recuperare il lessico mentale o memoria lungo termine. - Unificazione: integrazione di informazioni fonologiche, semantiche e sintattiche recuperate dal lessico in una rappresentazione unitaria dll’intero enunciato. Durante la comprensione del linguaggio i processi di unificazione di informazioni fonologiche, semantiche e sintattiche possono avvenire in parallelo questo permette l’interazione tra i diversi tipi di informazioni. - Controllo: riferire il linguaggio ad un’azione. Le proprietà fonologiche e fonetica delle parole sono immagazzinate nella parte centrale o posteriore del giro temporale superiore fino al solco temporale superiore, le informazioni semantiche invece sono distribuite in parti diverse del giro temporale medio e inferiore sinistro. I processi che permettono di combinare di integrare le informazioni fonologiche, semantiche e sintattiche richiamano le aree frontali del cervello come l’area di Broca e il giro frontale inferiore sinistro. Il giro frontale inferiore sinistro è coinvolto in tutti e tre i processi di unificazione: l’unificazione semantica nell’aria 47, l’unificazione sintattica nelle aree 45 e 44 e l’unificazione fonologica nelle aree 44 e 6. La parte del controllo diventa cruciale quando le persone devono ad esempio rispettare i turni per prendere la parola. sebbene il controllo cognitivo non sia stato studiato a dovere le aree coinvolte sembrerebbero la corteccia cingolata anteriore e la corteccia prefrontale dorsolaterale che si attivano durante il controllo cognitivo nella comprensione del linguaggio. Rete linguistica dell’emisfero sinistro: Per la comprensione del linguaggio parlato si possono individuare quattro circuiti: due vie ventrali connettono i lobi temporali posteriori con il lobo temporale anteriore e l’opercolo frontale. Le vie ventrali sono importanti per la comprensione del significato delle parole. Due vie dorsali collegano i lobi temporali posteriori e lobi frontali.la via dorsale che porta la corteccia premotoria è coinvolta nella preparazione del linguaggio.l’altra via dorsale connette l’area di Broca con il giro temporale superiore e il solco temporale superiore (circuito per gli aspetti di elaborazione sintattica) 57 Modelli neurali di produzione linguistica: Innanzitutto bisogna distinguere la comprensione della produzione linguistica. La comprensione linguistica comincia con un imputo orale o scritto che deve essere trasformato in concetto mentre la produzione linguistica inizia da un concetto al quale dobbiamo trovare le parole appropriate per esprimerlo. dunque per produrre il linguaggio il primo passo è quello di preparazione del messaggio. Questo processo è costituito da due aspetti cruciali la macro pianificazione e la micro pianificazione. B è una progettazione del messaggio che costituisce la macro pianificazione. La micro pianificazione invece riguarda il modo in cui le informazioni sono espresse ovvero l’assunzione di un certo punto di vista. la micro pianificazione determina anche la scelta delle parole e loro ruolo grammaticale. Il risultato dell’integrazione della micro pianificazione della macro pianificazione è un messaggio concettuale che forma l’input per un ipotetico dispositivo. il formula Thor che traspone il messaggio nella forma corretta dal punto di vista grammaticale e fonologico. Durante la codifica grammaticale viene computata la struttura superficiale di un messaggio ovvero la sua rappresentazione sintattica ma non quella concettuale.e io oggetti di primo livello della struttura superficiale sono i lemmi.i lemmi sono immagazzinati nel lessico mentale e contengono le informazioni riguardanti le proprietà sintattiche di una parola e le sue specificazioni semantiche e le condizioni concettuali in cui è opportuno usare quella determinata parola. I lemmi sono collegati tra di loro attraverso una rete che li collega sulla base del loro significato. Alla fine sarà individuato un Lemma appropiato per l’immagine presentata. Il processo viene definito selezione lessicale nella produzione linguistica. Delle volte non siamo in grado di attivare la forma sonora di una parola questo viene detto “avere una parola sulla punta della lingua” spesso viene superato con un piccolo suggerimento. Oltre che avere un blocco mentale della parola si può verificare anche errore nella fase di transizione dal livello di Lemma a quello di Lesemma. Può capitare di mischiare i suoni o invertire delle parole in una frase.una volta individuato il lessemma viene coordinato tra i loro programmi fonetici articolatori opportuni.nell’ultimo stadio della produzione della frase viene pianificata l’articolazione.le sillabe infatti sono articolate seguendo degli appositi schemi motori, in questa fase abbiamo la possibilità di correggere qualunque errore sia intervenuto nella preparazione del discorso ad esempio facendo “ mmm mmm” guadagnando tempo per generare il termine appropriato. Gli errori possono verificarsi anche in presenza di danni cerebrali. 58 Visione modulare del modello di Levet I modelli alternativi affermano invece che l’attivazione fonologica incominci subito dopo che sono state attivate le informazioni semantiche e sintattiche relative alle parole e che di fatto si sovrapponga nel tempo. Un’altra differenza è che i modelli interattivi prevedono circuiti a feedback dall’attivazione fonologica alle proprietà semantiche sintattiche delle parole, con conseguente aumento dell’attività azione di certe proprietà sintattiche semantiche. Per comprendere meglio la produzione di parole sono stati registrati risposte di singoli neuroni all’interno dell’area di Broca. I pazienti erano impegnati in un compito che comprendeva tre condizioni che distinguevano processi linguistici lessicali, grammaticali e fonologici. La maggior parte degli elettrodi diede forti risposte elettriche trifasica le risposte onda erano correlate a stadi distinti dei elaborazione linguistica.la prima onda a circa 200 ms sembrava riflettere l’identificazione lessicale; la seconda onda intorno a 320 ms era modulata da aspetti in flessione ali e non dall’elaborazione fonologica. La terza onda appariva a 450 ms e rifletteva la codifica fonologica. È stato osservato che i neuroni motori vengono emessi circa 50-100 ms prima dell’eloquio.gli stadi erano distinti sia a livello temporale che spaziale spostati solo di un paio di millimetri ed erano entrambi collocati nell’area di Broca.i risultati supportano l’ipotesi di un’elaborazione seriale almeno agli stadi iniziali di produzione del linguaggio. Altri studi hanno osservato l’attivazione dell’opercolo sinistro, potrebbe essere collegato alla codifica fonologica della produzione verbale. L’articolazione delle parole coinvolge probabilmente la parte posteriore dell’area di brocca e dell’insula.studi con la PET e la risonanza magnetica funzionale hanno osservato che le componenti motorie del linguaggio con volgono la parte opercolare del giro precentrale, le regioni posteriori del giro frontale inferiore, l’insula, il distretto della bocca della corteccia motoria sensoriale primaria, i gangli della base, e talamo e il cervelletto. 59 Evoluzione del linguaggio: la selezione naturale favorisce gli individui che mettono richiami per influenzare il comportamento di chi è in ascolto e di coloro che acquisiscono le informazioni apprese dalle vocalizzazioni ascoltate. In generale i richiami vocali dei primati tendono essere involontari e associati a uno stato emotivo specifico. Al contrario i gesti sono flessibili e vengono utilizzati in situazioni di non urgenza per dare la via ad azioni come giocare o spulciarsi e vengono appresi socialmente. L’utilizzo dei gesti richiede di conoscere lo stato attentivi del compagno. i gesti dei primati sono flessibili e socialmente appresi e richiedono un’attenzione condivisa. alcuni sostengono che linguaggio si sia evoluto dalla comunicazione gestuale. Alcuni ipotizzano che il linguaggio sia nato da gesti e movimenti facciali e che è un neuroni specchio facessero parte di questo puzzle. I neuroni specchio sono stati scoperti nell’area effe cinque della scimmia che si occupa del movimenti della mano mentre la porzione ventrale di quelli della bocca e della laringe. l’aria effe cinque della scimmia è omologa all’area di Brod manna 44 una porzione dell’area di brocca nell’essere umano. Molti studi associano il linguaggio a gesti manuali anche negli uomini. Le aree che controllano i movimenti manuali e le vocalizzazioni sono localizzate molto vicine in strutture omologhe sia nelle scimmie che negli esseri umani. 60 Cap 12 controllo cognitivo Il controllo cognitivo ci permette di utilizzare le nostre percezioni, conoscenze e scopi per orientare la selezione dell’azione e dirigere pensione attraverso una moltitudine di possibilità. Ci permettono di prevalere sui comportamenti processi automatici e di muoversi al di fuori dell’ambito di risposta abituato. Il controllo cognitivo e indispensabili ai fini di un comportamento orientato a uno scopo e dei processi decisionali. All’interno della corteccia prefrontale troviamo due sistemi di controllo. il primo include la corteccia prefrontale laterale e il polo frontale che supporta il comportamento orientato a uno scopo. Anatomia alla base del controllo cognitivo: Gli essere umani i lobi frontali comprendono circa un terzo della corteccia celebrale. Il confine posteriore e segnato dal solco centrale ed è l’unico solco che si estende per l’intera lunghezza della superficie laterale del cervello e discende lungo quella mediale. il lobo frontale è separato da quello temporale dal solco cerebrale laterale o scissura di Silvio. la parte più posteriore del lobo frontale comprende l’area motoria primaria e comprende la circonvoluzione situata anteriormente alla scissura di Rolando. In posizione enteroventrale vi sono le aree motorie secondarie tra cui la corteccia premotoria laterale e l’area motoria supplementare. Il resto del lobo frontale è detto corteccia prefrontale negli esseri umani comprende metà dell’intero lobo. La corteccia prefrontale: troviamo quattro regioni; corteccia prefrontale laterale; polo frontale; corteccia orbitofrontale detta delle volte zona ventro mediale e infine la corteccia frontale mediale. Negli esseri umani la corteccia prefrontale è molto più estesa. Si riscontra un aumento a livello di materia bianca piuttosto che materia grigia. Questo porterebbe a pensare che vi sia un aumento di connessioni piuttosto che di un numero di neuroni. l’espansione dei lobi frontali è correlata all’emergere umani. La corteccia prefrontale inoltre matura più tardi in termini di sviluppo. La corteccia prefrontale collabora con molte regioni del sistema nervoso centrale. È in collegamento con le regioni motorie, percettive e limbiche del cervello. La corteccia prefrontale riceve informazioni da quasi tutte le regioni della corteccia parietale e di quella temporale e da qualche regione della corteccia occipitale. gli input maggiori provengono dal talamo che connette la corteccia prefrontale con le regioni sottocorticali inclusi i gangli della base e il cervelletto e nuclei tronco-encefalici. La corteccia prefrontale a sua volta in via proiezioni alla maggior parte delle aree di cui riceve afferenze compresa l’area motoria e quella premotoria. In via proiezioni anche alle aree prefrontali dell’emisfero controlaterale tramite il corpo calloso e proiezioni B laterali alle regioni premotorie e Sub corticali. Il sistema dopaminergico include l’aria tegmentale ventrale, un nucleo tronco-encefalico, i gangli della base e la regione dorsale e ventrale dello striato. 61 - grediente ventrale-dorsale: mantenimento e manipolazione del materiale. Gradiente antero-posteriore: rappresentazioni astratte, carico cognitivo. Gradiente laterale-mediale: integrazione informazioni per memoria e comportamento finalizzato. Danni alla corteccia frontale può portare diversi danni che non sempre vengono riconosciuti. Il più comune è la perseverazione (continuare a dare una risposta nonostante si dica che sia sbagliata). Possono presentare anche apatia, difficoltà a comprendere le conseguenze delle azioni o impulsività. Mancherebbe il comportamento orientato allo scopo. Il comportamento è guidato dallo stimolo. I soggetti con danno ai lobi frontali mantengono una conoscenza sull’uso prototipo degli oggetti come ad esempio utilizzare il martello per appendere un chiodo, vedendo lo stimolo in questione rispondono di conseguenza (appendono il quadro) non sono tuttavia in grado di inibire la loro risposta o cambiarla con flessibilità sulla base del contesto in cui si trovano appare così evidente che il loro meccanismo di controllo cognitivo e fuori uso. Comportamento orientato allo scopo: si distinguono due importanti tipologie di azioni. 1) Azioni dirette a uno scopo: si basa sulla ricompensa e sulla consapevolezza di una relazione causale tra l’azione e l’eventuale ricompensa “azione-esito”; 2) Abitudine: è un’azione non controllata da ricompensa ma guidata dallo stimolo. Viene considerata automatica. Le azioni abituali si presentano davanti alla presenza di specifici stimoli che attivano delle associazioni. Vi è però la presenza del controllo cognitivo che fa da mediatore tra il comportamento e gli scopi del comportamento. Ci permette di focalizzarsi sull’informazione rilevante e monitorare i nostri comportamenti per arrivare all’obiettivo. il controllo cognitivo richiede anche la memoria di lavoro. ovvero una memoria breve termine utile per rappresentare le informazioni rilevanti per il compito. La corteccia prefrontale laterale è il primo luogo in cui vi è un’interazione tra le informazioni percettive e le conoscenze immagazzinate in memoria; rappresenta un punto importante nella memoria di lavoro. la corteccia prefrontale è importante per il controllo cognitivo. È stata indagata per capirne l’importanza nella memoria di lavoro utilizzando il compito di risposta ritardata. Alla scimmia viene fatto vedere dove viene posto il cibo poi viene abbassata la grata, fatto passare un intervallo di tempo e la scimmia è in grado di ricordare dov’è il cibo questo dimostra che l’animale ha delle rappresentazioni mentali di dov’è il cibo. Con lesioni alla corteccia prefrontale laterale il compito è molto scarso. Quando venivano invece messi dei cartoncini che suggerivano la posizione del cibo (memoria associativa) danni alla corteccia prefrontale laterale non portavano a un peggioramento della prestazione ovvero gli animali riuscivano ad individuare attraverso il suggerimento è una posizione corretta del cibo. Il compito di risposta ritardata l’animale deve tenere in memoria la risposta per un periodo di tempo, nell’apprendimento associativo l’associazione viene riattivata dal suggerimento. L’associazione dei lobi frontali nella memoria è stata dimostrata anche negli esseri umani. Ad esempio si è visto una difficoltà nell’organizzare le azioni secondo una sequenza temporale appropriata. una donna riportava di non riuscire a preparare la cena nonostante si ricordasse tutti gli ingredienti. In bambini di 1 anno difficoltà perché cortecce non sviluppate. 62 L’associazione tra memoria di lavoro e intelligenza è stata studiata prendendo in considerazione sia la materia grigia che quella bianca (connettività). ma oltre a un maggior volume si sono distinti anche un tipo di intelligenza cristallizzata che si riferisce alla conoscenza e un tipo di intelligenza fluida che si riferisce alla capacità di problem solving. È stato visto anche che la capacità di mantenere il focus attentivi su un’informazione rilevante per un compito di memoria di lavoro era il punto cruciale dell’intelligenza fluida. il neurotrasmettitore dopamina ha un ruolo importante sull’apprendimento si è visto che in euro recettori dopamina sono molti nella corteccia prefrontale. Si pensa che abbiano una funzione modulatori affinando la risposta dei neuroni della corteccia prefrontale. la performance dell’apprendimento è influenzata quindi dalla presenza di dopamina che però segue un andamento tipico di una funzione a U-rovesciata. questo si vede anche nella malattia di Parkinson Dove vi è una riduzione dei livelli di dopamina nello striato dorsale e quando viene somministrata vi è un effetto overdose nello striato ventrale e nel lobo frontale in quanto prima in queste zone era a livelli consoni. Si sono visti anche degli alleli che influenzano i livelli di dopamina e come ad esempio il (COMT) che produce un enzima che distrugge la dopamina. Ritornando al compito di risposta ritardata, I ricercatori hanno misurato l’attività di singoli neuroni scoprendo che alcuni di questi cominciavano ad attivarsi subito dopo l’inizio del periodo di ritardo ovvero quando lo stimolo non è più visibile. queste cellule consentono una registrazione continua della risposta che l’animale deve produrre per ottenere la ricompensa. Sì scoperto anche che le cellule della corteccia prefrontale sono selettive per certi attributi dello stimolo e che tale selettività è specifica per il compito. sono state individuate cellule “What”, “Where” e “What-Where”. Queste cellule si attivano in maniera selettiva sul cosa, dove o combinazione di entrambi dello stimolo. un’altra alternativa e che l’attivazione delle cellule della corteccia prefrontale rappresenti una rappresentazione delle finalità del compito e che mettere in relazione le nostre rappresentazioni a lungo termine rilevanti per il compito con altri sistemi neurali. questa ipotesi richiama il fatto che la corteccia prefrontale è connessa con regioni sensoriali posteriori della corteccia temporale e di quella parietale. quando viene percepito uno stimolo nella corteccia prefrontale viene attivata una rappresentazione temporanea tramite queste connessioni con le regioni posteriori del cervello, questo facilita il comportamento orientato allo scopo. Questo è stato evidenziato da studi di neuro immagine funzionale che hanno utilizzato una variante del compito di risposta ritardata. Ai partecipanti venivano mostrate sia dei volti completi che dei volti frammentati. Inoltre vi era una variazione di numero dei target. successivamente dopo un tempo di latenza di otto secondi veniva presentato un volto detto “probe”. Ai partecipanti veniva chiesto di decidere se il probe ma corrispondeva a uno dei volti presentati nella fase precedente. Si sono registrate delle risposte BOLD nella corteccia prefrontale laterale all’inizio del periodo di codifica. Questa risposta rimane anche per tutto il periodo di ritardo. Sì osservato anche che la risposta aumenta in relazione al numero di stimoli presentati in precedenza. discriminatoria dell’utilizzato dei volti hanno constatato anche che vi era un’attivazione della corteccia prefrontale e dell’area fusiforme per i volti nel lobo temporale inferiore. La registrazione delle risposte BOLD nelle due regioni ha evidenziato che durante la fase di codifica la risposta era maggiore nell’area fusiforme per i volti piuttosto che nella corteccia prefrontale. durante il periodo di risposta le risposte della corteccia prefrontale risultavano alte. tuttavia durante questo periodo si registrava comunque una risposta anche nell’area fusiforme dei volti a testimonianza che rimaneva comunque attiva. Si registrò un effetto di rimbalzo. Dunque sebbene vi sia una risposta piccola dell’area fusiforme dei volti questa rimane comunque attiva rispetto a quando lo stimolo non è di tipo facciale. Si è registrato anche una risposta più precoce nell’area fusiforme dei volti durante il periodo di codifica, più precoce invece è stata la risposta nella corteccia prefrontale durante il periodo di recupero. Questo ha dimostrato che la corteccia prefrontale laterale è importante per la memoria breve termine poiché mantiene una rappresentazione degli scopi del compito e interagisce con la corteccia infero temporale per mantenere tali informazioni durante il periodo di ritardo. Queste informazioni sono pertinenti al conseguimento dell’obiettivo. 63 La memoria del lavoro è responsabile della manipolazione in vista di particolari scopi comportamentali. Uno dei modelli di memoria di lavoro afferma che l’informazione conservata nella corteccia posteriore è attivata, recuperate mantenuta nella corteccia prefrontale ventro laterale e qui rielaborata insieme alle informazioni rilevanti in regioni più dorsale della corteccia prefrontale laterale così da permettere il comportamento più efficace per raggiungere lo scopo. In un disegno sperimentale è stato osservato che quando il soggetto veniva chiesto di compiere un compito che utilizzasse la memoria di lavoro in avanti trovavano meno difficoltà di quando veniva chiesto loro una condizione “all’indietro”, questo perché la rappresentazione degli elementi in questo caso doveva essere ricordata e manipolata. Si è osservata una risposta BOLD nella regione della corteccia prefrontale laterale simile in tutte le condizioni, mentre la risposta della corteccia prefrontale dorso laterale è la più alta nelle condizioni all’indietro. questo ha portato a confermare che la corteccia prefrontale dorso laterale è fondamentale quando i contenuti della memoria di lavoro devono essere rielaborati. per studiare questo si utilizzano di solito i test “n-back”. Si è osservato anche che l’attivazione della corteccia prefrontale laterale aumenta in funzione delle difficoltà del compito più “N” si chiede di individuare maggiore sarà l’attivazione. Corteccia prefrontale ventrale-dorsale: mantenimento e manipolazione delle informazioni. Ci si è chiesto la parte frontale quale scopo avesse. Sembrerebbe essere indispensabile per integrare i contenuti specifici dell’attività mentale in schemi più generali.si è evidenziata la presenza di una gerarchia grossolana della corteccia prefrontale, i compiti più elementari che richiedono la memoria di lavoro attivano solamente la corteccia più posteriore o anche le aree motorie secondarie. qualora il compito diventasse più impegnativo come ad esempio oltre che a visualizzare la forma tener conto anche del colore, si evidenziano attivazioni anche nel polo frontale E se le richieste dovessero portare a dei cambiamenti di regole si osserva un’attivazione ulteriore anche in direzione anteriore. Ne deriva che la corteccia prefrontale può essere organizzata lungo i tre assi distinti: 1) Gradiente ventrale-dorsale organizzato in termini di mantenimento e manipolazione delle informazioni. riflette anche i principi organizzativi generali osservati nella parte più posteriore della corteccia come le vie visive ventrali e dorsali per il “cosa” e il “dove”. 2) Ingrediente antero-posteriore dove vi è un’attivazione durante le rappresentazioni più astratte che coinvolgono regioni più anteriori come ad esempio il polo frontale. Si potrebbe dire che il lobo frontale, la corteccia motoria primaria siano le regioni in cui le intenzioni astratte vengono tradotti in movimenti concreti. 3) Gradiente laterale-mediale: è legato all’influenza dell’ambiente rispetto alla memoria di lavoro o alle informazioni legate alla storia personale e agli stati emotivi, le prime la porzione più laterale le seconde quella più mediale. Queste regioni integrano le informazioni provenienti dall’esterno che sono rilevanti ai fini del comportamento orientato a uno scopo 64 con le motivazioni e la possibile ricompensa che ne potrebbe derivare. L’integrazione permette così di regolare il comportamento. Processi decisionali: Mettere in atto un comportamento orientato allo scopo presuppone l’aver preso una decisione per perseguire l’obiettivo. queste decisioni possono seguire diverse teorie: - Teorie decisionali normative: corrispondono alle decisioni che le persone dovrebbero prendere perché risultano la scelta più ottimale. - Teorie decisionali descrittive: descrivono ciò che le persone fanno effettivamente e non quello che dovrebbero fare. Spesso le nostre decisioni risultano in coerenti per il mondo che ci circonda (come mangiare un dolce quando si è a dieta) ma bisogna considerare che il nostro cervello funziona considerando un punto di vista evoluzionistico. Le decisioni possono essere orientate a uno scopo oppure abitudinari. Le prime considerano una ricompensa mentre le seconde non sono più sotto il controllo della ricompensa. un altro modo per classificare le decisioni e quello di dividerle in: - Decisioni che risultano in un’azione: sono quelle decisioni che valutano l’esito atteso. - Decisioni stimolo-risposta quando il processo decisionale diventa abituale. Un’altra distinzione: - Decisioni basate su modello: il soggetto possiede una rappresentazione interna di qualche aspetto del mondo e lo utilizza per valutare diverse azioni. - Prive di modello: vi è solo una mappa d’ingresso-uscita simile a quella della decisione stimolo-risposta. - Le decisioni che coinvolgono altre persone sono dette decisioni sociali. Uno dei concetti più importanti dei processi decisionali E il calcolo che soggetti fanno su ogni singolo valore delle opzioni che hanno disposizione facendo successivamente un confronto tra i diversi valori. Il processo decisionale riguarda il fare scelte in grado di massimizzare il valore e dipende da situazioni esterne, interne ed integrazioni e confronto. Abbiamo diversi tipi di ricompense, alcuni premi sono considerati dei rinforzi primari “cibo, acqua” e producono dei benefici diretti alla sopravvivenza fisica. Il valore è la risposta che diamo a questi rinforzi e calibrata dal nostro codice genetico e modellata dall’esperienza. I rinforzi secondari “Denaro o status sociale” non sono di per sé dei rinforzi ma portano a delle gratificazioni secondarie. Il valore dei rinforzi è dato dall’integrazione tra fattori esterni ed interni e va a costituire un valore personale complessivo. Bisogna dunque calcolare: la ricompensa, la probabilità che questo avvenga, gli sforzi e i costi, il deprezzamento temporale, il contesto e le preferenze. Questa 65 complessità porta a prendere delle decisioni differenti che per alcuni possono essere razionali e per altri no. Gli scienziati si sono chiesti quali fossero le aree del cervello responsabili della ricompensa. Utilizzando elettrodi multipli posizionati nella corteccia frontale della scimmia (corteccia cingolata anteriore, corteccia prefrontale laterale e corteccia orbitofrontale). Si è valutato utilizzando un esperimento in cui una scimmia doveva prendere una decisione guardando delle immagini.ad ogni immagine era associato un valore specifico su tre dimensioni: costo della ricompensa, entità e probabilità di ottenerla. (l’immagine della chitarra associata a tre gocce di succo di frutta) il valore della scelta veniva manipolato lungo un’unica dimensione mantenendo le altre due costanti ad esempio quando veniva manipolato il costo, le immagini e la probabilità della vincita erano costanti. Si è visto che ogni aria aveva delle cellule che rispondevano a una determinata dimensione manche cellule che rispondevano in combinazione. non era presente una netta separazione ma +1 sfumatura. molte cellule della corteccia cingolata anteriore E della corteccia orbitofrontale rispondevano a tutte e tre le dimensioni. Le cellule della corteccia prefrontale laterale codificavano di solito solo una variabile decisionale. L’attività celebrale si attivava prima, “in preparazione” suggerendo una risposta motoria, della risposta. studi su gli umani attraverso l’uso della risonanza magnetica funzionale hanno osservato come diverse dimensioni riescono ad attivare in maniera preferenziale alcune regioni neurali. la corteccia orbitofrontale risultò legata alla variabilità nella ricompensa; lo striato dei gangli della base allo sforzo. La corteccia prefrontale laterale alla probabilità della ricompensa e la corteccia prefrontale mediale e nel lobo parietale laterale erano sensibili al ritardo tra il momento in cui veniva eseguita l’azione e la ricompensa. Si è osservato anche che la corteccia prefrontale ventro-mediale compresa la corteccia orbitofrontale si attivava in base alla preferenza per il gusto mentre la corteccia prefrontale dorso laterale era legata più al grado di controllo.ad esempio più attiva quando si rifiuta un cibo buono ma poco sano Meno attiva quando il cibo malsano veniva scelto. Dunque la corteccia prefrontale dorsolaterale potrebbe essersi evoluta per decisioni e gratificazioni a lungo termine. Modulando così i segnali che arrivano dalla corteccia prefrontale ventro mediale. La corteccia orbitofrontale hanno ruolo chiave nella rappresentazione del valore. Mentre le regioni più laterali della corteccia prefrontale sono importanti per il controllo modulatori. Teorema del valore marginale: sfruttare una zona per foraggiarsi finché il suo tasso di raccolta scende al di sotto del tasso medio per l’ambiente nel suo complesso. a questo punto il comportamento dell’animale diventa esplorativo. Si pensa che tale decisione abbia aspetti biologici ed è soggetta a delle variabili decisionali. Il livello in cui viene attivata la decisione è soggetto a diversi fattori come ad esempio i costi e benefici. Studi sulle scimmie hanno osservato che l’attività neurale nella corteccia cingolata anteriore era correlata la capacità di monitoraggio dell’azione e dei suoi risultati. In accordo con il teorema del valore marginale gli animali erano propensi a scegliere lo stimolo associato a una certa ricompensa se tempi d’attesa aumentavano o se la qualità del cibo diminuiva. L’attività della corteccia cingolata anteriore era predittiva per calcolare il tempo che l’animale avrebbe speso per continuarsi a nutrire scegliendo lo stimolo più gratificante. Le cellule presentavano una soglia ovvero quando la frequenza di scarica era superiore a 20 pixel secondo l’animale abbandonava l’apprezzamento. Dunque la corteccia cingolata anteriore e correlata alla 66 decisione delle scimmie di passare ad una fase esplorativa dell’ambiente alla ricerca di alternative migliori rispetto al corso attuale dell’azione. Le ricompense soprattutto quelle dei rinforzi primari come il cibo e il sesso sono fondate su comportamenti molto antichi per questo ne consegue che aree più vecchie si attivino durante le azioni di ricompensa come ad esempio aree sottocorticali che sono coinvolte nella rappresentazione della ricompensa, lo striato dorsale ventrale, l’ipotalamo, l’amigdala e l’abenula laterale. Molti studi hanno osservato che il neurotrasmettitore dopamina è importante negli studi sulla ricompensa. anche in verità la serotonina. sebbene le cellule dopaminergica siano presenti in tutto il mesencefalo in due regioni se ne trovano di più; sono: In due nuclei principali del tronco-encefalico: - Substantia nigra pars compacta; - Area tegmentale ventrale. Le proiezioni avvengono: - Substantia nigra --> proietta allo striato dorsale (principale nucleo di input dei gangli della base) troviamo una perdita di neuroni E nei pazienti con la malattia di Parkinson. - Area tegmentale (VTA)segue 2 vie: 1) Via mesolimbica- cruciale per le emozioni proietta al: nucleo accumbens (striato ventrale) dei gangli della base, all’amigdala, l’ippocampo e la corteccia cingolata anteriore. 2) Via mesocorticale: proiezioni di tipo dopaminergica verso la neocorteccia nello specifico nella porzione mediale del lobo frontale. Il ruolo della dopamina sulla ricompensa è stato scoperto durante esperimenti sui ratti a cui erano stati messi degli elettrodi. alcuni topi che avevano degli elettrodi che stimolavano una via dopaminergica tendevano a abbassare una levetta per la ricompensa in maniera ossessiva mentre gli altri no. – recettori domaminergici eccitatori – D1 _ recettori dopaminergici inibitori-D2 Si è visto che utilizzando tecniche più raffinate come l’optogenetica per stimolare i recettori dopaminergici nello striato alcuni topi premevano la levetta e altri no. questo dipendeva che tipo di recettore attivava l’elettrodo. gli scienziati hanno anche compreso che l’attivazione dei neuroni dopaminergici non è legata alla ricompensa in sé ma all’aspettativa. è stato osservato che quando la ricompensa è inaspettata l’attivazione risulta più forte. Il ruolo della dopamina nella ricompensa è prevedere l’errore ovvero la discrepanza tra la ricompensa ottenuta e quella attesa. l’aperta parentesi esperimento con le scimmie di tipo comportamentista con stimolo condizionato succo di frutta e stimolo incondizionato luce) dunque i neuroni dopaminergici nella via mesocorticale rispondono maggiormente quando la ricompensa non è attesa. La risposta dopaminergica varia anche a seconda della relazione di ritardo tra lo stimolo condizionato e lo stimolo incondizionato.se lo stimolo condizionato. Per quanto riguarda invece l’associazione della dopamina la punizione la ricompensa è stato osservato che nell’abenula (Struttura del talamo dorsale) si trovano proiezioni che riguardano stati emotivi e motivazionali. Si osservato che i neuroni di quest’aria mostrano una dipendenza dal contesto. Dipende da come la ricompensa viene vista nel contesto ad esempio se viene offerto un po’ di succo oppure niente quest’area si attiverà quando viene effettuata la scelta che non produce niente oppure anche nel caso di una risposta avversiva. Cv per alcuni l’apprendimento avviene con il rilascio della dopamina per altri invece la dopamina sarebbe il risultato e non la causa dell’apprendimento questo perché è stato osservato che nei topi in cui veniva tolta la dopamina erano comunque capaci di apprendere.per alcuni la ricompensa è formata da tre elementi il volere, la prendere e il piacere e la dopamina risponde solamente al volere. Rimane comunque che la dopamina a molteplici funzioni ne vengono descritte principalmente due: -un sottogruppo di neuroni dopaminergici risponde in termini valenza (stimoli predittivi di una ricompensa diminuiscono in risposta a stimoli avversi); (risposta errori di previsione) - un altro gruppo di neuroni dopaminergici rimaneva eccitato in qualsiasi tipo di rinforzo sia che si trattasse di un premio che di una posizione soprattutto quando era imprevedibile. (risposta ad eventi 67 che richiedono attenzione). Questo ci riporta alla corteccia del lobo frontale che partecipa sia a sistemi decisionali sia a comportamenti orientati allo scopo. Pianificazione degli scopi: Per poter pianificare bisogna Sviluppare un piano d’azione prendendo in considerazione più sotto-obiettivi possibili. Gli obiettivi dunque possono essere considerati in maniera gerarchica. Durante il compito della competizione della risposta i soggetti sono stati chiamati a rispondere a quesiti sempre più complessi. È stato osservato che quando il compito diventava più complesso venivano attivate regioni più anteriori della corteccia prefrontale mentre quando il compito è la più semplice come ad esempio scegliere soltanto il colore dello stimolo si attivava la zona della corteccia premotoria. quando il compito risultava più complesso vi era anche un’attivazione più estesa della corteccia prefrontale e del polo frontale anteriori mostrano dei pattern e di attivazione più specifici. La corteccia premotoria (A) era sensibile a tutti e quattro i compiti; la corteccia premotoria anteriore (B) era sensibile al compito della caratteristica, della dimensione e del contesto; il solco frontale inferiore (C) era sensibili ai compiti della dimensione del contesto e la corteccia frontopolare (D) era sensibile e unicamente al compito del contesto. Il comportamento orientato allo scopo richiede che vi sia una selezione delle informazioni rilevanti per il compito specialmente sulle caratteristiche e sulle informazioni in memoria. La corteccia prefrontale può essere considerata come un filtro dinamico che permette le proiezioni reciproche tra la corteccia prefrontale e la corteccia posteriore consentendo in questo modo di mantenere in memoria informazioni rilevanti per il compito che richiedono conoscenze a lungo termine e che sono dunque immagazzinate nella parte posteriore della corteccia. Per confermare il ruolo dinamico filtrante della corteccia prefrontale si è osservato che durante compiti di generazione di verbi i soggetti che dovevano richiamare alla memoria verbi legati a un’immagine che verrà presentata avevano una maggiore attivazione di momenti in cui l’input richiamava più verbi più più che pochi verbi come il vedere le forbici (tagliare). Ai soggetti poi venne chiesto anche di riprodurre sia il 68 nome dell’azione associata al sostantivo che dire il colore a esso associato. Si osservò che le aree prefrontali e temporali venivano attivati e che l’aria temporale inferiore era reclutata per entrambi i compiti di associazione semantica. Quando le richieste venivano cambiate si evidenziò un aumento di risposta nella corteccia prefrontale ma una diminuzione in quella temporale. Quando il filtro dinamico si lesiona si perde si osserva che i pazienti con danni alla corteccia prefrontale abbiano difficoltà specialmente in occasioni in cui vi sono molte informazioni di mantenere l’attenzione. Sì osservato anche che i pazienti con lesioni alla corteccia prefrontale abbiano difficoltà a recuperare un suggerimento utile per rispondere al compito.questo si è visto con un esperimento in cui prima i soggetti dovevano rispondere leggendo una lettera o il numero presentato su un cartoncino nel il secondo compito invece veniva utilizzato un suggeritore “colore” ad esempio a ogni cartoncino rosso bisognava leggere la lettera e a quello verde il numero. I soggetti con lesioni alla corteccia prefrontale avevano tempi di risposta molto più lunghi se non sbagliati questo perché la corteccia prefrontale è utile non solo a coordinare i comportamenti orientati allo scopo ma anche a recuperare la memoria di lavoro. infatti I soggetti devono ricordare che al verde è associato il numero. Controllo cognitivo basato su un obiettivo: Bisogna comprendere quali sono i meccanismi neurali attraverso i quali si ottiene il controllo basato su un obiettivo. bisogna comprendere le interazioni di cortecce anche distanti fra loro. Per studiare come gli attivazione della corteccia prefrontale influisca su regioni posteriori gli scienziati hanno cercato di indagare nello specifico la regione dei campi oculari frontali importante nell’attenzione spaziale. Questa regione lavora insieme al collicolo superiore nel controllo dei movimenti degli occhi. Questo perché l’attenzione richiede un equilibrio tra il mantenimento del focus attentivi e l’orientamento verso stimoli salienti nuovi. La regione dei campi oculari frontali è importante per la capacità di sopprimere la tendenza a guardare stimoli nuovi. Si è osservato che la risposta risultava attenuata nella visione centrale piuttosto che laterale. Suggerendo che il controllo basato su obiettivi operato dalle regioni dei campi visivi aiuta a mantenere la concentrazione per questo i segnali percettivi provenienti dalla fovea venivano attenuati. Danni ai lobi frontali producono una difficoltà se non la perdita del controllo inibitorio.ad esempio durante un esperimento in cui ai soggetti veniva chiesto di prestare attenzione a uno stimolo acustico inviato ad un orecchio e ad ignorare suoni simili dell’altro. Nei soggetti normali le risposte arrivavano dopo 100 ms cosa che non avviene nei pazienti con lesioni prefrontali. lo stimolo 69 disatteso riceve anche una risposta più intensa. La capacità di inibire stimoli irrilevanti è utile anche durante compiti di memoria di lavoro in cui stimoli possono disturbare la capacità di tenere a mente l’informazione utile. Utilizzando esperimenti che attivino regioni del lobo temporale inferiore in particolare l’area fusiforme e quella ippocampale utilizzando stimoli coinvolti in luoghi si osservato che quando hai soggetti veniva rilevato l’obiettivo del compito si registrava una modulazione percettiva dell’informazione enfatizzando quelle utili ed inibendo quelli rilevanti. Si fece questo esperimento anche con partecipanti più anziani e si osservò che la risposta eccitatoria rimaneva alta mentre risultava difficile quella di inibizione. Con questo si può pensare che la corteccia prefrontale sia la prima a subire l’invecchiamento. Per osservare che effettivamente la corteccia prefrontale intervenisse il sistema eccitatorio e di inibitorio si utilizzò stimolazione magnetica trans cranica sulla corteccia frontale e si vede che la corteccia frontale inferiore è importante per inibire le informazioni irrilevanti per il compito mentre la corteccia frontale dorsale è importante per enfatizzare l’elaborazione di informazioni rilevanti. Inibizione dell’azione: l’inibizione e dunque utile per il controllo cognitivo. anche inibire un’azione che già è stata pianificata è molto comune. la capacità di inibire l’azione è stata studiata attraverso il compito del segnale di stop. le risposte a questo compito possono essere di tre tipi: 1) Prove senza segnale di stop; 2) Prove in cui la persona in grado di arrestare in tempo il comportamento; 3) Prove in cui la persona non riesce a fermarsi in tempo. Si è osservato che durante le prove efficaci ed inefficaci vi era una risposta del giro frontale inferiore destro mentre nelle prove senza segnale di stop non era presente. Questo dimostra la presenza di un processo inibitorio anche quando questo non va a termine. Gli scienziati si sono chiesti anche come mai delle volte la risposta può essere inibita e altre no e hanno studiato la corteccia motoria. si osservato una risposta della corteccia motoria molto elevata nelle risposte Go e nelle prove di arresto ed efficace. Vi è dunque un’elevata attivazione pre-stimolo. La corteccia prefrontale destra pur cercando di inibire l’attivazione non è capace di inibirla del tutto. Si è osservato anche un’attivazione del giro frontale inferiore destro presente nel nucleo subtalamico dei gangli della base che sono implicati nella via della risposta. Il nucleo subtalamico fornisce un forte segnale eccitatorio al globo pallido contribuendo a mantenere l’inibizione della corteccia. l’attivazione della corteccia prefrontale destra genera il comando per interrompere una risposta e questo comando viene implementato reclutando il nucleo subtalamico.vi è dunque una connessione tra la corteccia frontale inferiore destra e l’area supplementare premotoria e il nucleo subtalamico dei gangli della base. L’area supplementare motoria è una parte della corteccia frontale mediale che si attiva quando c’è un conflitto di risposta.bisogna tenere in considerazione che la stimolazione profonda del nucleo subtalamico che viene fatta per agevolare il movimento al paziente Parkinson può portare ad essere troppo impulsivi perché inibisce il sistema di controllo inibitorio. 70 Cv controllo dell’efficacia del comportamento orientato allo scopo: utilizza un modello per dimostrare che la selezione dell’azione è un processo competitivo. Modello della selezione della risposta proposto da Shallince e Norman: le azioni sono associate a unità di controllo di schemi, che possono riguardare diverse abilità come il significato semantico o le rappresentazioni associative. Le unità di controllo di schemi ricevono degli imputo da molte fonti hanno un’importanza Maggiore input percettivi.la selezione di questi input viene comunque pilotata dal conflitto regolato dal sistema attentivi superiore. Gli autori hanno suggerito che esistono due tipi di selezione degli imput: 1) uno passivo ma veloce detto conflitto regolato: in quanto gli schemi non sono solo guidati da input percettivi ma competono tra di loro soprattutto quando sono esclusive e l’uno dell’altro.ad esempio non si può seguire due oggetti in movimento. dunque è uno solo schema che riesce a vincere la competizione. Se non avviene la soluzione del conflitto non vi sarà azione in quanto nessuno degli schemi sarà attivato in maniera sufficiente per innescare una risposta. 2) Sistema attentivi o supervisore: permette al comportamento di essere flessibile. favorisce il controllo di alcuni schemi così da poter seguire le esigenze specifiche della situazione e dare rilevanza ad alcuni obiettivi piuttosto che ad altri. Le informazioni di controllo fanno parte di una rete celebrale ben distribuita. negli anni una parte di grande interesse è stata occupata dalla corteccia frontale mediale e specialmente dalla corteccia cingolata anteriore che sembrava attivarsi e fungere da supervisore. in verità si osservato che la corteccia frontale mediale soprattutto quella cingolata si attivavano quando il compito diventava più difficile e si richiedeva un maggior monitoraggio. In realtà sono state individuate solamente all’interno della corteccia cingolata anteriore 11 sottoregioni. Che proiettano maggiormente in alcune aree del cervello e altre regioni in altri. Sicuramente è importante la posizione che occupa.sono state fatte molte ipotesi per spiegare il ruolo della corteccia frontale mediale. La prima ipotesi è quella della gerarchia attentiva in cui l’rispetto alla condizione di controllo in cui soggetti devono operare vi è un numero di attivazione di regioni specifiche della corteccia. Inoltre il legame tra corteccia frontale mediale e l’attenzione viene anche dal fatto che in questa regione vi è un cambiamento al diminuire delle richieste attenzionali. Ad esempio nel compito della ricerca dei verbi quando al soggetto viene chiesto di trovare dei verbi a una nuova parola vi è un compito più impegnativo di associazione e vi è un’attivazione maggiore. Cosa che invece non vi è quando viene chiesto di rifare il compito in questo caso il soggetto andrà ad utilizzare informazioni già presenti. Una seconda ipotesi è quella del rilevamento dell’errore: È stato rilevato che quando si commette un errore vi è una risposta negativa che si diffonde sulla corteccia prefrontale dopo l’inizio del movimento. questo segnale viene indicato come negatività correlata all’errore o risposta ERN. Le risposte fisiologiche della negatività correlata all’errore potrebbero essere utili per riattivare l’obiettivo sperimentale memoria di lavoro che magari avevamo perso perché distratti. Ipotesi del conflitto di risposta: Le situazioni nuove di solito generano un conflitto di risposte e gli errori sono situazioni in cui esiste per definizione un conflitto. Utilizzando il compito di Stroop I ricercatori hanno visto che viene un’attivazione maggiore nella corteccia prefrontale laterale quando il compito veniva reso più difficile anche prima che lo stimolo venisse effettivamente presentato. La corteccia frontale mediale invece era sensibile al grado di conflitto di risposta e risultava maggiore quando la parola e il colore dello stimolo erano diversi. dunque la corteccia prefrontale laterale rappresenta l’obiettivo del compito mentre quella frontale mediale se l’obiettivo viene raggiunto oppure no. A differenza del modello di rilevamento dell’errore che affermava che l’aria se ti va se soltanto in presenza di errori in questo si afferma che l’aria è impegnata ogni volta che vi è un conflitto e la probabilità che si verifichi l’errore. 71 72 Cognizione sociale cap 13 E condizioni sociali è un’aria importante di ricerca perché studia i meccanismi che supportano le capacità complesse di una persona di comprendere gli stati mentali interni di un’altra persona. uno degli obiettivi centrali della ricerca e studiare come riusciamo a mentalizzarci a vicenda. Osservazioni su pazienti che presentavano danno alla corteccia frontale hanno rilevato che vi era un cambiamento del comportamento e della personalità tanto da non essere più riconosciuti. Substrati anatomici della cognizione sociale: - Aree della corteccia prefrontale: faccia anteriore del lobo frontale. - La corteccia prefrontale si divide in corteccia prefrontale dorsale e corteccia prefrontale ventro laterale. - Le regioni mediali sono la corteccia orbito frontale e la corteccia prefrontale ventro-mediale. Le regioni coinvolte nell’elaborazione delle se sono la corteccia prefrontale ventro-mediale, la corteccia prefrontale dorso laterale E la corteccia cingolata anteriore, l’insula e al di fuori della corteccia che includono il sistema nervoso autonomo, l’asse ipotalamo-ipofisi-surrene e sistemi endocrini che regolano gli stati corporei, le emozioni e la reattività. Poiché anche la memoria è coinvolta nell’azione autoreferenziale di un coinvolgimento anche del lobo temporale. Quindi a seconda del compito possiamo riscontrare attivazione dell’amigdala e delle sue interconnessioni con il solco temporale superiore, la corteccia prefrontale mediale e la corteccia orbito frontale, la corteccia cingolata anteriore dell’area fusiforme dei volti. Inoltre vi è l’attivazione di sistemi dei neuroni specchio, poli temporali, la giunzione temporoparietale la corteccia parietale mediale. Alcune persone presentano dei deficit nella capacità sociale come ad esempio avviene per l’autismo. L’autismo è un disturbo dello sviluppo caratterizzato da un comportamento sociale anomalo.le persone autistiche si concentrano spesso su la autostimolazione e mostrano poco interesse per le azioni degli altri individui una delle ipotesi è che queste persone abbiano delle difficoltà nella capacità di comprendere gli stati mentali degli altri “cecità mentale” 73 Lamenta lizzazione sono i processi cognitivi che permettono una persona di fare inferenze accurate e rapide sugli stati interni di un’altra persona. Le parti del cervello coinvolte nella mentalità azione sono: - corteccia prefrontale mediale; - Giunzione temporo-parietale; - Cingolo precuno-posteriore; - Amigdala; - Solco temporale superiore; - Poli temporali. L’imperativo socratico conosci te stesso: Quando percepiamo noi stessi in realtà colui che percepisce e il percepito coincidono. Secondo il modello di memoria dei livelli di elaborazione la profondità dell’elaborazione influisce profondamente sulla memorizzazione delle informazioni. Questo vuol dire che le informazioni elaborate in maniera più significativa vengono ricordate meglio di altri. Inoltre l’effetto autoreferenziale ci dice che l’informazione elaborata in relazione a noi stessi si ricordano meglio degli altri. Per spiegare questo effetto sono state fatte due ipotesi la prima afferma che la concezione del sé attivi una struttura cognitiva unica dotata di elementi di memoria e di organizzazione unitari che elaborano le informazioni in modo diverso rispetto a tutte le altre strutture cognitive; l’altra ipotesi nega l’esistenza di un’area specifica e afferma che noi abbiamo semplicemente una maggiore coscienza del sé e questo porta a codificare le informazioni in modo più elaborato. L’elaborazione autoreferenziale è collegata anche un’attivazione della corteccia prefrontale mediale. Inoltre si è notato che quando dobbiamo autodescriverci tendiamo a fare affidamento a percezioni che sono delle sintesi dei nostri tratti di personalità. Se dobbiamo descrivere un’altra persona utilizziamo invece degli esempi particolari. Scienziati hanno riscontrato che la corteccia prefrontale mediale si attiva anche quando il cervello è a riposo. In questa zona del cervello affluiscono notevoli quantità di sangue rendendo quest’operazione molto costosa. Per alcuni questo è dovuto al fatto che mentre il cervello a riposo continua essere impegnato in una serie di processi psicologici che descrivono una modalità funzionale cerebrale predefinita. le regioni che supportano questi processi sono chiamate rete funzionale predefinita o rete di default. Rete di default: formata da corteccia prefrontale mediale, dal precuneo, dalla giunzione tempo parietale, dal lobo temporale mediale, dalla corteccia parietale laterale e dalla corteccia cingolata posteriore. Si ipotizza che la sua funzione sia quella di garantire sempre un’idea di ciò che ci sta succedendo attorno questa è detta “ipotesi della sentinella”. È collegata al sistema di memoria del lobo temporale mediale questo potrebbe spiegare il perché di riflessioni spesso del passato. la rete si disattiva quando bisogna mettere in atto un’attività specifica. La rete di default si attiva quando vengono messi in atto diversità compiti autobiografici come di memoria autobiografica, di pensare assistessi nel futuro o in un viaggio e così via. L’attivazione di quest’aria porta la disattivazione di altre aree e non soltanto per un risparmio di costo ma anche perché in questo modo siamo più abili nel mettere in atto il compito. Non sarebbe proficuo pensare ad esempio al pane che brucia all’interno del tostapane mettersi nei suoi panni. Quindi sebbene gli esseri umani siano naturalmente predisposti a pensare agli stati mentali propri e a mettersi nei panni degli altri per interagire in maniera adeguata con aspetti non sociali del nostro ambiente dobbiamo disattivare la rete di default durante l’esecuzione di compiti non sociali. 74 Noi siamo più propensi a pensare di avere più probabilità di altri di sperimentare eventi futuri positivi piuttosto che negativi. Questo è spiegato perché regioni prefrontali specifiche di ordine superiore permettono le persone di concentrarsi in maniera selettiva su aspetti positivi di se stessi evitando loro di deviare troppo lontano dalla realtà. studi suggeriscono che la parte più ventrale della corteccia cingolata anteriore sia responsabile della focalizzazione dell’attenzione su informazioni positive riguardo il sé. È stato osservato che quest’aria si attivava maggiormente quando veniva chiesto ai soggetti di scegliere degli aggettivi considerati autodescrittivi che erano positivi piuttosto che negativi. In un altro studio la corteccia cingolata anteriore si attivava quando i partecipanti immaginavano di vivere un evento positivo futuro rispetto a un evento negativo.dunque la corteccia cingolata anteriore è importante per distinguere informazioni positive relative al sé da quelle negative. La percezione comunque rimane realistica e questo consente di seguire le norme sociali.i pazienti invece con danni alla corteccia orbitofrontale tendono ad avere una visione di se stessi non realistica positiva con un comportamento sociale inadeguato. I pazienti con danni alla corteccia orbitofrontale hanno difficoltà ad avere una percezione di sé accurata e spontanea ma nel momento in cui vengono messi davanti ai loro atteggiamenti ad esempio facendo riguardare loro un’intervista si rendono conto degli errori. Questo porta a pensare che siano a conoscenza delle norme sociali ma che vi sia una mancanza di intuito su come usarle. Quando dobbiamo pensare a un evento futuro che non c’è mai capitato tendiamo a proiettarci nell’esperienza cercando di prevenirla.gli studi hanno evidenziato come la corteccia presentare il mediale nella parte più ventrale fosse continuamente impegnata, questo supporta l’idea che questa zona della corteccia sia utile per le previsioni relative alle specifiche simpatie e antipatie di un individuo. Quando è presente un inganno le persone tendono a preferire in maniera incoerente. Questo è stato osservato durante un compito in cui le persone dovevano scegliere chi preferivano tra due attori. Delle volte però mettiamo in atto delle scelte miopi questo perché questa zona tende a attivarsi in maniera più intensa quando dobbiamo prevedere di ricevere piacere da un evento nel presente rispetto a quando si giudicano eventi futuri. Osservando il livello di attivazione gli esperti potevano anche prevedere quanto i soggetti erano più propensi a scegliere eventi futuri più favorevoli oppure a mettere in atto delle scelte nuove (20 € oggi 23 la settimana prossima) e maggiore era l’attivazione mentre si predicevano eventi futuri meno venivano prese decisioni miopi. Teoria della mente: l’accuratezza empatica si riferisce alla precisione con cui un partecipante in grado di dedurre i pensieri e sentimenti di una persona Target. Più di confidenza con quella persona maggiore sarà l’accuratezza empatica. Questo perché comprendere gli altri è uno dei principi fondamentali che alla base della cooperazione umana. La capacità di dedurre gli stati mentali degli altri viene detta appunto teoria della mente. I bambini ad esempio preferiscono guardare un volto umano piuttosto che altri oggetti. anche a quattro mesi neonati mostrano un’attività gamma precoce evocata sui siti occipitali ed esplosioni di un ritmo gamma tardivo sui siti della corteccia prefrontale destra in risposta al contatto visivo diretto. (Studi ERP). Per molto tempo si è creduto che la teoria della mente avesse delle basi innate ma che si sviluppasse nei primi 10 anni di sviluppo del bambino acquisendo competenze sempre più elaborate.ad esempio il bambino a 12 mesi di età che conoscono la posizione di un oggetto dichiarano dove si trova lo getto ma smettono di dichiararlo si 75 comprendono che la persona ha già questa informazione, o ancora bambini di 15 mesi mostra una sorpresa quando qualcuno cerca l’oggetto in un contenitore secondo loro sbagliato. a 17 mesi comprendono che la persona ha una falsa credenza e così via. In verità alcuni autori ipotizzano che la teoria della mente sia del tutto innata e automatica. Questo è stato studiato attraverso l’esperimento in cui veniva fatto vedere uno schermo è una palla. i possibili scenari erano quattro e i soggetti partecipanti due. Uno era la gente e l’altro osservava. è stato preso in considerazione il loro tempo di reazione alla fine dell’esperimento e si è osservato che l’osservatore influenzava comunque la risposta del partecipante suggerendo che la teoria della mente sia innata e che la sola presenza di un altro essere umano faccio a scattare automaticamente l’analisi delle sue credenze. Questi risultati si sono avuti anche in bambini di sette mesi di età. Gli scienziati si sono chiesti come riusciamo a dedurre pensieri degli altri traducendo ciò che è osservabile ovvero il comportamento in un’inferenza su ciò che non è osservabile ovvero il suo stato psicologico. Si sono ipotizzate due teorie principali: - Teoria della simulazione: detta anche teoria del sistema di condivisione delle esperienze. Afferma che mentre osserviamo il comportamento dell’altro e noi vi è una certa reazione che innesca uno stato psicologico e presupponiamo che questa stessa reazione sia uguale a quella che sta percependo quella persona. Possiamo fare questo o in maniera inconscia coinvolgendo il sistema di rispecchiamento simile ai sistemi di neuroni specchio coinvolti nelle azioni dirette oppure in maniera consapevole mettendoci nei panni di qualcun altro. Questa teoria però non spiega come sia possibile percepire l’altro anche a distanza o quando mette in atto un comportamento opposto al suo stato interno. Questa teoria ipotizzando un “mettersi nei panni degli altri” dovrebbe portare all’attivazione della corteccia prefrontale mediale associata alla percezione sia del sé che delle altre persone. Questo è stato studiato chiedendo ai partecipanti di osservare due descrizioni in cui vi era una persona simile alla persona che guardava e lancia molto diverse. Questi studi portarono ad evidenziare un’attività in una zona più dorsale rispetto alla corteccia prefrontale mediale quando la persona non era simile e una più ventrale della corteccia prefrontale mediale quando era simile. Studi successivi hanno evidenziato che le risposte più ventrali o dorsali non dipendevano dalla somiglianza rispetto al sé ma il grado di relazione tra le due persone. Aree della corteccia prefrontale mediale simili ai primi esperimenti si attivavano anche quando il partner doveva percepire la sua relazione amorosa. - La seconda ipotesi è la teoria della teoria: detta anche teoria del sistema di attribuzione degli stati mentali propone che possiamo dedurre gli stati mentali degli altri partendo da ciò che sappiamo su di loro compresi i ricordi sugli altri, le situazioni in cui si trovano, la famiglia, la cultura e così via. Un concetto importante è quello di empatia ovvero la capacità di capire le emozioni e le esperienze uniche di un’altra persona.per poter rispondere in maniera adeguata dobbiamo essere capaci di percepire in maniera accurata le informazioni emotive da questi trasmesse. Secondo il modello della percezione-azione dell’empatia questo avviene in maniera automatica innescando risposte automatiche e somatiche. Si presuppone anche che i neuroni specchio siano un meccanismo fisiologico cruciale che ci permette di avere la stessa rappresentazione dello stato interno di un’altra persona all’interno del nostro corpo. questo viene detto anche “simulazione incarnata” negli esseri umani è stato visto che l’esperienza di disgusto e la percezione delle espressioni facciali di disgusto attivano regioni simili nell’insula anteriore. Un altro studio osservare le reazioni dell’insula in un paziente con dei danni in quest’aria e si osservò che non era capace di riconoscere il disgusto. Altri studi hanno visto che l’insula e il cingolato anteriore si attivano quando si prova dolore fisico su se stessi e quando si percepisce dolore fisico degli altri. Pazienti con la corteccia somatosensoriale selezionata mostravano difficoltà ad identificare lo stato emotivo dell’altro. Questi studi dimostrano che esistono diverse regioni della corteccia che si attivano quando sperimentiamo uno stato interno o quando lo osserviamo. 76 Per comprendere come capiamo quando siamo noi a provare il dolore o quando lo serviamo sono stati fatti degli esperimenti. È stato osservato che l’insula anteriore veniva attivata sia quando si valutavano le informazioni su di sé oppure su persone vicine ma non quando si valutavano dei lavoravano informazioni su personaggi pubblici.questo ha portato a pensare che quando elaboriamo le informazioni su di noi sulle persone vicine le sentiamo a livello fisiologico e le rappresentiamo mentalmente influenzando la prospettiva emotiva di ciascuna persona.questo processo viene detto “consapevolezza incarnata”. L’elaborazione specifica per sé viene associata alla corteccia cingolata anteriore e ventrale e alla corteccia cingolata anteriore dorsale che non si attivavano quando si chiedeva di valutare altre persone vicine oppure personaggi pubblici. La corteccia cingolata anteriore dorsale è coinvolta nel meccanismo neurale diretta a un fine che richiede uno sforzo, è coinvolta anche nella regolazione dell’attenzione e nell’auto consapevolezza e anche nel monitoraggio dell’azione. Inoltre si è visto che all’interno della corteccia prefrontale mediale viene un delle attivazioni diverse per se stessi, per le persone vicine e per gli altri personaggi pubblici. L’attivazione per il sé era raggruppata soprattutto nella corteccia prefrontale mediale ventrale di destra; le persone vicine era raggruppata principalmente nella corteccia prefrontale ventrale mediale di sinistra; la corteccia prefrontale mediale dorsale invece era coinvolta nei personaggi pubblici soprattutto a livello del giro frontale superiore sinistro. Vi è anche una modulazione delle risposte empatica, dove le intenzioni e l’autoregolazione influenza il grado di esperienza empatica e la probabilità di comportamenti che risultano positivi per gli altri. Cioè la regola azione è diretta a un fine. Come avviene ad esempio per i dentisti o per gli agopuntori. In questi mesi è stato osservato che mentre osservavano delle immagini di agopuntura si attivavano la corteccia prefrontale mediale e la giunzione tempo parietale destra mentre in persone che non praticavano l’agopuntura si attivavano regioni associate all’esperienza di dolore come l’insula, il cingolo anteriore e la corteccia sommato sensoriale. Dunque l’attività di neuroni specchio può essere modulata. Si è vista anche una differenza di risposta a seconda se osserviamo una persona che consideriamo giusta o se pensiamo che quella persona un po’ se l’è meritato.si è vista una riduzione di risposta nel secondo caso. Queste risposte attivavano in maniera maggiore lo striato e il nucleo alcumbens ventrale che si attivano in caso di ricompensa. Se invece la punizione veniva considerata ingiusta vi era un’attivazione maggiore dell’insula e della corteccia cingolata anteriore. Attribuzione di stati mentali: - Corteccia prefrontale mediale: quando si formano delle impressioni si vede un’attivazione della corteccia prefrontale mediale maggiore di quando si effettua un giudizio sequenziale.si è dunque compreso che la corteccia prefrontale mediale gioca un ruolo importante nella formazione di impressioni sugli stati interiori di altre persone ma no su i tipi di informazioni riguardanti un’altra persona. È presente una risposta maggiore quando si fanno impressioni su esseri animati piuttosto che essere inanimati. - La giunzione tempo parietale destra: anche questa regione è stata associata nel fare inferenze sugli stati mentali degli altri. È stato osservato che vi è un’attivazione dell’area fusiforme 77 - per i volti. Per individuare bene questa regione all’interno della giunzione tempo parietale destra si è utilizzato il compito delle false credenze (che indaga la teoria della mente). Altri studi hanno indagato l’attivazione di quest’aria ponendo i soggetti dinanzi a informazioni di contesto sociale, stato mentale ed eventi della vita; è stata evidenziata una risposta superiore alle informazioni sugli stati mentali. Interazione tra la corteccia prefrontale mediale e la giunzione temporale destra richiama molte ipotesi. una di queste dice che la giunzione temporale destra si attivi per gli stati mentali mentre la corteccia prefrontale mediale supporti i ragionamenti sui compiti sociali includendo anche gli stati mentali ma non solo quelli… una seconda ipotesi afferma che la corteccia prefrontale mediale supporta i ragionamenti sui compiti sociali e la giunzione tempo parietale destra sia importante per indirizzare l’attenzione nei compiti sociali. Gli ultimi studi sulla giunzione tempo parietale destra hanno evidenziato che questa si divida in due parti dove da una parte vi sono neuroni che si attivano nei compiti di mentalizzazione e dall’altra si attivano per orientare l’attenzione sia su compiti sociali che non sociali. Il solco temporale superiore: integrazione dei segnali non verbali con gli stati mentali. I segnali verbali come il linguaggio del corpo la postura e l’espressione del viso sono importanti per percepire lo stato di una persona. Vi è un’attivazione dell’area fusiforme per i volti ma anche dell’amigdala. Nel primo anno di vita i bambini sviluppano l’attenzione congiunta e la capacità di monitorare l’attenzione di un’altra persona. Questo si osserva perché il bambino tende a osservare dove guarda l’altra persona. Lo sguardo è utile anche per capire quando le parole non corrispondono agli stati d’animo delle persone.si è osservato che le cellule del solco temporale superiore aiutano nell’identificare la posizione della testa e la direzione dello sguardo. il solco temporale superiore si trova al di sotto del giro temporale superiore e al di sopra del giro temporale medio. Alcune delle cellule rispondono alla posizione della testa mentre altre rispondono alla direzione dello sguardo. Questi due tipi di risposte permettono di individuare le differenze. Il solco temporale superiore è importante per interpretare lo sguardo in relazione agli stati mentali. Sì osservato che è una regione del solco temporale superiore variava in relazione alla direzione degli spostamenti dello sguardo.se lo sguardo veniva spostato su uno spazio vuoto la risposta dell’aria era più duratura rispetto a quando lo sguardo si spostava verso il target. È stato osservato anche che l’aria era sensibile al tempo che si impiegava per spostare lo sguardo. 78 L’autismo: I bambini con autismo si osserva la nascita una circonferenza del capo inferiore seguita da un brusco rigonfiamento nel primo anno di vita. Per questo si pensa che le anomalie cerebrali siano dovute ad una riduzione del volume di più aree cerebrali tra cui i lobi frontali, il solco temporale superiore, l’amigdala, il cervelletto e l’ippocampo. Si è osservato anche una connettività iper sviluppata nelle regioni del lobo frontale e una diminuita nella connettività a lungo raggio e delle alterazioni reciproche tra le altre regioni corticali. Altre regioni che vengono implicate nell’autismo sono la corteccia prefrontale mediale, l’amigdala, l’lare fusiforme per i volti, il solco temporale superiore, l’insula anteriore e la giunzione temporoparietale destra. I compiti delle false credenze sono molto difficili per i bambini autistici anche oltre l’età in cui normalmente si sviluppa questa capacità in bambini autistici invece completano la storia come sei personaggi avessero tutte le informazioni. Durante la ricerca è stato scoperto che la giunzione tempo parietale destra funzionava in modo anomalo e. quest’aria si attivavano le persone non autistiche quando pensavano a pensieri più che a giudizi fisici. Nei soggetti autistici invece quest’aria non si attivava. Si è riscontrato che minore era la selettività maggiore era la compromissione mostrata dell’individuo nella capacità di rappresentarsi gli stati mentali altrui. E noto che i bambini autistici non tendono a guardare negli occhi. Uno studio finlandese ha notato che in verità sono sensibili all’estensione della sclera cioè la parte bianca dell’occhio. Ma l’ipotesi è quella che non riescono a codificare lo sguardo (l’espressione) per avere notizie riguardo lo stato d’animo degli altri.vi è presente anche una minore attivazione del solco temporale superiore in compiti di teoria della mente ma vi è un’attivazione più estesa in più tipologie di condizioni. questo potrebbe spiegare che vi è anche una difficoltà nello spostare lo sguardo. Per spiegare la difficoltà a spostare lo sguardo può essere evidenziata anche l’amigdala più piccola. Inoltre si è osservato che non vi è una inibizione della corteccia prefrontale mediale cioè della rete di default. non vi è dunque nessun cambiamento tra lo stato di riposo e quello di esecuzione del compito. Sembrerebbe che questa sia sempre disattivata. Il non doversi preoccupare della risposta sociale potrebbe essere il motivo per il quale gli autistici sono molto abili nella risoluzione del compito come ad esempio risolvere il puzzle o calcoli matematici. Alcuni hanno ipotizzato anche un deficit dei neuroni specchio. Lo studio italiano ha individuato che i bambini autistici non presentavano l’attivazione del muscolo Milo io ideo e durante un compito in cui veniva chiesto loro di raggiungere un pezzo di cibo, afferrarlo e mangiarlo. Il muscolo si attivava solo nell’ultimo passaggio al contrario nei soggetti non autistici l’attivazione era già presente nelle prime fasi del raggiungimento comprendendo così il fine del movimento. La comprensione delle intenzioni altrui può avvenire in due modi diversi: nel primo ci si basa su informazioni derivate dalle interazioni manuali con gli oggetti, il secondo utilizzando informazioni semantiche derivate dal normale uso degli oggetti o del contesto in cui vengono utilizzati. i bambini autistici non presentano difficoltà nel secondo. Ma nel primo. Conoscenza sociale: la ricerca evidenzia l’importanza dei lobi frontali per prendere in considerazione la situazione particolare al fine di applicare le regole appropriate. in quanto il comportamento sociale non ha regole definite e chiare ma molto dipende dalla situazione. La corteccia orbitofrontale si è danneggiata porta a difficoltà nel dare un senso all’interazioni sociali. Per studiare questo fenomeno è stato formulato il compito delle gaffe sociali dove si è notato che le persone che presentavano danni orbitofrontale non erano in grado di usare la loro conoscenza sociale per ragionare sulle intenzioni sociali. Si registra una totale mancanza di consapevolezza che può portare le persone con danni in quest’aria a non comprendere in modo diverso in cui poter agire. La sensazione di imbarazzo è utile per evitare di sentirci nuovamente in quel modo. La corteccia orbitofrontale è importante sia per l’apprendimento della conoscenza sociale sia per l’applicazione aspecifica interazioni sociali. Di prendere decisioni: prendere decisioni non può non tenere in considerazione l’utilizzo della coscienza sociale. è stato osservato nei primi studi che i pazienti con danni alla corteccia prefrontale ventro-mediale risultavano incapaci di prendere decisioni di tipo sociale. i primi studi vennero fatti utilizzando il gioco d’azzardo. Sì osservato che le persone con danni alla corteccia orbitofrontale 79 non apprendono dall’esperienza negativa ovvero non avviene l’inversione dell’apprendimento. Ci si è però chiesti come fosse possibile che la corteccia prefrontale ventro-mediale potesse sebbene lesionata a prendere il valore dello stimolo iniziale ma non l’inversione dello stimolo quando si arrivava a una risposta negativa. Si osservato dunque che la corteccia orbitofrontale in realtà non era sensibile al valore dello stimolo ma piuttosto segnalasse la violazione all’amigdala dell’aspettativa sul valore.i pazienti che hanno partecipato a questo studio avevano dei danni al lobo frontale ventro-mediale che si trova in una regione che comprende sia la corteccia orbitofrontale mediale sia la corteccia prefrontale ventrale e mediale. studi anche su pazienti normali e pazienti con danni frontale dorsolaterale. è stato chiesto loro di svolgere un compito di apprendimento probabilistico con feedback sia positivi che negativi. È stato osservato che nel lobo frontale ventromediale vi era la capacità di apprendere da un feedback negativo ma non da uno positivo mentre soggetti con danni alla corteccia frontale dorsolaterale erano in grado di imparare dal feedback sia positivo che negativo. Neuro economia: integrazione di psicologia, neuroscienze, economia e modelli computazionale per comprendere come le persone prendano decisioni basate sul valore. l’affermano che le persone prendono decisioni razionali cioè massimizzando i benefici e minimizzando le perdite. La ricerca dimostrato che le persone che prendono decisioni basate su principi finanziari hanno un’attivazione maggiore della corteccia orbitofrontale mentre persone che prendono decisioni basandosi più sulle emozioni avevano un’attivazione maggiore dell’amigdala. Valutando ed analizzando il gioco dell’ultimatum in cui un giocatore deve offrire del denaro a un secondo giocatore e il secondo giocatore deve accettare oppure non l’offerta, in caso la rifiutasse nessuno dei due avrebbe il denaro si osservato che: quando la divisione non era equa vi era un’attivazione della corteccia prefrontale dorso laterale e dell’insula quest’ultima associata a emozioni negative. L’attivazione dell’insula era anche predittiva della probabilità che l’offerta sarebbe stata respinta. Sebbene la risposta finanziariamente corretta è quella di accettare qualsiasi proposta quando l’offerta era ingiusta si tendeva a rifiutarla per non compromettere la posizione sociale. Respingendo l’offerta punite l’altro giocatore che non riceve nulla ottenendo così status sociale ponendo i giocatori sleali. In studi che indagano la cooperazione vi era un’attivazione delle aree associate alla ricompensa come il nucleo alccunbens, la corteccia orbitofrontale, il cingolato anteriore e nucleo caudato. Queste aree vengono attivate anche in mancanza di ricompense dirette in compiti prosociali. Decisioni morali: dilemmi morali la maggior parte delle persone sarebbe d’accordo nel deviare il tram nel dilemma morale del tram per salvare cinque persone sacrificandone una.ma non sarebbero d’accordo nello spingere lo sconosciuto dal cavalcavia per fermare il tram ed evitare che esso uccida le persone. La differenza potrebbe stare nel livello di coinvolgimento nel causare la morte di una persona. Le decisioni impersonali attivavano la corteccia prefrontale laterale destra e il lobo parietale bilaterale cioè aree associate alla memoria di lavoro. I dilemmi personali invece attivavano aree associate a processi cognitivi emozionali e sociali. Dunque le risposte ai dilemmi sociali dipendono da quanto da come permettiamo alle nostre mozioni di influenzare le nostre decisioni su ciò che è moralmente accettabile. 80 Parte di sviluppo Riassunti slide 1 Per studiare lo sviluppo bisogna assumere un punto di vista dinamico e non statico. Il processo di sviluppo inteso come una sequenza di trasformazioni e non come un semplice incremento. alcune caratteristiche tendono a rimanere costanti nel tempo manifestandosi in maniera precoce mentre altre possono estinguersi o variare. Durante lo studio di sviluppo bisogna analizzare tre livelli di indagine: - Cambiamento cerebrale - Cambiamento comportamentale - Cambiamento mentale (strutture e funzioni.) Bisogna inoltre prestare attenzione all’interpretazione dei dati. L’approccio neurocostruttivista interpreta in maniera diretta il difetto genetico e l’aspetto cognitivo. La neurodiversità e dunque lo sviluppo atipico non è carente o in ritardo rispetto a quello tipico ma è diverso perché segue un percorso evolutivo diverso. Molte teorie per studiare lo sviluppo fanno a pezzi e riducono la complessità del soggetto di studio. Le teorie cognitive classiche spiegano le strutture dell’attività cognitiva ma anche le funzioni di tale attività dunque spiegano sia la natura strutturale del sistema cognitivo nelle diverse età sia quali processi vengono utilizzati per costruire ed elaborare le conoscenze. Oggi si va verso nuovi orientamenti cercando di superare la dicotomia strutturale verso funzionale. L’attività cognitiva diventa dunque il prodotto del cervello che si caratterizza per avere una sua specifica architettura è una sua specifica modalità di funzionamento. Oggi questo è possibile anche grazie ai nuovi metodi di ricerca come le registrazioni ERP, sEMG, NIRS, eye-tracker. Slide2. Sviluppo embrionale e fetale del cervello umano - 18 giorni dal concepimento l’embrione inizia impiantarsi nella parete uterina ( È formato da tre strati di cellule endoderma, mesoderma e ectorderma quest’ultimo si inspessisce e porta allo sviluppo della piastra neurale) - 20 giorni il solco neurale inizia svilupparsi - A 22 giorni il solco neurale si chiude lungo la lunghezza dell’embrione che forma un tubo; alcuni giorni più tardi si osservano quattro divisioni principali del cervello che sono: -telencefalo – diencefalo – rombencefalo -lopment (della piastra neurale) 81 8 fasi dello sviluppo embrionale: 1) Mitosi/ proliferazione: si verifica nella zona ventricolare. Nella prima fase una cellula staminale genera neuroblasti. Nella seconda fase, subisce una specifica divisione a simmetrica in cui cambia dal generare i neuroni alla glia. 2) Migrazione: la differenziazione avviene mentre neuroni migrano. Le cellule gliali radiali fungono da fili guida per la migrazione dei neuroni. I coni di crescita avanzano lentamente trainando gli azioni alle loro spalle. La loro dimensione è influenzata dall’ambiente esterno che alla fine li indirizza verso i bersagli appropriati. All’estremità troviamo delle sottili estensioni filiformi dette filopodi. I neuroni per orientarsi nella direzione seguono segnali provenienti da diverse fonti estrinseci. 3) Sensazione: in questa fase neuroni diventano fissi e specializzati sviluppando assoni e dendriti e la conduzione elettrica. Dopo la migrazione e la differenziazione abbiamo 6 strati corticali. 4) Aggregazione: neuroni in questa fase si muovono insieme formando gli strati. 5) Sinaptogenesi: gli assoni formano una sinapsi con altri neuroni o tessuti come ad esempio i muscoli. Le cellule bersaglio rilasciano una sostanza chimica che crea una sfumatura attorno a loro. I coni di crescita orientano e seguono il gradiente verso la cellula. esistono però anche delle proteine come la chemopellent Slit che rispinge la maggior parte degli assoni. 6) Morte neurale: tra il 40 e il 75% di tutti i neuroni nati nello sviluppo embrionale e fetale non sopravvive.spesso perché non riescono a fare sinapsiottimali.con la morte del neurone B hai bisogno di Riarrangiare la sinapsi. 7) Riarrangiamento sinaptico: le sinapsi attive assumono molto probabilmente un fattore neurotrofico che mantiene la sinapsi. Le sinapsi in attiva invece hanno troppi fattori trofici che non le mantengono stabili. 82 8) Mielinazione: dura fino a trent’anni. In questa fascia di età si ha il peso maggiore del cervello. Le malformazioni (teratogeni) possono essere date da agenti fisici, prodotti chimici, microrganismi. Slide 2b: stress e tossico e arricchimento ambientale Lo sviluppo celebrale avviene dal basso verso l’alto E le capacità di cambiamento diminuiscono con l’età.è stato osservato anche che l’interazione dei geni e dell’esperienza porta ad un modellamento dell’architettura del cervello. Le più genetica è quella parte della biologia che studia le interazioni causali fra i geni e loro prodotto, tra geni e ambiente e ci spiega come le interazioni possono modificare le funzioni neurali sia in termini strutturali e di comportamenti. Per modifiche e più genetiche si intende: - Modifiche fenotipiche, - Stabili ed ereditabile, - Geni cellule specifiche, - O in assenza di cambiamenti nel genotipo. I principali meccanismi epigenetici sono: 1) Metilazione: del DNA consiste nel legame di un gruppo metile ad una base azotata permette di gestire l’espressione genica. 2) Modificazioni istoniche: istoni proteine che condensano ordinatamente ed imballano il DNA nei cromosomi. 3) Rimodellamento della cromatina: precede accompagna l’attivazione trascrizionale, cromatina, definita nucleo soma composta da DNA e proteine. I geni nelle prime fasi dello sviluppo guidano il sistema nervoso centrale e la formazione di connessioni e dei circuiti neurali. I fattori ambientali come la dieta, lo stress i farmaci eccetera possono influenzare e modellare la struttura del sistema nervoso centrale durante il suo sviluppo. Il cervello presenta una plasticità neurale che permette di modificarne la struttura e le connessioni cerebrali.la plasticità neurale può avere nature differenti come essere determinata unicamente da attività spontanee geneticamente determinate, determinata da una predisposizione innata a riconoscere rispondere a stimoli specifici esterni oppure influenzata dall’esperienza e dall’apprendimento. La plasticità celebrale sebbene presente in tutta la vita arriva al suo massimo potenziale nei primi anni di vita detto anche “periodo critico” e in queste finestre di vulnerabilità la plasticità celebrale è maggiore ed anche la sensibilità per l’ambiente esterno. I periodi critici possono essere positivi come un arricchimento ambientale, o negativi come quando avviene la deprivazione sensoriale, socio emotiva o è presente un alto tasso di stress o abusi. Ad esempio è stato osservato che è una privazione del sistema visivo porta a drammatico cambiamento della struttura visiva celebrale.si osservato che vi è uno spostamento neuronale dei neuroni dell’occhio deprivato a favore dell’occhio non deprivato questo porta a dei danni come una riduzione 83 dell’acuità una non capacità di visione binoculare. Uno dei siti che maggiormente viene influenzato negativamente dallo stress nella prima infanzia e l’ippocampo. eventi stressanti possono interferire con la sua maturazione strutturale e funzionale anche in modo permanente. Sì osservato che anche una alimentazione poco sana e poco ricca di omega tre possono portare ad un aumento della vulnerabilità allo stress. Un’attivazione dello stress può innescare meccanismi neuro-endocrini I quali con l’attivazione dell’asse ipotalamo-ipofisi-surrene rilasciano un ormone che regola il rilascio della corti contro Pina e dell’ormone dello stress. Diversi studi hanno dimostrato che un’esposizione precoce ad eventi stressanti aumenta la vulnerabilità all’insorgenza di psicopatologie come deficit cognitivi e disturbi dello spettro d’ansia. Lo stress post natale può essere dato da una deprivazione materna, una ripetuta e prolungata separazione della madre e da un basso livello di cura. Gli studi hanno osservato che dopo la separazione dalla madre vi è una diminuzione dell’attività cerebrale soprattutto nella corteccia prefrontale ed un effetto sul sistema endocrino che aumenta la risposta allo stress. in età adulta queste persone tenderanno ad essere più sensibili agli eventi stressanti ad avere delle risposte inferiori a test di apprendimento, di memoria spaziale e di riconoscimento di oggetti. Studi effettuati su bambini cresciuti in orfanotrofi evidenziano che gli è una riduzione del giro orbitale frontale, dell’amigdala, dell’ippocampo, della corteccia temporale laterale, della sostanza bianca della grigia ed un volume ridotto dell’amigdala oltre che una sensibilità maggiore allo stress. Disposizione può aumentare il livello di resilienza rimane comunque legato alla suscettibilità genetica individuale. Arricchimento ambientale: studi sui roditori inseriti in grado di gabbia e condivise in cui vengono posizionati i giochi che permettono ai roditori di svolgere attività motoria cognitiva in maniera volontaria hanno evidenziato un aumento del peso del cervello e dello spessore corticale ed un maggiore grado di plasticità neuronale. Ma anche delle prestazioni migliori e soprattutto su test di apprendimento e di memoria. Questi studi evidenziano ulteriormente la sensibilità dell’ippocampo all’ambiente. Sono state riscontrate anche fattori protettivi nei confronti del declino cognitivo. Intervento di arricchimento ambientale nei confronti del bambino dovrebbe essere tempestivo, personalizzato sui bisogni di ciascun bambino in base alle caratteristiche individuali e all’età e dovrebbe essere multidisciplinare cioè coinvolgere diversi ambiti di sviluppo e basato sulla centralità della famiglia. tali programmi possono essere intrapresi già in epoca neonatale o subito dopo la dimissione ospedaliera. Se alla nascita il bambino presenta delle difficoltà come un peso ridotto possono essere utilizzate anche durante la permanenza l’unità di terapia intensiva chiedendo ai genitori un contatto pelle pelle precoce ed incoraggiando l’allattamento al seno. Altri programmi incoraggiano il coinvolgimento dei genitori insegnando loro delle strategie volte a ridurre lo stress dei fattori esterni. 84 Connetoma: studia la fitta rete delle possibili connessioni neurali. l’obiettivo è quello di mappare i circuiti cerebrali così da poter osservare il contributo di fattori genetici ed ambientali delle differenze individuali. Slide3: il ruolo dell’ambiente e dell’esperienza Durante i primi anni di vita il cervello subisce importanti modifiche le più importanti sono: la crescita di volume, lo sviluppo della Mileena e l’incremento del numero di neuroni e aree celebrale.l’incremento dell’attività sinaptica è fortemente influenzato dall’ambiente dall’esperienza che ne modifica connessioni e ne crea di nuovi in risposta agli input esterni. Nella prima fase di sviluppo si registra una sovrapproduzione sinaptica per poi arrivare a un decadimento sinaptico detto anche potatura in questa fase vengono eliminate molte sinapsi e si arriva a un livello di densità tipicamente presente nell’adulto. Nei primi tre anni il numero di connessioni aumenta verso gli otto e quasi pari a quello di un adulto in un secondo momento ovvero nella pubertà alcune regioni del cervello cominciano a svilupparsi di nuovo per poi tornare nuovamente al numero di connessioni di un cervello adulto intorno ai 18- 20 anni. Alcune abilità vengono a sviluppate dopo la nascita come ad esempio avviene per la vista. Sebbene vi sia un vincolo innato dominio generale che predispone il neonato a prestare attenzione ad alcune caratteristiche strutturali del volto esso verrà influenzato nella sua maturazione dell’ambiente specifico ricco di volti solo così si può sviluppare un sistema dominio specifico deputato alla elaborazione dei volti che permetterà all’adulto in pochi secondi di riconoscere il genere, età espressione emotiva e identità di genere. Quando un vincolo è immerso in un ambiente tipico come quello dei volti si avrà una specializzazione funzionale ovvero la capacità in modo sempre più selettivo di elaborare informazioni specifiche, e una localizzazione strutturale ovvero un restringimento dell’ampiezza della porzione di corteccia che si attiva in risposta a una determinata stimolazione. Ad esempio inizialmente durante la prestazione dei volti si riscontra un’attività della corteccia molto vasta in entrambi gli emisferi, dopo il sesto mese l’attività si localizza sull’emisfero destro permettendo di riconoscere delle caratteristiche configurabili, alla fine del primo anno di vita la localizzazione delle facce corrisponde a un’area più ristretta nella zona temporale destra e in età scolare diventa ancora più localizzata e selettiva corrispondendo all’aria specifica per le facce che si trova nell’adulto. Deficit visivi severi: (PIV) I bambini affetti da BBC sono in grado di rilevare solo luci ed ombre ma non sono in grado di percepire e discriminare le forme la diagnosi di solito viene fatta prima dei sei mesi. Cataratta congenita: I bambini nati con la cataratta congenita bilaterale sei operato intorno ai sei mesi sviluppano la sensibilità al contrasto normale per le basse frequenze spaziali ma manifesteranno un deficit permanente nella sensibilità al contrasto per le frequenze spaziali medie ed alte. 85 Riconoscimento dei volti segue processi percettivi: - Informazioni di primordine: posizione degli elementi che definiscono la geometria del volto. - Informazioni di secondo ordine: relazioni spaziali tra le caratteristiche interne. - Informazioni olistiche unire le singole caratteristiche in un tutto indifferenziato. La discriminazione dei volti avviene sulla base degli elementi locali (base frequenze spaziali) e di elementi configurali (alte frequenze). Nella PIV l’assenza di informazioni visive altre anche lo sviluppo posturale e motorio questo porta questi individui a mettere in atto movimenti stereotipati e ripetitivi. Una buona parte di bambini con PIV sviluppa tratti autistici o problemi nello sviluppo del linguaggio ed utilizza i gesti per condividere e comunicare. Slide 4: Ambiente: l’esperienza e l’ambiente può essere studiata osservando l’apprendimento. Si è infatti osservato che vi era un cambiamento cerebrale come un aumento di volume del cervelletto dell’area di Broca nei musicisti è un maggior volume dell’ippocampo nei guidatori di Taxi. Studiando l’architettura del cervello gli scienziati cercano di studiarne l’attività cognitiva e dunque le reti neurali. La rete neurale più comune è composta da tre strati: 1) Uno strato di unità in input 2) Uno strato di unità nascoste 3) Uno strato di unità in output Ogni rete neurale a una soglia di attivazione ed ogni connessione a uno specifico peso dunque l’attivazione di un’unità di elaborazione avviene solo se la somma dei pesi delle connessioni in input supera la soglia di attivazione dell’unità in questione. Inoltre i modelli connessionistici ci dicono che oltre a input e citatori potrebbero esserci anche input inibitori. Una delle caratteristiche più importanti delle reti neurali e quella di riuscire a modificare i propri pesi in risposta all’output dell’ambiente. Questo è possibile perché la rete riesce ad apprendere e ad estrarre delle regolarità dell’ambiente seguendo la legge che dice che se un evento si manifesta con maggior frequenza sarà più probabile che si ripeta. L’ambiente riesce a modificare il cambiamento grazie a degli stimoli e a dei rinforzi il condizionamento operante è una forma di apprendimento in cui viene fornito il rinforzo solo se il soggetto produce una specifica risposta.ad esempio se un nastro viene attaccato alla caviglia di un bambino farà muovere anche la Juice trina sopra di lui si è osservato che a partire dai due mesi il bambino a prendere la relazione tra un suo specifico comportamento e il rinforzo visivo. Bambini di 1/3 mesi modificano la pressione con la quale succhiano il ciuccio per ottenere un rinforzo visivo.si è osservato che i rinforzi di tipo sociale sono molto efficaci. L’assenza di un rinforzo produce frustrazione. L’apprendimento avviene anche 86 attraverso i rinforzi vicari ovvero attraverso l’osservazione di bambini e di adulti. Ad esempio si sono osservati comportamenti aggressivi soprattutto nei maschi dopo che i bambini sono stati esposti a un contenuto aggressivo. Teoria della mediazione: alcuni comportamenti manifesti possono essere però mediati da risposte non osservabili “mediatori interni” dunque non ci si può basare unicamente sul comportamento osservato ma osservare anche il processo che le genera. Bisogna anche considerare che il livello di comprensione che è un soggetto ha del materiale influenza il modo e la precisione con cui il materiale viene ricordato ma l’apprendimento e la memoria non funzionano nello stesso modo indipendentemente dall’età del soggetto. il livello di sviluppo del soggetto influenza i processi della memorizzazione e le strategie che vengono utilizzate per memorizzare ricordare il materiale. L’uomo potendo manipolare gli input che gli arrivano dall’esterno mostra di essere creativo. I risultati di ricerche recenti suggeriscono che alla nascita il bambino possiede i processi di base necessari per apprendere e generare rappresentazioni del mondo esterno, i bambini sono in grado di formare rappresentazioni anche prima della nascita nell’ambiente intrauterino. inoltre si è osservato che riescono ad avere sofisticate abilità di percezione trasmodare riuscendo ad identificare un oggetto familiare che avevano esplorato nella modalità tattile con la bocca. Non solo dunque hanno capacità di rappresentazione ma sono guidati anche da domini specifici ereditati per via genetica. La permanenza dell’oggetto: durante il periodo sensomotorio piacere per stabilire quando i bambini cominciassero a capire che gli oggetti continuavano ad esistere anche quando non erano percepiti utilizzò una tecnica in cui nascondeva un oggetto con cui il bambino stava giocando. utilizzò l’assenza di ricerca del giocattolo da parte del bambino come dimostrazione della mancanza di capacità rappresentativa. Il bambino solo nel quarto stadio a otto-12 mesi manifesta di ricercare l’oggetto anche quando è completamente nascosto ma tenderà a cercare l’oggetto nel luogo 87 dove lo ha trovato la prima volta anche se il nascondiglio è stato spostato davanti suoi occhi. I bambini dunque possiedono una rappresentazione procedurale di ciò che devono fare quando un giocattolo scompare non una vera e propria rappresentazione mentale dell’oggetto. Per arrivare ai concetti e a come vengono individuati dalle percezioni noi tendiamo a raggruppare gli oggetti sulla base di proprietà percettive e funzionali simili così da poter estrarre delle regolarità semantiche come tutti i triangoli hanno tre lati. I concetti riducono la complessità e sono più maneggevoli inoltre hanno anche aspetti sociali come il consenso culturale la condivisione e lo scambio tra individui. E permette all’uomo di utilizzare un linguaggio comune. Slide 5: metodi di indagine e di neuro modulazione in età evolutiva. Il comportamento e le attività del sistema nervoso vengono studiate in condizioni controllate. Il comportamento viene misurato attraverso indici particolari come il tempo, prestazioni corrette e sbagliate, o accuratezza sul tempo ovvero l’efficienza. Ci sono diversi modi per studiare il comportamento in maniera controllata come ad esempio studi su esseri umani e animali a cui vengono assegnati dei compiti. Possono essere utilizzati anche dei modi di neurofisiologia comportamentale che attraverso la registrazione di un singolo elettrodo registrano l’attività celebrale durante l’esecuzione di compiti dati oppure vanno a stimolare determinati neuroni. Ci possono essere anche registrazioni di unità multiple come ECoG ( simile all’EEG) ma gli elettrodi sono posizionati direttamente nel cervello. Una fonte molto utile sono gli studi sulle lesioni che possono essere sia indotte sia di origine spontanea osservando le lesioni si possono confrontare le prestazioni con soggetti senza lesione oppure con soggetti che presentano lesioni in altre aree. Esistono poi dei mezzi che permettono lo studio delle immagini del cervello in vivo come ad esempio la tac la risonanza magnetica la PET la risonanza magnetica funzionale l’elettroencefalografia eccetera. TAC: permette di ricostruire l’immagine tridimensionalmente al contrario delle normali radiografie. È uno scanner a raggi X che ruota più di 180° . Un fascio di raggi 88 X viene proiettato attraverso la testa del paziente e l’immagine che si ottiene fornisce una misura della densità dei tessuti questo perché il materiale a bassa intensità come l’aria o liquidò assorbirà una quantità modesta di raggi X mentre l’osso ne assorbirà una quantità notevole. Sebbene la TAC sia lo strumento molto importante la sua risoluzione non permette analisi approfondite inoltre è molto difficile distinguere tra sostanza bianca e sostanza grigia. MRI risonanza magnetica: sfrutta le proprietà magnetiche degli atomi di idrogeno.normalmente l’orientamento dei protoni è casuale e non è influenzato dal campo magnetico generato dalla gravità terrestre. La risonanza magnetica fa assumere un orientamento parallelo alla forza del campo magnetico dell’apparecchio ai protoni. Poi vengono inviate onde radio che turbano l’orientamento dei protoni producendo energia elettromagnetica che viene rilasciata anche dopo che l’impulso viene spento. i diversi tessuti producono energia e differenti in questo modo il computer riesce a ricostruire le immagini. I vantaggi sono che non vi è esposizione a radiazioni e sia una risoluzione spaziale migliore infatti si riescono a distinguere la materia bianca e quella grigia. di contro è un apparecchio molto costoso che non può essere utilizzato in presenza di nessun metallo e risulta essere molto rumoroso. DTI Tensore di diffusione: È una variante della risonanza magnetica che misura la densità e il movimento dell’acqua contenuta all’interno degli assoni. Poiché la distribuzione dell’acqua e controllata dalla membrana mielinica degli assoni in quanto è lipidica questo permette di analizzare la diffusione e la direzione del flusso delle molecole d’acqua all’interno dei tessuti riuscendo a costruire una mappatura tridimensionale delle fibre di sostanza bianca. PET tomografia a emissione di positroni: riesce a rilevare le variazioni del metabolismo che si verificano durante l’esecuzione di un compito cognitivo. Poiché i neuroni richiedono ossigeno e glucosio così da poter avere l’energia necessaria per l’esecuzione del compito. Quando un’area celebrale e attiva riceve una quantità maggiore di ossigeno e di glucosio attraverso un aumento del flusso sanguigno a scapito di altre parti del cervello. La PET misura le variazioni del flusso sanguigno locale. per fare ciò viene inserita nel sangue una sostanza radioattiva. Isotopi contenuti nella sostanza radioattiva tendono a decadere rapidamente e mettendo un positrone. quando un positrone si scontra con un elettrone si genera un n’erano due fotoni o raggi gamma. i due fotoni si muovono nella direzione opposta l’una rispetto all’altro attraversando il tessuto del cervello la PET riesce a tracciare i raggi gamma determinando il punto in cui è avvenuta la collisione. l’immagine mostrerà la distribuzione del flusso sanguigno. 89 FRI funzionale (FMRI) risonanza magnetica funzionale: studia il consumo di ossigeno in quanto vi è un aumento del flusso sanguigno nelle regioni dove si verifica maggiore attività neurale. L’immagine viene costruita sulle proprietà magnetiche dell’emoglobina la cellula che trasporta ossigeno. L’emoglobina risulta di amagnetica quando ossigenata e para magnetica quando non è ossigenata. Sebbene ci si aspetterebbe una diminuzione delle aree attive di emoglobina ossigenata in realtà poiché ne viene mandata tanta il tessuto non è in grado di assorbire tutto e dunque si registrano contrariamente a quello che si pensa un aumento di ossigenazione. Psicofisiologia: - Elettroencefalogramma (EEG): permette di misurare il campo elettrico dei neuroni in modo non invasivo attraverso dei sensori che vengono posizionati sulla cute. I sensori possono essere 200 o 256 e vengono montati su una cuffia elastica. sono più grandi di quelli utilizzati per la registrazione di singoli neuroni. fornisce una registrazione continua dell’attività globale del cervello. Attività diverse portano a tracciati differenti ad esempio mentre dormiamo profondamente le oscillazioni sono lente e molto ampie. Permettono di modulare anche l’attività cerebrale durante un compito. le risposte vengono detti i potenziali correlati a eventi (ERP). - Magnetoencefalografia (MEG): simile agli ERP in quanto l’attività elettrica in quanto l’attività elettrica insieme all’attività sinaptica produce dei piccoli campi elettromagnetici.fornisce anche la stessa risoluzione ma è più precisa a localizzare la fonte del segnale in quanto i segnali magnetici non sono influenzati dalla struttura ossea o liquida del cranio e del cuoio capelluto. - Elettrocortigramma (ECoC): simile all’EEG ma gli elettrodi sono collegati direttamente sulla superficie del cervello al di fuori della dura madre e al di sotto di essa.viene utilizzato solo per persone che devono essere sottoposte ad un intervento chirurgico dove l’équipe deve monitorare l’attività celebrale durante l’operazione. Stimolazione celebrale. TMS stimolazione magnetica trans cranica: È un metodo non invasivo per simulare in maniera focale il cervello umano. È formata da una bobina rivestita da materiale isolante e collegata a potenti condensatori elettrici. Una volta che la corrente arriva la bobina si forma un campo magnetico. La bobina viene posata sulla superficie del cranio così il campo magnetico riesce a superare la cute e lo scalpo portando una corrente fisiologica che fa generare impulsi ai neuroni. Il flusso di corrente provoca una depolarizzazione. la stimolazione della corteccia ad esempio quella motoria permette di valutare l’integrità delle vie motorie perché attiva la muscolatura dopo la 90 stimolazione.ma può anche condurre a lesioni virtuali stimolando il cervello in aree particolari alterandone l’attività normale. TDCS stimolazione transcranica in corrente continua: viene inviata una corrente tra due piccoli elettrodi un anodo e un catodo che sono appoggiati sul cuoio capelluto.i neuroni al di sotto del anodo divengono dei polarizzati.in questo modo sono più eccitabili e più probabile che avvenga l’avvio del potenziale d’azione, al contrario i neuroni sotto il catodo diventano iper polarizzanti e dunque una minore probabilità di scarica. L’effetto della stimolazione può durare anche dopo la fine della stimolazione infatti più lunga sarà la stimolazione più lento sarà il ritorno all’origine. Questo metodo viene utilizzato anche per condizioni patologiche come depressione, riabilitazione cognitiva e dolore cronico. Le tecniche sopradescritte sono utilizzate perlopiù con gli adulti nel caso di indagini in età pediatrica dobbiamo considerare dei mezzi che siano tollerabili e sicuri. EEG: risulta sicura sui neonati bene è posizionata una cuffia con più o meno 128 canali. la registrazione nei bambini è buona perché la cuffietta risulta morbida e gli elettrodi sono posizionati al suo interno e ammorbiditi con morbide spugne imbevute di semplice soluzione salina.si presenta veloce da applicare e non porta ad abrasioni. La copertura del canale ad alta densità consente la massima risoluzione spaziale. MRI/FMRI: sebbene presenti una risoluzione spaziale migliore presenta dei limiti poiché i bambini vanno spesso sedati e le analisi statistiche del segnale fanno riferimento a valori adulti. Esistono comunque dei macchinari colorati per bambini così da rendere l’ambiente meno fastidioso. FNIRS Nirs funzionale: È una tecnica di immagine di ricerca specializzata che utilizza la luce infrarossa per esaminare la funzione del cervello vivente. è un metodo sicuro ed economico che monitor ai cambiamenti emodinamici. TDCS stimolazione celebrale: È abbastanza sicura non essendo invasiva anche se è molto costosa.quelle a rilascio continuo sono più pericolose in quanto contengono degli impianti chirurgici nel cervello. Recenti studi però hanno dimostrato che vi è un’alta tollerabilità in età evolutiva riducendo il rischio.questo metodo risulta buono come risposta ad alcune malattie come l’autismo il disordine del linguaggio e l’ADHD ma anche nei disturbi del comportamento alimentare e altri disturbi psichiatrici. Qualora i metodi di farmaci ecologici non funzionassero si può rivelare un ottimo strumento. Neurofeedback: proponendo al soggetto il proprio parametro EEG attraverso strategie mentali e corporee si riesce a controllare tale funzionamento facendolo rientrare entro dei limiti significativi sono stati osservati dei risultati 91 significativi per bambini con ADHD, spettro autistico, epilessia, problemi del sonno, recupero del trauma eccetera. L’EEG misura i potenziali post sinaptici a livello delle cellule piramidali che si trovano sulla superficie dunque non può misurare l’attività elettrica sotto corticale. Slide 6 sviluppo delle funzioni sensoriali: funzioni sensoriali nel feto: il periodo prenatale è un periodo in cui si sviluppano diverse funzioni sensoriali come ad esempio il tatto. A 8/9 settimane di gestazione si vengono toccate le labbra e le guance il feto sposta la testa dallo stimolo. Nel secondo trimestre non si allontana più dallo stimolo ma si muove verso di esso. A 14 settimane la maggior parte del corpo risulta sensibile al contatto e anche la settimana in cui il feto inizia a succhiare il dito e si tocca il viso. Udito: vi è una risposta agli stimoli sonori già a partire dal terzo trimestre di gravidanza. A 27 settimane è possibile registrare risposte evocate da stimoli acustici con la magnetoencefalografia. La risposta del feto è influenzata dalla frequenza e dall’intensità del suono. Le risposte inizialmente sono limitate ai suoni di bassa frequenza da 200 500 Herz. Gusto e olfato: generalmente si riscontra che il feto ingoia il liquidò amniotico già a partire dalla 12ª settimana.se nel liquidò viene aggiunto dello zucchero il feto né inghiottirà di più al contrario si verrà aggiunto un gusto spiacevole nei inghiottirà di meno. I feti tendono ad orientarsi verso il loro liquidò amniotico dimostrando una continuità sensoriale che dura fino ai primi quattro giorni di vita.i neonati preferiscono l’odore dell’oro liquidò piuttosto che quello del latte artificiale. Visione: esponendo il feto ad un Flesh molto brillante si registrano cambiamenti a livello di frequenza cardiaca. a 26 settimane potenziali evocati nei prematuri. Ma considerando l’ambiente intrauterino il sistema visivo non è stimolato per questo motivo la visione è il senso che inizia un avere propria interazione con l’ambiente e il momento della nascita. Nel momento della nascita le capacità visive del bambino sono molto diverse. Le vie visive si sono già formate e la corteccia visiva riceve i segnali della retina anche il nucleo genetico lato laterale del talamo ha già l’aspetto tipico di quell’adulto e sono presenti anche le colonne di dominanza oculare a livello del quarto e del sesto strato. Rimangono però diverse la densità neuronale, la morfologia delle cellule e l’organizzazione dei campi recettivi nelle diverse vie. Ad esempio nella corteccia visiva primaria gli strati più profondi hanno una maggiore complessità e lunghezza dell’albero dentro litico rispetto agli stati superficiali. 92 La capacità di discriminare i dettagli dello stimolo visivo (acuità) e 40 volte più bassa alla nascita rispetto a un adulto. L’acuità visiva si può stimare o con tabelle ottiche oppure utilizzando i reticoli che sono degli stimoli visivi costituiti da righe chiare ed a righe scure E viene valutata per mezzo di potenziali evocati visivi. Nei neonati si valuta con tecniche comportamentali come la fissazione preferenziale o con tecniche elettrofisiologiche potenziali evocati visivi. L’acuità visiva si sviluppa molto velocemente nei primi mesi di vita nei sei mesi risulta essere otto volte più bassa anche nell’adulto. dopo il sesto mese però la velocità diminuisce e i valori dell’adulto si raggiungono solo verso i sei anni di età. La sensibilità al contrasto e basta alla nascita per misurarla vengono utilizzati dei reticoli in cui si riduce il contrasto finché il bambino non è più in grado di vederlo. La sensibilità al contrasto migliora con l’età ma non raggiunge valori dell’adulto fino ai sei anni per alcuni autori fino ai 15 anni. Anatomicamente la fovea è responsabile della qualità visiva nel neonato risulta immatura. nell’adulto i coni raggiungono i 160.000 per millimetro quadrato e risultano anche molto vicini gli uni agli altri ordinati in file ordinate. nei neonati non solo si registra una densità quattro volte più bassa ma risultano posizionati in maniera più larga. a 45 mesi risultano essere la metà rispetto a quelli di un adulto. Non solo risultano un numero inferiore ma anche le caratteristiche strutturali dei neuroni del nucleo genetico lato laterale della corteccia visiva primaria risultano in maturi. alla nascita le dimensioni dei neuroni sono molto più piccole e continuano ad aumentare fino ai due anni nel nucleo genetico lato laterale e fino a 8/12 mesi in V1. Lo sviluppo però tende poi a diminuire e raggiunge circa 11 anni quello dell’adulto. Avviene la cosiddetta potatura che permette una riorganizzazione degli ingressi sinaptici sui neuroni corticali. La capacità di visione binoculare è assente alla nascita. Il bambino tende a fissare più allungo uno stimolo tridimensionale che uno stimolo piatto. L’hai vista binoculare comincia a manifestarsi a quattro mesi di età e risulta simile a quella di un adulto intorno ai sette anni. A quattro mesi infatti vi è un rapido sviluppo dei movimenti oculari. L’accomodamento ovvero la capacità di mettere a fuoco oggetti a distanza è presente alla nascita ma solo una distanza di circa 30 cm e tipica della distanza del viso della madre quando allatta. Intorno alle 10-13 settimane il bambino in grado di mettere a fuoco accuratamente sia oggetti vicini che lontani. La visione dei colori dipende dai coni e dalle cellule P. La capacità di discriminare i colori e documentata a partire dai due mesi di vita e non è revocabile prima delle cinque settimane. La sensibilità al contrasto cromatico matura in fretta già pochi mesi è come quella dell’adulto. Il riconoscimento degli oggetti appare già in bambini di due giorni. 93 Slide 7: importante per lo sviluppo dell’integrazione dei contorni e lo sviluppo delle connessioni intracorticali orizzontali presenti nella corteccia visiva primaria. Correndo in parallelo alla superficie corticale connettono colonne diverse e consentono di integrare l’informazione su zone di campo visivo molto più grandi delle dimensioni dei campi recettivi delle singole cellule corticali.il risultato è che è una cellula può essere influenzata da stimoli al di fuori del suo campo ricettivo e può quindi scambiare la sua risposta in funzione del contesto in cui lo stimolo visivo appare. Se non vi fosse questa connessione orizzontale ad esempio un bordo di uno specchio non apparirebbe continuo ma composto da tanti frammenti verticali. Tali connessioni però maturano in maniera più lenta. Le connessioni verticali che collegano strati diversi come lo strato VI e IV si sviluppano intorno alla 26ª e 28ª settimana di gestazione. Le connessioni orizzontali invece sviluppano più lentamente e sembrano apparire solo intorno alla 37ª settimana di gestazione nello strato IV b e V. Dopo la nascita le fibre aumentano rapidamente fino a che a sette settimane non è evidente un plesso uniforme di connessioni. Il fatto che le connessioni verticali appaiono prima di quelle orizzontali potrebbe essere dato che le caratteristiche locali sviluppano prima di quelle necessarie per integrare le caratteristiche e dare un’idea di continuità e coerenza all’immagine. Il fatto che vi siano delle connessioni già dopo i quattro mesi di vita e che si siano mature al 15º mese nello strato IVB fa pensare che l’elaborazione al movimento maturi prima rispetto alla forma. Illusione della forma è stata valutata utilizzando l’illusione dei pallini circondati da altri pallini in bambini di quattro anni e negli adulti si è mostrato che l’illusione era minore nei bambini rispetto agli adulti dimostrando che il contesto è meno importante. In altri esperimenti sono stati confrontati bambini di tre e di 11 anni i primi dimostravano una minore suscettibilità all’inclusione degli adulti ma già 5-6 anni la grandezza dell’effetto non differiva più dalla risposta adulta. Le connessioni orizzontali sono molto sensibili all’esperienza visiva per questo motivo sono vulnerabili ad alterazioni dell’esperienza e possono portare a un’anomalia visiva. I soggetti ambliopi hanno una difficoltà a percepire i contorni. tale difficoltà è stata osservata anche in pazienti Strabici che avevano una acuità normale ad entrambi gli occhi ma una stereopsi ridotta o assente. Questo fa pensare che la capacità di distinguere i contorni dipende anche dalla presenza di interazioni binoculari normali durante lo sviluppo. Orientamento visivo del bambino: inutilmente oculari che spostano la direzione dello sguardo sono detti overt Mentre gli spostamenti dell’attenzione visiva tra zone del campo visivo o da un oggetto all’altro che avvengono indipendentemente dei miei vuoi menti della testa o degli occhi sono chiamati covert. Nei primi mesi l’orientamento 94 over cambia notevolmente.vi è una via diretta della retina al collicolo superiore coinvolta nel rapido inizio dei movimenti oculari verso uno stimolo semplice che riceve principalmente dal campo visivo temporale retina e tre vie corticali. La seconda via invece cortico-conllicolare ha origine in V1 e da MT guidata dalla via magno cellulare. La terza via cortico-corticale riceve in ingresso sia della via M Ket dalla via si è coinvolge le aree frontali. Questa via è coinvolta nel controllare le sequenza temporale dei movimenti oculari all’interno di un’immagine complessa che permette un’analisi dettagliata degli stimoli visivi. 95