caricato da
common.user5889
Anatomia III: Tronco cerebrale e cervelletto - Appunti

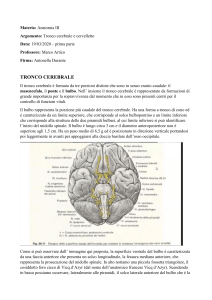
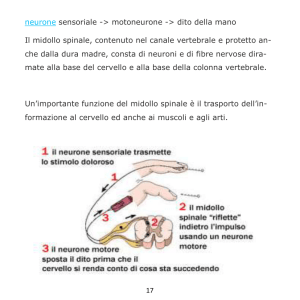
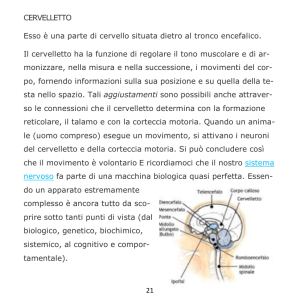
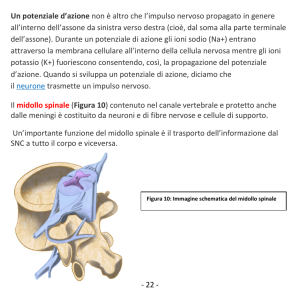
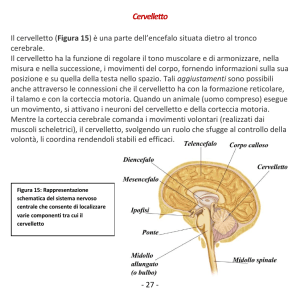
Materia: Anatomia III Argomento: Tronco cerebrale e cervelletto Data: 19/03/2020 – prima parte Professore: Marco Artico Firma: Antonella Durante TRONCO CEREBRALE Il tronco cerebrale è formato da tre porzioni distinte che sono in senso cranio-caudale: il mesencefalo, il ponte e il bulbo. Nell’ insieme il tronco cerebrale è rappresentato da formazioni di grande importanza per la sopravvivenza dal momento che in esso sono presenti centri per il controllo di funzioni vitali. Il bulbo rappresenta la porzione più caudale del tronco cerebrale. Ha una forma a tronco di cono ed è caratterizzato da un limite superiore, che corrisponde al solco bulbopontino e un limite inferiore che corrisponde alla struttura delle due piramidi bulbari, al cui limite inferiore si può identificare l’inizio del midollo spinale. Il bulbo è lungo circa 3 cm e il diametro anteroposteriore non è superiore agli 1,5 cm. Ha un peso medio di 6,5 g ed è posizionato in direzione verticale portandosi poi leggermente in avanti per appoggiarsi alla doccia basilare dell’osso occipitale. Come si può osservare dall’ immagine qui proposta, la superficie ventrale del bulbo è caratterizzata da una faccia anteriore che presenta un solco longitudinale, la fessura mediana anteriore, che rappresenta la prosecuzione del midollo spinale. In alto notiamo una piccola fossetta triangolare, il cosiddetto foro cieco di Vicq d’Azyr (dal nome dell’anatomico francese Vicq d’Azyr). Scendendo in basso possiamo osservare, lateralmente alle piramidi, il solco laterale anteriore del bulbo che è la continuazione dell’omonimo solco del midollo a livello del quale emergono le radici anteriori dei nervi spinali. I rilievi che si riconoscono sulla superficie anteriore del bulbo sono principalmente le due piramidi che sono piuttosto evidenti e voluminose e che lateralmente presentano le olive bulbari. Posteriormente il bulbo presenta in basso un solco mediano posteriore. La porzione inferiore del bulbo assomiglia al midollo spinale e presenta gli stessi solchi. Superiormente invece la forma del bulbo è condizionata dalla presenza del quarto ventricolo. In alto, a livello bulbare, si riconosce la presenza di cordoni di sostanza bianca che corrispondono al fascicolo cuneato e al fascicolo gracile, che sono la prosecuzione degli omonimi fascicoli che si riscontrano a livello del midollo spinale. Le olive bulbari, invece, occupano la parte anteriore del cordone laterale. Internamente il bulbo ha caratteristiche diverse rispetto al midollo spinale. Una sezione praticata all’ altezza della zona di passaggio tra midollo spinale e bulbo, permette di identificare la fessura mediana anteriore e anche il solco mediano posteriore e in particolare la sostanza grigia ha una forma che ricorda abbastanza quella del midollo spinale. Se invece pratichiamo una sezione a livello superiore del bulbo, la disposizione non è quella sin ora descritta, infatti si può osservare sempre una simmetria ma non c’è più una separazione con la fessura mediana anteriore mentre il solco mediano posteriore è scomparso; il canale centrale si allarga perché si va ad aprire nel quarto ventricolo e le metà laterali sono formate da sostanza bianca e sostanza grigia ma con rapporti modificati. Infatti la disposizione cambia proprio in relazione alla disposizione, all’ interno del bulbo, del quarto ventricolo. Nell’ immagine riportata in alto si riconosce in basso il bulbo con le due piramidi bulbari in primo piano e le olive bulbari appena al lato di esse. Il tronco cerebrale visto ventralmente è caratterizzato dalla emergenza dei nervi cranici. Questi emergono tutti a livello ventrale tranne uno, il nervo trocleare (IV nervo cranico) che emerge dorsalmente. Nella precedente immagine sono inoltre facilmente riconoscibili i tre segmenti: il segmento superiore del tronco cerebrale, vale a dire il mesencefalo, il segmento intermedio, il ponte di Variolo, detto anche protuberanza anulare per la sua voluminosa sporgenza e infine, in basso, il bulbo. È opportuno osservare come nel bulbo siano presenti le origini apparenti, ossia le emergenze di numerosi nervi cranici che già dal limite fra ponte e bulbo si cominciano ad evidenziare e si portano in senso mediolaterale per allontanarsi dal tronco cerebrale stesso. Si riconoscono molto bene le piramidi bulbari e al lato le olive bulbari. In questa immagine è possibile osservare la differenza tra midollo spinale e il bulbo, nel quale si riconoscono le piramidi; è poi visibile il pavimento del quarto ventricolo relativamente al triangolo bulbare e si nota la disposizione dei fasci che si collocano, assieme anche ai nuclei dei nervi cranici, nella regione immediatamente sottostante al pavimento del quarto ventricolo. In questa immagine possiamo vedere delle sezioni praticate a vari livelli. A seconda se ci troviamo nel bulbo (in basso) nel ponte (nel mezzo) e a livello mesencefalico (in alto), notiamo la collocazione dei nuclei dei nervi cranici che sono posizionati all’ interno del tronco cerebrale e a livello mesencefalico. È visibile essenzialmente il nucleo del nervo oculomotore e il nucleo del trocleare, con una piccola propaggine del nucleo del trigemino che si prolunga appena verso il mesencefalo. In realtà il trigemino ha un nucleo molto lungo e voluminoso che è contenuto all’ interno del ponte e assieme ad esso si trovano, sempre a livello pontino, i nuclei del VI nervo cranico (il nervo cranico con il decorso intracisternale più lungo, cosa che lo rende particolarmente suscettibile nei casi di ipertensione endocranica) e poi troviamo anche il nucleo del V, VII e VIII nervo cranico. Poi scendendo più in basso troviamo i nuclei dei nervi contenuti nel segmento più caudale del tronco cerebrale, identificando cosi il IX, il X e il XII nervo cranico. L’XI, l’accessorio spinale, ha un nucleo in parte anche contenuto nel midollo spinale. In questa immagine si apprezza molto bene la conformazione del quarto ventricolo. È possibile notare anteriormente la voluminosa prominenza del ponte, mentre più in basso notiamo la presenza del bulbo. Il quarto ventricolo si connette in alto attraverso un condottino, l’acquedotto mesencefalico di Silvio, con il terzo ventricolo. In questa foto è possibile apprezzare il mesencefalo e l’acquedotto cerebrale di Silvio, localizzato nel pieno del mesencefalo. Più in basso rispetto all’ acquedotto cerebrale si nota il nucleo rosso e la sostanza nera di Sommering, due dei più importanti nuclei del mesencefalo. Più in alto è possibile osservare la porzione più craniale del mesencefalo, la lamina quadrigemina formata dai quattro tubercoli quadrigemelli, due superiori e due inferiori. Più in basso si osservano il piede del peduncolo cerebrale con la fossa interpeduncolare a livello della quale emerge il terzo nervo cranico, l’oculomotore comune. Visto dorsalmente, il tronco cerebrale permette di osservare la presenza di quattro tubercoli quadrigemini e la lamina quadrigemina. I tubercoli quadrigemelli, chiamati anche collicoli, sono due superiori e due inferiori. Quelli superiori sono collegati, come visibile nell’ immagine schematica, alle vie ottiche, mentre quelli inferiori sono collegati alle vie acustiche. Facendo riferimento a sinistra dell’immagine precedente, è possibile notare in basso il quarto ventricolo, dove è stato sezionato l’insieme dei peduncoli cerebellari, permettendo di riconoscere la presenza dei peduncoli cerebellari superiori, medi e inferiori che sono responsabili del collegamento rispettivamente con il mesencefalo, per quanto riguarda il peduncolo superiore, il ponte, grazie al peduncolo cerebellare medio e il bulbo grazie all’ inferiore. Tra i sei peduncoli cerebellari, tre sul lato destro e tre sul lato sinistro, si nota la presenza della fossa romboidale o pavimento del quarto ventricolo. Notiamo anche la presenza delle barbe del calamo scriptorio, con una struttura simile a quella di un pennino per scrivere. Nell’immagine posta in alto, è possibile osservare diverse sezioni del tronco cerebrale praticate a vari livelli. Essa è utile, inoltre, per vedere in che modo decorrono le fibre della via della spinotalamica (indicata sulla sinistra). L’informazione viene raccolta dal neurone situato nel ganglio spinale, che ha un prolungamento verso la periferia e un altro che rientra nel corno posteriore. Dal corno posteriore passa controlaterlamente e ascende con i fasci spino-talamici verso l’alto, attraversando quindi il bulbo, il ponte, il mesencefalo fino ad arrivare al talamo dove proietta nella corteccia parietale. La via spino-talamica è importante per la trasmissione della sensibilità termodolorifica. Facendo riferimento a destra della precedente immagine, invece, viene riportata la via relativa della sensibilità propriocettiva che attraverso i fascicoli del cordone posteriore, fascicolo gracile di Goll e il fascicolo cuneato di Burdach, risale verso l’alto portandosi a livello della callotta mesencefalica e finalmente verso il talamo per la proiezione in corteccia. Possiamo quindi distinguere un sistema antero-laterale, quello del fascio spino-talamico anteriore e fascio spino-talamico laterale, responsabili il primo della sensibilità tattile protopatica e il secondo della sensibilità termo-dolorifica. Tutte queste informazioni arrivano al nucleo ventroposterolaterale del talamo dal quale poi l’informazione viene proiettata alla corteccia per la discriminazione finale. La sensibilità tattile epicritica e la sensibilità propriocettiva cosciente percorrono invece una strada diversa: funzionano sempre come colonne posteriori ma utilizzano il lemnisco mediale. Quindi c’è un primo neurone, un neurone spino-bulbare che dal midollo spinale si porta al bulbo e poi un secondo neurone che attraverso i nuclei gracile e cuneato permette di raggiungere sempre il talamo dal quale finalmente, con il terzo neurone talamo-corticale, l’informazione verrà inviata alla corteccia sensitiva. In questa immagine è visibile il mesencefalo in primo piano. Il nucleo rosso, che è particolarmente importante nel sistema extrapiramidale del quale costituisce una stazione fondamentale, è contraddistinto da una vaga colorazione rossastra e si distingue in una componente parvicellulare e una componente magnicellulare. Tra i nuclei più importanti del mesencefalo, oltre i nuclei propri dei nervi cranici, troviamo il nucleo interstiziale di Cajal e il nucleo interpeduncolare, ma sicuramente il nucleo di maggiore interesse dal punto di vista della neurotrasmissione del sistema extrapiramidale è la sostanza nera di Sommering. La sostanza nera è un nucleo allungato che si estende al limite tra la calotta, situata cranialmente, e il piede del peduncolo cerebrale situato più caudalmente e lateralmente; si tratta di un nucleo molto importante perché è deputato alla produzione di dopamina. Questo è il neurotrasmettitore più importante del sistema extrapiramidale e nel caso in cui la sostanza nera dovesse essere danneggiata o sofferente, come si verifica a causa di problemi vascolari o di traumi ripetuti, come accade nei pugili, o anche a causa di sostanze di natura farmacologica o per la presenza di alcuni insetticidi nell’ ambiente come esteri fosforici, parathion e malathion che sono particolarmente tossici per i neuroni della sostanza nera, tutto ciò può portare ad una condizione di sofferenza della sostanza nera che si traduce nella comparsa di una sindrome nota come morbo di Parkinson. Questa patologia è caratterizzata da una funzionalità completamente alterata della sostanza nera, con insufficiente produzione di dopamina. Ciò rende i movimenti estremamente difficoltosi dal momento che si viene a creare una condizione di sofferenza del sistema extrapiramidale che non è più in grado di assecondare l’esecuzione dei movimenti nella loro completa armonia e fluidità. (il sistema extrapiramidale coordina l’attività motoria involontaria automatica e associata). CERVELLETTO In questa immagine è possibile identificare il cervelletto in sezione longitudinale secondo un piano sagittale. Notiamo anche la presenza del quarto ventricolo e anteriormente ad esso troviamo il ponte e il bulbo. Il cervelletto presenta all’ interno il cosiddetto “arbor vitae”, la tipica arborizzazione della sostanza bianca che costituisce la porzione profonda di questo organo, mentre esternamente si può notare la presenza della corteccia cerebellare che è perfettamente identica, in ogni punto, al cervelletto, a differenza dell’encefalo in cui la corteccia è diversa a seconda della regione presa in considerazione. In questa sezione obliqua del cervelletto è possibile notare i due emisferi cerebellari, insieme alla sostanza bianca profonda dell’emisfero cerebellare destro e sinistro, e in mezzo tra i due vi è una porzione centrale chiamata verme. In questa immagine è stato invece lasciato in sede l’encefalo con i due emisferi. Si nota la scissura interemisferica e più in basso si osserva il cervelletto. La dura madre è stata aperta; in basso è conservato il segmento cervicale del midollo spinale con le radicole che fuoriescono dai solchi posterolaterali delle radici spinali. Questo disegno schematico permette di apprezzare le diverse porzioni del cervelletto. Notiamo la presenza dei due emisferi lateralmente e del verme al centro. Il verme presenta alcune formazioni come la lingula, il culmen, il declive, il folium, il tuber, il monticulus. I solchi e le scissure permettono di suddividere prima in lobuli e poi in lamelle la struttura del cervelletto. Morfologicamente e filogeneticamente è possibile distinguere nel cervelletto 3 circuiti. Il primo circuito, quello più antico dal punto di vista filogenetico, corrisponde al cosiddetto lobulo flocculo nodulare che comprende le formazioni dei flocculi, che si trovano lateralmente e che sono collegate al nodulo (fare riferimento alla precedente foto). Questo circuito archicerebellare è collegato ai nuclei trigeminali e al sistema vestibolare ed ha una importanza notevole nel regolare l’equilibrio e quindi è particolarmente sviluppato nei volati e nelle scimmie arboricole, ovvero laddove sia necessario avere un controllo perfetto dei movimenti durante il volo o durante il passaggio da un ramo a un altro. Il secondo circuito, il cosiddetto circuito paleocerebellare, che è meno antico dell’archicerebellum ma è più antico del neocerebellum, è caratterizzato da una serie di vie afferenti che arrivano dal midollo spinale, come afferenze esterocettive e propriocettive, e in parte anche dal sistema vestibolare, ed è il circuito di maggiore rilievo per il controllo del tono muscolare e della postura. Questo circuito comprende alcune porzioni vermiane, ovvero il lobulo centrale, il culmen, la piramide e l’uvola, ma presenta anche una parte degli emisferi, vale a dire il lobulo centrale, il quadrangolare e i paraflocculi. Esso fa capo a due nuclei: il nucleo globoso e il nucleo emboliforme che si trovano nella profondità della regione centrale del cervelletto. Al contrario, il lobulo flocculo nodulare del circuito archicerebellare fa capo al nucleo del fastigio o nucleo del tetto. Infine, il terzo circuito, cioè il circuito neocerebellare, filogeneticamente più recente, è massimamente sviluppato nei mammiferi e riceve afferenze corticali tramite i nuclei basilari del ponte e le olive inferiori, ma anche afferenze esterocettive, visive e acustiche e fa capo a livello del verme, al declive e al tuber, mentre per quanto riguarda gli emisferi è collegato ai lobuli semilunari superiori e inferiori, al lobulo digastrico e alle tonsille. Il nucleo di riferimento nella profondità del cervelletto è il nucleo dentato. La funzione del neocerebellum è quella di integrare le afferenze visive, acustiche ed extrapiramidali in modo tale da garantire l’esecuzione di movimenti complessi e di determinare il coordinamento dell’attività motoria degli arti superiori e inferiori, come di fatto si rende necessario durante attività motorie complesse come la camminata, la corsa, il salto e l’atterraggio. In questa immagine è stato realizzato un taglio trasversale del cervelletto, rendendo evidente la distribuzione della sostanza grigia e della sostanza bianca. Si nota la presenza del verme in posizione centrale in tutte e tre le sezioni. Divergendo in senso mediolaterale si notano i due emisferi cerebellari. La corteccia del cervelletto è caratterizzata da 3 strati cellulari: -quello più esterno è lo strato molecolare -in mezzo c’è lo strato delle cellule gangliari o strato delle cellule di Purkinje (N.B: in lingua boema la “j” non si pronuncia, quindi la terminologia corretta è cellule di “Purkine”) -a cui fa seguito in profondità lo strato dei granuli, caratterizzato da neuroni che sono in grado di esaurire a breve distanza dal pirenoforo i dendriti che li caratterizzano. Attraverso questi collegamenti a più livelli, sia in senso verticale che in senso orizzontale, il cervelletto è in grado di integrare in maniera complessa informazioni che provengono dai diversi circuiti che lo caratterizzano, facendo in modo di collegare poi queste informazioni da ciascuno strato, in orizzontale prima e in verticale poi, per raggiungere i nuclei profondi del cervelletto stesso. Da notare in alto nell’ultima immagine, in rosso, una cellula gangliare di Purkinje con un grosso pirenoforo di forma a fiasco, la quale presenta una spiccata ed estesa arborizzazione dendritica che si allarga in tutto lo strato soprastante, cioè lo strato molecolare, mentre l’assone scende in profondità per andare verso i nuclei centrali del cervelletto stesso. Materia: Anatomia III Argomento: Cervelletto Data: 19/03/2020 - seconda parte Prof.: Marco Artico Firma: Roberta Dortucci Nel disegno schematico qui proposto si nota meglio l’esempio appena fatto: si individua l’arborizzazione dendritica della cellula del Purkinje, con l’assone che scende in profondità all’interno del cervelletto. Si notano, quindi, le stratificazioni: Lo strato molecolare, quello più esterno, con la presenza di fibre parallele e l’interconessione in orizzontale all’interno dello stesso strato; Lo strato delle cellule gangliari, in cui si posizionano i nuclei delle cellule del Purkinje; Lo strato dei granuli, più in profondità. In ogni punto del cervelletto una biopsia della corteccia cerebellare mostra sempre e comunque questi tre strati. La semplicità morfologica della corteccia cerebellare rende ragione del fatto che non esiste una perturbazione dell’attività elettrica cerebellare (la così detta “epilessia cerebellare”), perché il cervelletto non dà luogo a manifestazioni parossistiche, come sono quelle caratteristiche dell’epilessia che sono, invece, esclusive del cervello, nel quale la notevole complessità degli strati di corteccia, che presenta una pluristratificazione fino a sei strati, e la complessità cellulare citoarchitettonica e morfologica fanno sì che la corteccia cerebrale possa andare incontro a queste manifestazioni parossistiche, epilettiche (dal greco epilambanein= giungere all’improvviso, sopraggiungere senza alcun preavviso). In questo preparato istologico, riconosciamo la presenza dei pirenofori delle cellule del Purkinje e, in alto, lo strato molecolare con le cellule dei canestri, le fibre rampicanti e le fibre parallele. Più in basso, sorpassate le cellule del Purkinje, ritroviamo lo strato dei neuroni granulari. ARCHICEREBELLUM L'Archicerebellum, dal punto di vista morfologico, è contraddistinto da formazioni vermiane e dal lobo flocculo-nodulare (costituito da nodulo e flocculi, particolarmente sviluppati nei volatili e nelle scimmie arboricole). A livello del lobo, quindi, flocculo-nodulare troviamo una prevalenza di affluenze vestibolari e questo è spiegabile proprio perché a questo livello devono arrivare le informazioni relative all’equilibrio, onde gestire e modificare l’equilibrio in relazione al volo o al salto da un ramo ad un altro. Le afferenze derivano dal ganglio vestibolare di Scarpa che emette un prolungamento che trae informazioni dai recettori vestibolari situati all’interno della componente vestibolare dell’orecchio interno. L'informazione che, invece, risale verso le formazioni nucleari vestibolari che si trovano nel bulbo al limite con il ponte e verso il lobo flocculo-nodulare del cervelletto prendono la via delle frecce di colore azzurro. Questo è particolarmente importante per l’integrazione delle informazioni in rapporto alla necessità della correzione dei movimenti, che potrebbe essere necessaria in caso di deviazione della traiettoria o di perdita di equilibrio. Tutta questa complessa afferenza si traduce poi in un collegamento con altre strutture, come la formazione reticolare bulbo-pontina, la formazione bulbo-pontina controlaterale e i nuclei vestibolari, sempre per integrare le informazioni e attivare risposte opportune (come quelle che vanno ad innescare l’attivazione dei nuclei motori, responsabili del controllo dei movimenti conseguenti alla modificazione della traiettoria o dei gruppi muscolari inizialmente coinvolti). Questa importante attività di correzione e modificazione, di controllo da parte del archicerebellum si esplica poi attraverso le vie vestibolo-spinali, cioè circuiti che consentono la risposta che viene modulata attraverso il lobo flocculo-nodulare e, soprattutto, i nuclei vestibolari, e che va ad innescare le opportune risposte, con l’attivazione dei muscoli coinvolti in questi cambiamenti e in queste condizioni di attivazione o inibizione del reclutamento di unità motorie a seconda delle condizioni che si vengono a verificare di volta in volta. In questa immagine viene proposto l’insieme delle proiezioni dei nuclei vestibolari che presentano anche dei collegamenti con i nuclei di alcuni nervi dell’oculomozione (in questo caso, l’oculomotore comune e il trocleare). Questo è particolarmente importante anche in rapporto alla necessità di coordinare il controllo visivo della situazione alla stimolazione dovuta a perturbazione dell’equilibrio. È chiaro che se viene innescata una perturbazione dell’equilibrio il controllo visivo ci consente di perfezionare la risposta di cui abbiamo bisogno per evitare danni al nostro corpo. PALEOCEREBELLUM Si occupa di gestire il tono dei muscoli e questo fa sì che un’importanza notevole nell’afferenza ad esso di impulsi che provengono dalla periferia del corpo sia appannaggio di quelle fibre che raccolgono le informazioni dai fusi neuromuscolari e dagli organi muscolo-tendinei del Golgi. Questi particolari organi danno un contributo essenziale all’informazione delle strutture paleocerebellari da parte del rilevamento che questi microscopici organi svolgono sia a livello dei ventri muscolari sia a livello della zona di giunzione tra tendine e muscolo. Come si comportano questi particolari organuli? Gli organi muscolo-tendini sono posizionati con delle terminazioni nervose a livello della zona di giunzione tra tendine e muscolo e vanno a rilevare l’entità dello stiramento, dell’entità della sollecitazione forniscono un’importante informazione al sistema nervoso centrale circa l’entità dello sforzo e della sollecitazione a cui è sottoposto il tendine. L'attivazione degli organi muscolo-tendinei del Golgi consente di informare il cervello dell’importanza o gravità della sollecitazione che il tendine sta subendo in modo tale da rendere dotto il cervello stesso dell’opportunità o meno di proseguire nello sforzo che il tendine sta compiendo. È un meccanismo di salvaguardia per evitare rottura, lacerazione, stiramento del tendine che avrebbe conseguenze gravi. Per quanto riguarda i fusi neuromuscolari, questi hanno forma fusata e presentano al loro interno delle terminazioni anulospinali, che si avvolgono a spirale attorno alle fibre intrafusali, le quali sono sensibili al grado di contrazione delle fibre muscolari scheletriche striate che si trovano all'esterno, nelle immediate vicinanze del fuso neuromuscolare. Cosa succede in particolare? La fibra muscolare scheletrica (nell'immagine viene preso come esempio il muscolo quadricipite femorale) viene attivata tramite l'informazione che il fuso neuromuscolare fornisce al midollo spinale attraverso la branca periferica della fibra che proviene dal nucleo del neurone gangliare (che si trova nel ganglio spinale). La terminazione anulospinale del fuso raccoglie l'informazione che concerne la sollecitazione, l'attivazione, la contrazione, la distensione del muscolo quadricipite. Questa informazione viene veicolata fino al ganglio, dal ganglio riparte, entra nel corno posteriore e va a raggiungere o direttamente il motoneurone alfa, andando a potenziare e modulare l'attivazione delle stesse fibre del muscolo quadricipite (in rapporto, per esempio, all’esecuzione di un movimento in cui l'attivazione dei centri muscolari del quadricipite richieda un coordinamento maggiore per un movimento più complesso), oppure un interneurone inibitorio, e quindi si va a ‘staccare la spina” al motoneurone che normalmente attiva il muscolo antagonista (che, nel caso del quadricipite, si trova a livello della faccia posteriore della coscia). Questo, in altre parole, determina che all'attivazione del muscolo quadricipite femorale, con conseguente attivazione ulteriore delle fibre muscolari del muscolo medesimo, si associa a un’inibizione, uno spegnimento, del muscolo antagonista. Questo è fondamentale perché solo in questo modo si può ottenere la contrazione del quadricipite, inibendo l'attività contrattile del muscolo antagonista. Questo fenomeno possiamo riscontrarlo anche altrove, ad esempio a livello del braccio, quando consideriamo l'attivazione del muscolo bicipite e l'inibizione del tricipite (antagonista) quando si crea la flessione dell'avambraccio sul braccio; avviene esattamente l'opposto quando andiamo a svolgere l’attività dell’estensione dell’arto superiore, in cui si ha la contrazione del tricipite e l’inibizione del bicipite omerale. NEOCEREBELLUM Quando andiamo a considerare le fibre che trasportano le informazioni a livello del circuito neocerebellare, il neocerebello ha un'attività di controllo molto più complessa che deve servire a coordinare più gruppi muscolari anche in modo da innescare un’attività coordinata, facendo capo al nucleo dentato. Questo è molto importante nella gestione dei movimenti coordinati e sincronizzati degli arti, nonché nell'attivazione contrattile della muscolatura degli stessi. Il nucleo dentato, infatti, in caso di condizione di spasticità, può essere lesionato durante una procedura stereotassica in modo tale da garantire una buona riduzione dell’eccesso di contrazione dei muscoli che è alla base della spasticità stessa. AFFERENZE ED EFFERENZE DEL NUCLEO ROSSO A proposito del tronco cerebrale, era stata mostrata questa immagine che mostrava il mesencefalo: Ma non avevamo chiarito quali fossero le strutture mesencefaliche che vediamo schematizzare in questa immagine: È visibile la lamina quadrigemina (in alto), formata dai quattro tubercoli quadrigemini superiori; l'acquedotto di Silvio, al centro della calotta mesencefalica; le formazioni della calotta, tra le quali riconosciamo il nucleo rosso, grosso, tondeggiante e voluminoso, e altri nuclei, come quello di Edinger-Westphal (quello della componente vegetativa del terzo nervo cranico). Scendendo ancora più in basso, ritroviamo le formazioni della sostanza nera di Sommering e il cosiddetto piede del ladruncolo cerebrale. I due peduncoli cerebrali divergono lateralmente formando la fossa interpeduncolare e, in essi, troviamo sistemi di fibre. In particolare, nei 3/5 medi del piede del peduncolo cerebrale troviamo le fibre del fascio piramidale; a livello del quinto laterale e del quinto mediale troviamo le fibre del sistema extrapiramidale come fascio fronto-pontino di Arnold e fascio temporo-pontino. Il collegamento che a livello della calotta mesencefalica ci interessa maggiormente è il collegamento del nucleo rosso. Il nucleo rosso è rappresentato da due strutture quadrangolari concentriche, con all’esterno la parte parvicellulare e all’interno la parte magnicellulare (cioè che contiene neuroni di dimensioni maggiori). Notiamo che i collegamenti sono molteplici. Anzitutto c’è un collegamento che parte sia dalla parte parvicellulare che da quella magnicellulare attraverso il fascio rubro-spinale, che va dal nucleo rosso al midollo spinale e rappresenta una via discendente del sistema extrapiramidale. Una parte delle fibre parvicellulari raggiunge, però, la formazione reticolare bulbare e le olive bulbari, attraverso il reticolo-spinale e olivo-spinale. Un'altra porzione, invece, va alla formazione reticolare mesencefalica e poi al talamo. Dunque, queste sono tutte vie che fuoriescono dal nucleo rosso come vie extrapiramidali (indicate in figura col rosso) che controllano la motilità involontaria automatica associata, quindi contribuiscono a regolare l’attività del sistema piramidale. Viceversa, i sistemi afferenti (indicate in figura con l’azzurro) al nucleo rosso possono derivare sia dal cervelletto, come fibre cerebellorubre (possono provenire dai nuclei globoso ed emboliforme ed arrivare alla componente parvicellulare), sia dal nucleo dentato, come fibre dentatorubre (arrivano sia alla componente parvicellulare che magnicellulare). Abbiamo poi delle afferenze dal talamo (arrivano alla componente parvicellulare), dai nuclei del telencefalo (detti “nuclei o gangli della base”), in particolare dal nucleo lenticolare come fibre lenticolorubre), e, infine, delle afferenze dalla corteccia cerebrale al nucleo lenticolare e al nucleo rosso, come fibre corticolenticolari o corticorubre. C'è quindi un’interconnessione complessa di queste informazioni che in parte provengono dal midollo spinale e arrivano ai nuclei del cervelletto come fibre spinocerebellari, le quali poi proiettano al nucleo rosso, il quale deve poi innescare delle risposte attraverso le vie efferenti che sono, appunto, il fascio rubrospinale, il fascio reticolospinale, il fascio olivospinale. Queste correzioni dovute alle sollecitazioni da parte dei nuclei vestibolari e da parte del cervelletto hanno bisogno di un’integrazione superiore attraverso l’azione del talamo e, soprattutto, della corteccia cerebrale che va a perfezionare, ad “aggiustare” l’attività del nucleo rosso e dei nuclei della base. Ecco quindi la complessa interconnessione che regola buona parte dell’attività del sistema extrapiramidale. Infatti, come vediamo anche da questa immagine, dal nucleo rosso e dalla componente parvicellulare prendono origine quei circuiti a cui abbiamo accennato prima, cioè la via rubrospinale (molto importante, che si porta contro lateralmente) e le altre vie extrapiramidali, come il tratto olivo-spinale, rubro-olivare, olivo-spinale rubro-reticolare (dalla formazione reticolare del bulbo al midollo spinale attraverso la via reticolo-spinale). Queste ultime tre vie, quindi, sono quelle attraverso cui il nucleo rosso regola l’attività di controllo extrapiramidale di gestione e coordinamento, quindi dei movimenti volontari automatici associati che sono collegati alla modulazione dell’attività motoria volontaria. Nell'immagine vediamo in azzurro i fasci ascendenti del midollo spinale e in rosso quelli discendenti. Vediamo come, a parte il fascio corticospinale laterale (molto voluminoso, rappresenta il 75% della via corticospinale piramidale) e il fascio piramidale diretto che sono i più rilevanti per la totalità delle fibre piramidali, gli altri sono tutti fasci del sistema extrapiramidale. In questa immagine schematica, vediamo la distribuzione dei fasci nella sostanza bianca dei cordoni, in particolare quello laterale e quello anteriore.