Corso di Istologia ed Embriologia
June 4, 2011
Anno Accademico 2008-2009, Semestre I
1
Contents
I
Istologia
4
1 La cellula
1.1
1.2
4
Il nucleo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
1.1.1
La cromatina . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
1.1.2
Il nucleolo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
1.1.3
L'involucro nucleare
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
Gli organuli interni . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
1.2.1
Il reticolo endoplasmatico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4
1.2.2
L'apparato del Golgi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5
1.2.3
I lisosomi
1.2.4
I mitocondri
1.2.5
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
6
Il citoscheletro
1.2.5.1
I microtubuli . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
6
1.2.5.2
I microlamenti
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
6
1.2.5.3
I lamenti intermedi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
6
2 Il tessuto epiteliale
6
2.1
Gli epiteli di rivestimento
2.2
Specializzazioni di membrana
2.3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
7
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
8
Ghiandole esocrine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
8
3 Il tessuto connettivo
3.1
7
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Gli epiteli ghiandolari
2.3.1
5
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
9
Connettivi propriamente detti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.1.1
Tessuto connettivo lasso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.1.2
Tessuto connettivo denso
9
10
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
10
3.2
Il tessuto adiposo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
10
3.3
Il sangue . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
10
3.4
La cartilagine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
11
3.5
3.4.1
La cartilagine ialina
3.4.2
La cartilagine elastica
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
12
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
12
3.4.3
La cartilagine brosa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
12
3.4.4
Istogenesi e accrescimento . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
12
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
12
L'osso
3.5.1
L'osso compatto
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
13
3.5.2
L'ossicazione . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
13
4 Il tessuto muscolare
4.1
13
Il muscolo scheletrico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
13
4.1.1
14
Il sarcomero . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.2
Il muscolo cardiaco . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
15
4.3
Il muscolo liscio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
15
5 Il tessuto nervoso
15
5.1
Il neurone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
16
5.2
La guaina mielinica . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
16
5.3
La sinapsi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
16
5.4
Cellule gliali . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
16
2
II
Embriologia
17
6 Introduzione alla riproduzione
17
6.1
La determinazione del sesso . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
17
6.2
La gametogenesi clinica
18
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
7 L'ovogenesi
18
7.1
La fase di inizio e la fase di arresto . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
18
7.2
La fase di ripresa . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
18
7.2.1
19
La fase follicolare . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
7.2.1.1
La fase preantrale
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
19
7.2.1.2
La fase antrale . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
19
7.2.1.3
La fase preovulatoria
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
19
7.3
L'ovulazione . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
19
7.4
La fase luteinica
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
20
7.5
Gli altri cicli femminili . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
20
8 La spermatogenesi
20
8.1
La fase di proliferazione
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
8.2
La fase di meiosi
21
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
21
8.3
La fase di spermiogenesi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
21
8.4
Organizzazione spaziotemporale della spermiogenesi
. . . . . . . . . . . . . . . . . . . . . . . . . . .
22
8.5
Controllo ormonale . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
22
9 Le prime quattro settimane di sviluppo
23
10 La prima settimana di sviluppo
23
11 La seconda settimana di sviluppo
24
12 La terza settimana di sviluppo
25
12.1 La notocorda
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
25
12.2 La neurulazione . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
26
12.3 La suddivisione del mesoderma
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
26
12.4 La circolazione primitiva . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
27
13 La quarta settimana di sviluppo
27
13.1 Evoluzione dell'ectoderma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
28
13.2 Evoluzione dell'endoderma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
28
13.3 Evoluzione del mesoderma . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
29
3
Part I
Istologia
1
La cellula
1.1
Il nucleo
1.1.1 La cromatina
La cromatina è costituita da DNA e da proteine basiche dette istoni. Queste proteine formano strutture globulari
dette nucleosomi, attorno ai quali si avvolge il DNA.
Se i nucleosomi sono molto addensati tra loro si parla di eterocromatina, se lo sono meno di eucromatina.
L'eucromatina è poco elettrondensa ed è la porzione di DNA disponibile alla trascrizione mentre l'eterocromatina
essendo fortemente compattata non è trascrivibile e quindi inattiva.
1.1.2 Il nucleolo
Il nucleolo è una regione granulare densa all'interno del nucleo ed è formato da DNA, RNA e proteine; è il sito di
produzione dei ribosomi.
1.1.3 L'involucro nucleare
Questo involucro, che delimita il nucleo ed è caratteristico dei soli eucarioti, è formato da due membrane che sono
una estensione e una specializzazione delle membrane del reticolo endoplasmatico rugoso.
L'involucro nucleare è attraversato da numerosi pori ch epermettono la comunicazione tra nucleo e citoplasma.
1.2
Gli organuli interni
Gli organuli interni sono i compartimenti distinti in cui la cellula eucariotica è funzionalmente e morfologicamente
suddivisa. Sono tutti circondati da membrane e contengono enzimi e molecole caratteristiche.
Gli organuli presenti in una cellula umana sono:
Reticolo endoplasmatico
Apparato del Golgi
Lisosomi
Mitocondri
Perossisomi
1.2.1 Il reticolo endoplasmatico
Il reticolo endoplasmatico è un sistema di membrane organizzate in cisterne per lo più parallele tra loro. Sintetizza
quasi tutti i lipidi della cellula e sulla sua faccia citosolica le proteine destinate a:
Secrezione
Golgi
Lisosomi
Membrana plasmatica
4
Il reticolo viene suddiviso in liscio e rugoso; il reticolo rugoso presenta i ribosomi ed è sede sia della sintesi lipidica
che di quella proteica, contrariamente a quello liscio che si occupa solamente della sintesi dei lipidi.
Una delle attività fondamentali del reticolo è la glicosilazione aspecica delle proteine, cioè appena una proteina
entra nel suo lume, viene glicosilata senza distinzione.
1.2.2 L'apparato del Golgi
L'apparato del Golgi è un organulo costituito da membrane a cisterne e organizzato in quattro sezioni con compiti
dierenti:
1. Golgi Cis
2. Golgi mediale
3. Golgi trans
4. Trans Golgi Network (TGN)
Le vescicole contenenti le proteine secrete dal reticolo vengono direzionate al Golgi Cis, e da qui iniziano un percorso
a tappe forzate attraverso tutta la sequenza di sezioni; non è possibile saltare una tappa o tornare indietro a causa
della presenza di recettori specici sulle vescicole: una vescicola può legarsi solo alla sezione del Golgi verso la quale
è diretta poichè possiede solo le sue molecole segnale.
L'attività secretiva dell'apparato del Golgi segue fondamentalmente tre vie:
1. Secrezione costitutiva: serve a rinnovare la membrana cellulare e sfrutta il processo di esocitosi
2. Secrezione regolata: serve a secernere particolari sostanze solo quando ve ne è bisogno
3. Smistamento ai lisosomi: crea i lisosomi
La secrezione costitutiva è bilanciata dal processo di endocitosi; questa attività serve a rifornire il Golgi di materiale
con cui creare le vescicole e può, come la secrezione, essere costitutiva o mediata da recettori specici.
1.2.3 I lisosomi
I lisosomi sono vescicole rivestite da membrana contenenti enzimi idrolitici per la digestione intracellulare delle
macromolecole; gli enzimi sono delle idrolasi acide attive a pH 5, che è il pH interno al lisosoma.
La membrana di questi organuli contiene pompe ATP-dipendenti per mantenere basso il pH, proteine di trasporto
per espellere i prodotti della digestione, e numerosi glucidi legati ai lipidi e alla proteine della faccia interna con
funzione di protezione da un'eventuale autodigestione.
I lisosomi originano dalle vescicole idrolasiche del Golgi (o lisosomi primari), strutture prodotte dal Golgi e
contenenti gli enzimi ma non le pompe; diventeranno lisosomi a tutti gli eetti quando si fonderanno con un
endosoma, che porta con se le pompe per abbassare il pH.
L'endosoma necessario alle vescicole per attivarsi può provenire da fagocitosi (fagosoma), da endocitosi (endosoma) o da autofagia (autofagosoma).
1.2.4 I mitocondri
I mitocondri sono organuli cilindrici formati da due membrane e lunghi da 0,5 a 2
µm;
hanno un DNA di tipo
batterico (procariotico) e sintetizzano una piccola parte delle loro proteine, intorno al 5%.
Questi organuli contengono enzimi in grado di sfruttare l'ossigeno molecolare per ossidare completamente le
molecole organiche. L'attività di ossidazione non è esclusiva dei mitocondri, può avvenire infatti anche nel citosol,
ma in questo caso la resa è inma.
5
1.2.5 Il citoscheletro
Il citoscheletro è formato da tre classi di proteine del citosol che si aggregano a formare lamenti specici:
1. Microtubuli: Ø25nm, composti da tubulina
2. Microlamenti: Ø5nm, composti da actina
3. Filamenti intermedi: Ø10nm, formati da proteine diverse che variano a seconda del tipo di cellula
1.2.5.1 I microtubuli
β.
I microtubuli sono formati da 13 protolamenti di subunità alternate di tubulina
α
e
Sono strutture polari che formano un reticolo di sostegno per gli organuli; sono inoltre una via preferenziale di
movimento per le vescicole, costituiscono il fuso mitotico e si trovano in ciglia e agelli a dare loro mobilità.
La polimerizzazione della tubulina è legata a GTP, guanosintrifosfato, e procede per strutture dimeriche: l'intero
lamento è veloce sia in fase di crescita che di accorciamento.
I microtubuli presentano un estremità negativa costituita da un terzo tipo di tubulina, la tubulina
γ:
questa
estremità è sempre ancorata a un centro organizzatore, pertanto la crescita si ha solo in direzione del lato positivo.
Associate ai microtubuli vi sono delle proteine, della classe delle MAP (microtubes associated proteins) che
controllano la lunghezza, la conformazione tridimensionale e la distanza delle singole strutture.
La funzione di via preferenziale di movimento è possibile grazie a due proteine speciche che scorrono lungo i
microtubuli nelle due possibili direzioni: la chinesina (da - a +) e la dineina (da + a -).
1.2.5.2 I microlamenti
I microlamenti sono formati dalla proteina globulare actina G, che polimerizza in
lamenti polari. Sono presenti in tutte le cellule, anche se maggiormente in quelle contrattili, e formano una rete
che stabilizza la forma cellulare ma ne rende al contempo possibile un cambiamento in tempi brevi.
La struttura del microlamento completo non è stabile: in ogni istante vi è piuttosto un equilibrio tra le subunità
di actina entrante e quelle uscenti.
La rete tridimensionale creata dai microlamenti è controllata da proteine che formano il complesso ARP, che
costringe due lamenti a legarsi tra loro con un angolo di circa 70°.
La crescita dei microlamenti è il risultato della competizione di due proteine: la prolina, che stimola la crescita,
e la timosina, che invece la ostacola.
Le cellule che hanno grande necessità di cambiare la loro forma attuano una strategia per massimizzare la
velocità di costruzione dei lamenti; un singolo lamento presenta una sola estremità da cui crescere, ma se quello
stesso lamento viene tagliato in due si avrà lo stesso numero di monomeri ma con due estremità: le estremità extra
possono essere protette da crescita incontrollata da una proteina, la gelsolina, che può facilmente essere rimossa in
caso di necessità, come fanno ad esempio le piastrine.
1.2.5.3 I lamenti intermedi
I lamenti intermedi sono caratteristici della tipologia di cellula che li ospita e
quindi sono sucienti a riconoscere la natura istologica di una cellula in esame, caratteristica fondamentale nello
studio dei tumori trasformati.
I più importanti tipi di lamenti intermedi sono:
Filamento
2
Localizzazione
Citocheratine
Cellule epiteliali
Desmina
Muscolo liscio e striato
Proteine brillari acide della glia
Astrociti
Proteine neurolamento
Neuroni
Lamina nucleare
Nucleo di ogni cellula
Vimentina
Molti tessuti di origine mesodermiale
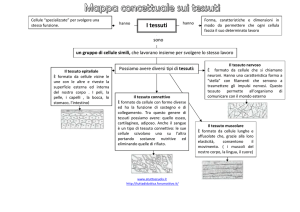
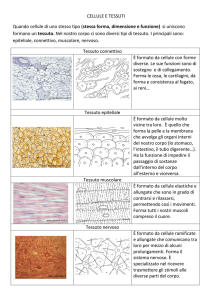
Il tessuto epiteliale
Il tessuto epiteliale è caratterizzato da cellule a stretto contatto tra loro, la cui coesione è assicurata da giunzioni
intercellulari adesive speciche.
6
La matrice extracellulare è molto scarsa e visibile solamente al microscopio elettronico.
Tra le cellule epiteliali non vi sono vasi sanguigni poichè non vi è spazio suciente:
nell'ordine dei 5-6
µm
i vasi più piccoli sono
mentre lo spazio tra le cellule è nell'ordine dei nanometri.
Tutti gli epiteli sono a contatto con un tessuto connettivo sottostante che fornisce loro nutrimento essendo
riccamente vascolarizzato: esiste una lamina basale a separare i due tipi di tessuto.
Le funzioni del tessuto epiteliale sono fondamentalmente due:
Rivestimento
Secrezione
→
→
epitelio di rivestimento
epitelio ghiandolare
A livello di embriogenesi gli epiteli derivano da tutti e tre i foglietti embrionali, ciascuno specializzato nel dar vita
a una diversa area:
Ectoderma: rivestimento della cute, del naso, di parte della bocca e dell'ano, ghiandole cutanee
Mesoderma: rivestimento dei vasi e delle sierose
Endoderma: rivestimento del canale respiratorio e digerente
2.1
Gli epiteli di rivestimento
Gli epiteli di rivestimento si classicano in base al numero di cellule in:
Epitelio semplice o monostraticato = singolo strato cellulare
Epitelio composto o pluristraticato = strati cellulari in numero da due in su
In base alla forma delle cellule si riconoscono invece:
Epitelio squamoso = cellule molto più larghe che alte, con caratteristico nucleo allungato
Epitelio cubico o isoprimatico = cellule con tre dimensioni sostanzialmente uguali
Epitelio cilindrico o batiprismatico = cellule molto più alte che larghe, con caratteristico nucleo basale
Esiste una classe di epiteli di rivestimento le cui cellule più distanti dalla lamina basale producono un eccesso di
keratina al punto da venirne uccise formando così una barriera di cellule morte: si tratta degli epiteli cheratinizzati,
che si formano con un processo di apoptosi continuo.
Gli epiteli cheratinizzati svolgono ottime funzioni di protezione essendo difesi meccanicamente e chimicamente:
oltre alla keratina vengono infatti secreti dei lipidi che garantiscono l'impermeabilità dello strato.
Gli epiteli di rivestimento svolgono per lo più due funzioni principali:
Protezione
Scambio
La prima funzione è solitamente deputata agli epiteli spessi, la seconda a quelli sottili.
2.2
Specializzazioni di membrana
La membrana cellulare di uno strato epiteliale presenta tre zone diverse:
Zona apicale
Zona laterale
Zona basale
7
Ciascuna di queste zone compie attività dierenti e presenta strutture speciche diverse.
La membrana laterale presenta quattro specializzazioni fondamentali alla funzionaltà della cellula: zonula occludens, zonula adhaerens, macula adhaerens e gap junction.
La zonula occludens è una giunzione presente lungo l'intera circonferenza cellulare in prossimità della zona
apicale; la sua funzione è quella di sigillare gli spazi intercellulari in modo da costringere ogni sostanza ad attraversare
la cellula. La zonula occludens è inoltre una barriera che impedisce alle altre specializzazioni di membrana laterale
di raggiungere la membrana apicale.
La zonula adhaerens è una cintura sottile e continua posta sotto la zonula occludens e serve a garantire adesione
tra le cellule contigue anche quando vanno incontro a modiche nella forma durante l'embriogenesi: fondamentalmente si tratta di collegamenti a base di caderina che legano lamenti di actina intracellulari.
La macula adharens, o desmosoma, è invece una struttura non continua che serve a mantere la forma delle
cellule e a distribuire gli sforzi meccanici sulle cellule sottostanti anzichè scaricarli su una piccola area. Un mezzo
desmosoma, o emidesmosoma, è presente sulla membrana basale ed è a metà poichè manca un'altra cellula in grado
di fornire i lamenti di citocheratina per chiuderlo.
Le gap junction sono fondamentalmente dei fori che permettono il libero transito tra cellule di aminoacidi,
nucleotidi, monosaccaridi e ioni: caratteristica importante è che non prevedono un meccanismo di chiusura. Tecnicamente sono strutture dette connessoni, ciascuno formato da 6 unità proteiche dette connessine. Il diametro di
una di queste strutture è pari a 1,5 nm.
2.3
Gli epiteli ghiandolari
Gli epiteli ghiandolari sono gli epiteli specializzati nell'attività di secrezione. Esistono due tipologie di ghiandole:
Ghiandole esocrine: emettono i loro secreti direttamente nell'ambiente esterno e sono caratterizzate da una
parte secernente, l'adenomero, e da una parte che porta all'esterno il secreto, il dotto escretore.
Ghiandole endocrine: emettono i loro secreti nel usso sanguigno: vi passano all'interno dei vasi e quindi sono
prive di dotto escretore.
2.3.1 Ghiandole esocrine
Le ghiandole esocrine vengono classicate secondo quattro diversi parametri.
Secondo la forma degli adenomeri si riconoscono:
Ghiandole alveolari, con adenomero tondo e con lume interno sviluppato
Ghiandole acinose, con adenomero tondo e lume interno sottile
Ghiandole tubolari, con adenomero allungato e lume interno sottile
Ghiandole glomerulari, con adenomero a forma di gomitolo
In base alla ramicazione dei dotti escretori si riconoscono
Ghiandole semplici, con un dotto e un adenomero
Ghiandole ramicate, con un dotto e più adenomeri
Ghiandole composte, con più dotti e più adenomeri
In base alle modalità di secrezione si riconoscono
Ghiandole merocrine, in cui il secreto è emesso per esocitosi
Ghiandole apocrine, in cui il secreto è emesso in vescicole ottenute strozzando il citoplasma
Ghiandole olocrine, in cui la cellula va in apoptosi e staccandosi e perdendo le membrane rilascia i secreti
8
L'ultima catalogazione è quella per natura chimica del secreto
Ghiandole a secrezione sierosa, con secreto proteico, si riconoscono da un nucleo tondo e basale
Ghiandole a secrezione mucosa, con secreto zuccherino, si riconoscono da un nucleo schiacciato e basale
Ghiandole a secrezione mista, con secreto di natura mista poichè presentano adenomeri mucosi e adenomeri
sierosi (non è sinonimo di ghiandola mista!)
3
Il tessuto connettivo
Sotto la categoria dei tessuti connettivi si raggruppano tessuto con comune origine embrionale, derivano infatti tutti
dal mesoderma, con comune organizzazione strutturale e con comune funzione, che può essere di:
Connessione
Sostegno
Nutrizione
Difesa
Ogni tipo di tessuto connettivo non le rappresenta tutte, ma solo alcune e in maniera più o meno sviluppata.
Caratteristica fondamentale di ogni connettivo è la presenza di una notevole quantità di matrice extracellulare
che può essere brosa o amorfa.
I tessuti connettivi sono fondamentalmente sei, due propriamente detti (popolazione cellulare eterogenea), due
liquidi (bre assenti) e due di sostegno (pochi tipi cellulari, matrice compatta):
Tessuto connettivo lasso, con poche bre, molte cellule
Tessuto connettivo denso, molte bre, poche cellule
Sangue
Linfa
Cartilagine, matrice gelatinosa
Osso, matrice mineralizzata
3.1
Connettivi propriamente detti
Nei connettivi propriamente detti le bre sono più o meno organizzate e presentano una sostanza fondamentale
viscosa.
La popolazione è divisa tra cellule sse (broblasti, macrofagi ssi, adipociti, melanociti), con ruoli di
omeostasi, riparazione, difesa e immagazzinamento energetico, e cellule migranti (macrofagi liberi, mastociti, granulociti, linfociti, plasmacellule), legate alla situazione ambientale.
Le bre dei connettivi sono in generale di tre tipi:
Fibre collagene, formate da proteine del collagene
Fibre reticolari, formate da proteine del collagene
Fibre elastiche, le uniche formate da elastina e brillina
Il collagene è una classe di proteine suddivisa in varie tipologie, di cui le più importanti sono:
Collagene I: forma il derma, le ossa, i tendini e le cartilagini brose
Collagene II: forma la cartilagine ialina e i dischi intervertebrali
9
Collagene III: forma il tessuto connettivo reticolare
Collagene IV: forma le lamine basali
Poichè tende naturalmente ad aggregarsi in polimeri troppo voluminosi, il collagene viene secreto all'interno della
cellula come procollagene e attivato solamente in ambiente extracellulare.
µm,
Le bre collagene hanno un diametro compreso tra 1 e 20
sono essibili e molto resistenti alla trazione, si
aggregano a formare fasci e sono quasi sempre presenti nei connettivi propriamente detti oltre ad essere componenti
essenziali di cartilagine ialina, brosa e osso.
Le bre reticolari sono sottili, con diametro tra 0,5 e 2
µm,
e si presentano associate a glicoproteine; sono
presenti in grande quantità in endonervrio ed endomisio.
Le bre elastiche sono costituite da brillina ed elastina e sotto trazione si allungano per poi ritornare alle
dimensioni di partenza.
La loro deformabilità è limitata e protetta dalle bre collagene: sono abbondanti nella
tonaca elastica delle arterie e dei legamenti.
La matrice extracellulare del connettivo propriamente detto contiene una quantità variabile di acqua legata alla
quantità di proteoglicani: essa diminuisce naturalmente con l'età e aumenta invece di molto durante l'edema e
l'inammazione.
Questa sostanza ha funzioni troche, in quanto in essa si compiono gli scambi nutritivi, difensive, in quanto la
sua viscosità è un ostacolo per i batteri, e meccaniche, poichè cementa le bre.
3.1.1 Tessuto connettivo lasso
Le caratteristiche fondamentali sono le poche bre e l'abbondante matrice extracellulare amorfa; sono presenti molte
cellule, per lo più broblasti e macrofagi.
Si trova generalmente sotto gli epiteli, intorno ai vasi e ai fasci muscolari e nelle vicinanze delle ghiandole.
3.1.2 Tessuto connettivo denso
Questo tessuto è per la maggior parte occupato da bre e si distingue in regolare e irregolare.
Nel tessuto connettivo denso regolare le bre sono strettamente impacchettate e allineate lungo le linee di
trazione; poichè molto resistente forma i tendini e i legamenti.
Nel tessuto connettivo denso irregolare le bre sono intrecciate casualmente e si trova soprattutto nel derma,
nelle capsule brose degli organi, nelle guaine dei tendini e dei nervi e nel periostio esterno.
3.2
Il tessuto adiposo
Nel tessuto adiposto le cellule più rappresentate sono gli adipociti, cellule di dimensioni comprese tra 50 e 150
µm
che accumulano al loro interno moltissimi trigliceridi.
Si riconosce il tessuto adiposo uniloculare o bianco, praticamente l'unico nell'adulto, e il tessuto multiloculare o
bruno, presente solo alla nascita e in sedi limitate.
Mentre il tessuto bruno ha solo funzione termica, quello bianco ha funzione di deposito energetico, di modellamento del corpo, di ammortizzazione, di isolamento termico e di sostegno.
3.3
Il sangue
Il sangue è un tessuto connettivo uido rinchiuso in un sistema di vasi comunicanti.
È costituito da una parte
liquida, il plasma, e da una serie di elementi gurati comprendenti globuli rossi, globuli bianchi e piastrine.
Il volume del sangue in un individuo adulto è pari a circa il 7-8% del suo peso corporeo.
10
Eritrociti
Piastrine
Neutroli
Eosinoli
Basoli
Monociti
Linfociti
Dimensioni (µm)
7
2-3
9-15
12-17
10-14
15-20
7-16
Vita media
4 mesi
10 giorni
1-2 giorni
1-2 giorni
ore-giorni
3 giorni
3 giorni-20 anni
Conta leucocitaria
99% degli e.f.
//
60%
1-3%
0-1%
4-10%
20-30%
5 ∗ 106
3 ∗ 105
Quantità per
µL
7 ∗ 103
Table 1: Elementi gurati del sangue
Gli eritrociti sono cellule prive di nucleo e di qualsiasi organulo citoplasmatico. Contengono emoglobina e hanno
un citoscheletro che permette loro di essere facilmente deformabili: quanto questa capacità viene meno vengono
distrutti dalla milza. La dimensione media è di 7
µm.
I leucociti, o globluli bianchi, si dividono in granulociti e agranulociti. I granulociti si suddividono a loro volta
in:
Granulociti neutroli
Granulociti eosinoli (=acidoli)
Granulociti basoli
I leucociti agranulociti si dividono invece in:
Monociti
Linfociti
I granulociti neutroli hanno un diametro compreso tra i 12 e i 14
µm, hanno un nucleo plurilobato e un citoplasma
pieno di granuli contenenti enzimi lisosomiali; sono estramementi mobili e hanno una spiccata attività fagocitaria:
la loro vita è inferiore alle dodici ore.
I granulociti eosinoli hanno un nucleo bilobato e rimangono in circolo per circa 6-10 ore prima di migrare nel
connettivo e li sopravvivervi per 8-12 giorni. Non fagocitano i batteri ma eliminano i complessi antigene-anticorpo
formati nelle reazioni allergiche.
I granulociti basoli hanno un nucleo ovoidale o bilobato con granulazioni dense e molto grandi, producono
eparina ed istamina e sono responsabili delle allergie.
I monociti hanno un nucleo reniforme e rimangono in circolo 1-4 giorni prima di migrare nel connettivo per
diventare macrofagi liberi.
Sono in grado di unirsi in una cellula fagocitaria gigante e sono le cellule che danno
origine a osteo e condroclasti.
I linfociti sono di piccole dimensioni (8-10
µm)
con un grande nucleo e un citoplasma sottile.
Sono gli unici
responsabili dell'immunità specica e hanno una lunga vita pur non essendo terminali. Si suddividono in linfociti
T e linfociti B.
I linfociti B una volta attivati si trasformano in plasmacellule e producono anticorpi; i linfociti T sono divisi in
T-Helper, che aiutano i B nella risposta umorale, e T-Citotossici, che uccidono le cellule infettate da virus.
Le piastrine sono piccoli (2-4
µm)
elementi privi di sostanza nucleare in numero di
2 − 4 ∗ 105
per mmc e vita
media tra gli otto e i dieci giorni. Sono prodotte nel midollo osseo per frammentazione del citoplasma di enormi
(200
3.4
µm)
cellule polinucleate dette megacariociti.
La cartilagine
La cartilagine è un connettivo di sostegno formato da soli condrociti circondati da una matrice extracellulare
compatta contenente bre, proteoglicani e glicoproteine. I condrociti sono situati in cavità dette lacune e non sono
presenti vasi sanguigni; la cartilagine è circondata da un connettivo denso, detto pericondrio, che è necessario per
il suo mantenimento (è vascolarizzato) e per il suo accrescimento.
La cartilagine in base al tipo e alla quantità di bre viene classicata in:
Ialina, traslucida e resistente a trazione e compressione
11
Elastica, opaca e essibile
Fibrosa, una forma di transizione tra connettivo denso e cartilagine
3.4.1 La cartilagine ialina
La cartilagine ialina è formata da collagene II non raccolto in fasci e presenta proteoglicani specici (aggregano)
responsabili dell'idratazione.
Poichè molto resistente a compressione e trazione è il tipo di cdartilagine che forma le articolazioni e che sostiene
la trachea.
Di cartilagine ialina è costituito quasi tutto lo scheletro dell'embrione e del feto, nell'adulto rimane invece solo
sulle superci articolari, nelle cartilagini costali, nel naso, nella laringe, nella trachea e nei bronchi.
3.4.2 La cartilagine elastica
Si trova nel padiglione auricolare, nel meato e nella tuba uditiva e nell'epiglottide; di colore giallastro, è più opaca
della ialina, ha meno sostanza amorfa ma moltissime bre elastiche.
3.4.3 La cartilagine brosa
Principale componente dei dischi intervertebrali, forma grossi fasci brosi di collagene I immersi in una scarsa
matrice amorfa.
3.4.4 Istogenesi e accrescimento
Le cellule del connettivo embrionale si dierenziano inizialmente in condroblasti, i quali iniziano a produrre bre
collagene e proteoglicani; questa produzione è talmente copiosa che queste cellule si ritrovano ad essere isolate
singolarmente in lacune e vengono indotte a dierenziarsi diventando condrociti, più piccoli e meno attivi.
L'accrescimento avviene invece per due vie:
Interstiziale, in cui la cartilagine si espande dall'interno per divisione dei condrociti
Per apposizione, in cui viene aggiunta nuova cartilagine sulla supercie esterna tramite la dierenziazione di
cellule del pericondrio in condroblasti.
3.5
L'osso
Il tessuto osseo è un connettivo la cui matrice è mineralizzata e quindi resistente e duro; al contempo resistente e
leggero, l'osso non è un tessuto statico ma viene continuamente rinnovato e rimodellato.
Le funzioni dell'osso sono:
Impalcatura interna
Protezione e sostegno degli organi
Inserzione dei muscoli
Protezione degli elementi emopoietici
Deposito di calcio e fosforo
La popolazione cellulare tipica dell'osso è quella degli osteociti, che come nella cartilagine sono accolti in lacune
ossee dalle quali si irradiano i canalicoli ossei che si anastomizzano tra loro.
Si riconoscono due tipologie di osso: l'osso lamellare, compatto e resistente, e l'osso spugnoso, in cui l'organizzazione
interna è caotica.
A livello di formazione si parla di osso primario, cioè neoformato, accostabile anche se non sovrapponibile all'osso
spugnoso, e di osso secondario o lamellare; l'osso primario è destinato a essere sostituito dall'osso lamellare.
12
3.5.1 L'osso compatto
L'osso compatto è rivestito all'esterno dal periostio (che presenta le bre di Sharpey per fornire una presa salda ai
tendini) e all'interno dall'endostio. Dall'esterno all'interno sono presenti quattro sistemi lamellari:
Sistema circonferenziale esterno
Osteoni e lamelle interstiziali
Sistema circonferenziale interno
Gli osteoni, o sistemi di Havers, sono strutture cilindriche lamellari che al loro interno presentano il canale di Havers
lungo il quale scorrono i vasi sanguigni che entrano nell'osso grazie ai canali di Volkmann, ad esso paralleli.
Gli spazi tra gli osteoni sono riempiti dai sistemi di lamelle interstiziali.
3.5.2 L'ossicazione
Esistono due vie di ossicazione: l'ossicazione diretta, o intramembranosa, e quella indiretta, o endocondrale.
L'ossicazione diretta si ha quando cellule mesenchimali si dierenziano in osteoblasti, che iniziano a produrre
osteoide che viene poi mineralizzato. Gli osteoblasti isolati dierenziano in osteociti analogamente a quanto accade
nella cartilagine.
L'ossicazione indiretta prevede invece la formazione di un modello del futuro osseo in cartilagine; una volta
pronto i condociti centrali vanno incontro ad apoptosi e la matrice a loro esterna inizia a calcicare.
A questo
punto le cellule più interne del pericondrio si dierenziano in osteoblasti che formano per ossicazione diretta un
manicotto osseo che circonda la futura zona centrale e che viene penetrato da un'arteria che entra nella cartilagine
portando cellule osteoprogenitrici, condroclasti e cellule emopoietiche.
Al termine di un'ossicazione indiretta rimane cartilagine a livello della piastra di accrescimento e sulle diasi,
in modo da permettere una crescita dell'osso.
Le ossa lunghe crescono in spessore con un meccanismo duplice:
da un lato il periostio prolifera, dall'altro
l'endostio riassorbe l'osso, questo per mantenere costante il rapporto tra il diametro della cavità midollare e lo
spessore dell'osso.
La crescita in lunghezza avviene invece nella zona della piastra di crescita, che può essere suddivisa in cinque
strati:
Zona di riposo
Zona proliferativa
Zona di maturazione
Zona ipetroca
Zona di ossicazione
4
Il tessuto muscolare
Il tessuto muscolare, unico dotato di contrattilità, si divide in muscolo striato, a sua volta suddiviso in scheletrico
e cardiaco, e muscolo liscio.
4.1
Il muscolo scheletrico
Le funzioni del muscolo scheletrico sono i movimenti volontari dello scheletro, il mantenimento della postura, la
protezione degli organi interni e il controllo degli orizi.
Questo tipo di tessuto è costituito da cellule molto grandi (20-200
cilindriche, polinucleate.
13
µm
di diametro, no a 20 cm di lunghezza),
Queste cellule sono dei sincizi, derivano cioè dalla fusione di centinaia di mioblasti embrionali; i nuclei si trovano
in periferia, appena sotto la membrana cellulare, e all'interno di queste cellule sono presenti proteine contrattili
organizzate in miobrille.
Esistono tre involucri connettivali a proteggere il muscolo e a tenerlo unito:
Epimisio, avvolge l'intero muscolo
Perimisio, avvolge un fascio di bre
Endomisio, avvolge ogni singola bra
Quando all'estremità del muscolo questi tre involucri si fondono uno all'altro si crea un tendine.
4.1.1 Il sarcomero
Il sarcomero è l'unità funzionale contrattile del muscolo e presenta delle strutture dette bande e altre dette linee;
da un estremità alla linea centrale queste strutture sono:
Banda I, dove è presente sola actina
Linea Z, contenuta nella banda I, è la linea lungo la quale l'actina compie uno scarto
Banda A, dove è presente sia miosina che actina e per questo appare più scura
Linea M, contenuta nella banda A, dove è presente solo miosina
A livello tridimensionale ogni lamento di miosina è circondato da 6 lamenti di actina.
La contrazione avviene per scorrimento dei lamenti sottili sui lamenti spessi e si può suddividere in quattro
fasi:
1. La testa miosinica è bloccata su un lamento di actina
2. Una molecola di ATP si lega al retro della testa e questo ne riduce l'anità con l'actina permettendole di
muoversi lungo il lamento
3. Lo slittamento è di circa 5nm prima che l'ATP venga idrolizzato ad ADP, che però insieme al fosfato inorganico
rimane attaccato alla proteina
4. Il rilascio del fosfato inorganico genera una rapida contrazione come una vogata, durante la quale anche l'ADP
viene perduto e il ciclo può ricominciare
Il controllo della contrazione è legato ad altre due proteine: la tropomiosina e la troponina. La prima è una sorta
di binario lungo tutta l'actina, la seconda forma delle stazioni di controllo che legano il calcio.
Quando il calcio è legato alla troponina, la tropomiosina viene spostata in modo da permettere all'actina di
legare le teste della miosina: la contrazione può quindi avvenire.
Pertanto basse concentrazioni di
Ca++
impediscono la contrazione mentre alte concentrazioni la rendono pos-
sibile.
Il calcio viene introdotto grazie ad uno stimolo nervoso che genera una cascata di eventi:
1. L'assone propaga lo stimolo nervoso sotto forma di depolarizzazione
2. Viene rilasciato un neurotrasmettitore, l'acetilcolina, che lega un recettore sul sarcolemma e lo depolarizza
3. La depolarizzazione si trasmette ai tubuli T, invaginazioni del sarcolemma
4. La depolarizzazione dai tubuli T raggiunge le cisterne terminali del reticolo sarcoplasmatico, che rilascia ioni
Ca++ .
La tossina botulinica impedisce il rilascio di acetilcolina, il curaro si lega al recettore sul sarcolemma: entrambi
inducono paralisi accida.
La stricnina e il tetano invece impediscono il rilassamento del muscolo antagonista: inducono dunque paralisi
spastica.
14
4.2
Il muscolo cardiaco
Le cellule del muscolo cardiaco sono uninucleate e prendono il nome di miocardiociti. Hanno dimensioni di circa
20µm × 100µm.
Il nucleo è posizionato al centro della cellula e tra un miocardiocita e l'altro sono presente linee trasversali dette
strie scalariformi dove si trovano zonule adhaerens, desmosomi e gap junction.
Le principali dierenze con il muscolo scheletrico sono:
Tubuli T più grandi e numerosi
Reticolo sarcoplasmatico meno organizzato
Numerosi mitocondri
Il calcio entra dall'esterno della cellula prima che dall'interno
La contrazione è autonoma
L'innervazione ha solo funzione modulatoria
4.3
Il muscolo liscio
Si torva nella parete dei visceri cavi, quindi nel tratto gastrointestinale e genitourinario; è formato da cellule fusiformi
con nucleo centrale e con dimensioni tra i 20 e i 500
µm.
Le cellule del muscolo liscio contengono solo actina che a riposo non è organizzata in lamenti. Manca la striatura
trasversale ma la contrazione avviene come nel muscolo scheletrico per scorrimetno dei lamenti.
Le cellule del tessuto muscolare liscio sono in grado di trasmettersi tra loro l'impulso e si possono contrarsi anche
in assenza di uno stimolo nervoso, per esempio a causa di ormoni.
5
Il tessuto nervoso
Il sistema nervoso si occupa di fornire sensazioni sull'ambiente interno ed esterno, integrando le informazioni sensoriali, coordinando le attività motorie e regolando strutture ed apparati.
Anatomicamente viene suddiviso in:
SNC, comprendente encefalo e midollo spinale
SNP, tutto il rimanente sistema
Funzionalmente si riconosce invece
Sistema nervoso somatico, controllante le attività volontarie
Sistema nervoso autonomo, controllante le attività involontarie
Il tessuto nervoso è costituito dai neuroni e dalle cellule gliali; i neurono hanno le seguenti caratteristiche:
Numero compreso tra i 100 e i 1000 miliardi
Eccitabilità
Conduttività
Secernenti
Perenni
15
5.1
Il neurone
Un neurone è costituito da un corpo cellulare, da dei dentriti e da un assone.
Il corpo cellulare, o pirenoforo, ha una morfologia variabile e dimensioni comprese tra i 5 e i 150
µm.
Il nucleo,
voluminoso e centrale è chiaro e il nucleolo è sede di elevata attività di sintesi ribosomiale.
Il citoplasma del pirenoforo contiene numerosi mitocondri e un apparato del Golgi molto sviluppato; sono presenti
inoltre molti ribosomi liberi.
I dendriti sono in genere multipli ed emergono da vari punti del corpo cellulare; sono brevi e ramicano nelle
vicinanze del pirenoforo. Contengono tutti gli organuli cellulari ad eccezione del Golgi e sono funzionalmente delle
espansioni del corpo cellulare generalmente deputate a ricevere gli stimoli.
L'assone è presente in tutti i neuroni con lunghezze no ad un metro, origina da una protrusione detto cono di
emergenza e di solito emette i suoi rami distalmente al pirenoforo nel territorio di innervazione.
L'impulso nervoso è un segnale di natura elettrica e il neurone è in grado di veicolarlo.
La supercie interna della membrana cellulare neuronale è leggermente più negativa rispetto a quella esterna:
si ha un potenziale di membrana a riposo di
−70mV .
In risposta ad uno stimolo adeguato si ha un inversione del potenziale di riposo e la generazione di un potenziale
d'azione: si passa a un potenziale di
5.2
+30mV .
La guaina mielinica
La guaina mielinica è formata dalle cellule di Schwann (SNP) o da cellule dette oligodentrociti (SNC) disposte a
formare una spirale di membrane cellulari avvolte intorno all'assone.
Questa disposizione isola il neurone a livello elettrico, in questo modo si passa da una conduzione lineare ad una
a tratti in cui è necessario ripolarizzare solo i tratti scoperti dalla guaina, cioè i nodi di Ranvier.
La velocità di conduzione dell'impulso dipende infatti dal diametro delle bre (più è elevato più è veloce la
trasmissione) e dalla presenza o l'assenza di mielina: la sclerosi multipla è una patologia legata alla sua distruzione
che quindi genera danni irreversibili al sistema nervoso.
5.3
La sinapsi
La porzione terminale di un assone presenta un rigonamento, il bottone sinaptico, che costituisce metà di una
sinapsi: l'altra metà è la porzione di membrana cellulare dell'assone a valle con i suoi recettori per i neurotrasmettitori. Tra le due porzioni vi è la fessura sinaptica che ha una dimensione di circa 20-30 nm.
Quando un impulso nervoso raggiunge un neurone, esso è veicolato a livello della singola cellula per via elettrica
ma viene trasmesso per via chimica, è questa infatti la funzione della sinapsi.
In risposta al potenziale d'azione le sinapsi secernono molecole dette neurotrasmettitori che si legano alla membrana della cellula a valle, che può uscirne:
Depolarizzata: stimolo eccitatorio, legato a neurotrasmettitori quali l'acetilcolina
Iperpolarizzata: stimolo inibitorio, legato a neurotrasmettitori quali GABA
Importante notare che il segnale attraversa la sinapsi in una sola direzione, sempre.
5.4
Cellule gliali
Le cellule gliali sono un gruppo eterogeneo di cellule 10 volte più numeroso dei neuroni che presenta funzioni diverse:
Funzioni di sostegno, nutrimento e riparazione: astrociti
Funzioni strutturali: cellule di Schwann, oligodentrociti
Funzioni difensive: microglia
16
Part II
Embriologia
6
Introduzione alla riproduzione
La riproduzione è una proprietà fondamentale degli organismi viventi che assicura la continuità della vita, l'adattamento
e l'evoluzione.
Le due uniche modalità di riproduzione sono:
Riproduzione asessuata (agamica): semplice e veloce, conferisce la forza del numero ma ha come conseguenza
l'uniformità genetica e quindi una minore probabilità di adattarsi ai cambiamenti
Riproduzione sessuata: Lenta e dispendiosa, conferisce la capacità di adattarsi con grande successo ai cambiamenti ed è l'unica forma di riproduzione negli animali superiori e nell'uomo
6.1
La determinazione del sesso
All'inizio del 1900 si dimostrò come il sesso dell'individuo fosse legato ai cromosomi, in particolare fu dimostrato
che nell'uomo il maschio è eterogametico, presenta cioè due cromosomi sessuali diversi XY, mentre la femmina è
omogametica, cioè presenta due cromosomi sessuali uguali XX.
L'acquisizione di caratteristiche maschili o femminili prevede alcune tappe:
Lo sviluppo di vie genitali e genitali interni è il sesso genitale
Lo sviluppo dei caratteri sessuali secondari e dei genitali esterni è il sesso fenotipico, cioè visibile
Esistono però individui che presentano un sesso genetico che diverge dal sesso fenotipico: si tratta degli individui
pseudoermafroditi.
Questo si spiega poichè la natura realizza spontaneamente il fenotipo femminile in assenza di qualcosa che sia
in grado di modicare questo comportamento.
I testicoli nel maschio sono dunque chiamati a una duplice azione:
Realizzazione attiva del fenotipo
Soppressione del fenotipo
♀
♂
con la secrezione di ormoni
Due ormoni sono legati a queste attività: l'ormone per la realizzazione attiva è il testosterone, l'ormone per la
soppressione il fattore antimullerano (AMH, Anti-Mulleran Hormone).
Si può concludere che un individuo diventerà maschio se rispetta due condizioni:
1. Vengono prodotti testosterone e AMH
2. L'individuo è sensibile a questi due ormoni
Con lo studio delle malattie legate alla presenza di cromosomi sessuali extra si è inoltre capito come sia tutto
attorno al cromosoma Y il lavoro di dierenziamento sessuale: la presenza di Y determina mascolinità, mentre la
sua assenza determina femminilità.
Si iniziò a metà del secolo scorso a cercare cosa scatenasse la mascolinità nel cromosoma Y: intorno al 1990
venne scoperto il gene SRY, sex region Y, che da solo attiva una cascata di eventi che portano allo sviluppo di un
individuo di sesso maschile.
17
6.2
La gametogenesi clinica
La gametogenesi è il processo che porta alla formazione dei gameti che poi si uniranno a formare lo zigote. Questo
processo spesso va incontro ad errori che generano patologie genetiche.
Uno degli errori comuni è la non disgiunzione tra i cromosomi, che porta ad uno zigote con cromosomi sovrannumerari, cioè in condizione di aneuploidia.
Lo sbilanciamento tra i geni crea un fenotipo anomalo o spesso la morte, tanto che sono solo tre le trisomie
autosomiche che possono portare a termine la gravidanza:
Trisomia 21, o sindrome di Down
Trisomia 13, o sindrome di Patau
Trisomia 18, o sindrome di Edwards
Meno letali sono le aneuploidie dei cromosomi sessuali, esistono individuo XXY, XYY, XXX, XXXX e XXXXX.
La monosomia è invece sempre incompatibile con la vita tranne il caso XO, sindrome di Turner
7
L'ovogenesi
L'ovogenesi, cioè il processo di formazione dei gameti femminili, o cellule uovo, è caratterizzato da una scansione
temporale a tre fasi:
1. Fase di inizio, durante lo sviluppo embrionale
2. Fase di arresto, durante lo sviluppo embrionale
3. Fase di ripresa, dalla pubertà alla menopausa
7.1
La fase di inizio e la fase di arresto
Durante la vita embrionale, in particolare dall'XI settimana al 7° mese di vita intrauterina, le cellule germinali
primordiali dette ovogoni sono indotte a trasformarsi in ovociti primari.
Questo processo è inevitabile per tutti gli ovogoni: o entrano in meiosi o muoiono.
Gli ovociti primari vengono bloccati in fase meiotica, precisamente nella profase, e questo stop può durare anche
50 anni.
In questa fase il follicolo primordiale è l'unità funzionale dell'ovaio ed è formato dall'ovocita I e dalle cellule
piatte che lo circondano.
Intorno al 5° mese di vita intrauterina sono presenti nell'organismo femminile
riusciranno a diventare follicoli primordiali sono in realtà solo
1 − 2 ∗ 10
6
6 − 7 ∗ 106
ovogoni ma quelli che
ed è questo il numero di unità funzionali
presenti alla nascita.
Poichè tutte le cellule progenitrici sono già presenti alla nascita della donna, non esistono cioè cellule staminali,
la loro distruzione porta inevitabilmente alla sterilità.
7.2
La fase di ripresa
La fase di ripresa inizia con la pubertà e interessa diversi follicoli primordiali ogni giorno. Di questi solo un ovocita
I con il suo follicolo completa lo sviluppo, l'ovulazione è quindi un processo ciclico.
Il ciclo ovarico è la nestra di tempo compresa tra due successive ovulazioni ma è più semplice considerarlo il
periodo tra due mestruazioni successive in modo da dividerlo in due fasi:
Fase follicolare, dominata dagli estrogeni follicolari, termina con l'ovulazione
Fase luteinica, segue l'ovulazione ed è dominata dai progestinici secreti dal corpo luteo
In generale il ciclo ovarico è legato a una regolazione ormonale di tipo feedback negativo, in cui si ha l'azione di
quattro ormoni divisi in coppie: progesterone ed estrogeni inibiscono ipotalamo ed iposi nella produzione di FSH
ed LH.
18
7.2.1 La fase follicolare
La fase follicolare è la fase in cui diversi follicoli primordiali iniziano la maturazione ma al suo termine uno sono di
essi avrà completato la il processo.
Questa fase viene suddivisa in tre sottofasi:
1. Fase preantrale, della durata di 3-5 giorni
2. Fase antrale, della durata di 8-12 giorni
3. Fase preovulatoria, della durata di 37 ore
7.2.1.1 La fase preantrale
In questa fase alcuni follicoli primordiali, quindi dotati di ovocita I, entrano nella
prima fase di accrescimento che li porta a diventare follicoli primari; questo passaggio prevede l'aumento di dimensioni dell'ovocita I e la formazione da parte delle cellule follicolari della granulosa.
Le cellule dello stroma ovarico formano la teca, suddivisa in teca interna, vascolarizzata, e teca esterna, brosa.
Le cellule della granulosa formano la zona pellucida, uno strato di glicoproteine (ZP1, ZP2 e ZP3) che circonda
l'ovocita e ha un ruolo fondamentale nel riconoscimento uovo-spermatozoo.
Oltre alle modicazioni morfologiche appaiono nella fase preantrale anche modicazioni funzionali, in particolare
compaiono i recettori per le gonadotropine (FSH e LH) che sono fondamentali per il passaggio alla fase successiva
che, a dierenza di questa, è totalmente dipendente dagli ormoni.
7.2.1.2 La fase antrale
I follicoli primari entrano in questa fase solamente se vi è un'adeguata concentrazione
di FSH e LH nel sangue e se essi stessi presentano un numero di recettori suciente: se una o entrambe queste
condizioni non sono esaudite il follicolo degenera e si ha una condizione di atresia.
Il numero di follicoli che raggiunge la fase antrale è solitamente tra le 15 e le 20 unità.
Durante questa fase si ha il passaggio da follicolo primario a follicolo secondario, passaggio scandito da modiche
strutturali e funzionali.
La più evidente trasformazione strutturale è la formazione di un antro all'interno del follicolo, e una divisione
tra le cellule della granulosa e l'ovocita (rimane un piccolo accumulo detto cumulo ooforo).
A questo punto il follicolo diventa una ghiandola endocrina, in cui la teca interna con recettori per LH produce
androgeni mentre la granulosa, che possiede recettori per FSH, produce estrogeni a partire dagli androgeni.
In questa fase si ha una competizione tra i follicoli delle due ovaie e il follicolo che acquista più recettori degli
altri cresce più rapidamente sottraendo FSH ed estrogeni agli altri in un circolo vizioso che porta ad una crescita
esplosiva: si parla ora di follicolo dominante, che passerà alla fase successiva.
7.2.1.3 La fase preovulatoria
La fase preovulatoria prevede il passaggio da follicolo secondario a follicolo
terziario in condizioni di alte concentrazioni di FSH ed LH.
Anche l'ovocita passa dalla condizione di ovocita I a quella di ovocita II completando dunque la I divisione
meiotica; la seconda cellula prodotta dalla divisione eredita pochissimo citoplasma ed è condannata a degenerare:
si denisce globulo polare.
L'ovocita II a questo punto viene ribloccato in meiosi, ma questa volta non più alla profase ma alla metafase,
in attesa della fecondazione.
7.3
L'ovulazione
Quando il follicolo terziario è vicino alla supercie dell'ovaio si forma un rigonamento e la parete ovarica diventa
avascolare; la membrana propria del follicolo tra teca e granulosa viene rotta, quest'ultima viene vascolarizzata e
inizia così la trasformazione in corpo luteo.
L'ovocita II e il cumulo ooforo intanto vengono espulsi dall'ovaio che viene catturato dalle mbrie dell'ovidotto.
19
7.4
La fase luteinica
Le cellule della granulosa dopo l'ovulazione si trasformano in cellule luteiniche che cessano di dividersi e diventano
ipertroche accumulando grandi quantità di lipidi e di luteina, un pigmento giallo.
Il corpo luteo ha le funzioni di una ghiandola endocrina e produce soprattutto progesterone ma in quantità
minori anche estrogeni.
Questa struttura è programmata per autodistruggersi dopo 14 giorni secondo un processo noto come luteolisi;
se questo processo ha luogo crolla la produzione di progesterone e di estrogeni, liberando così l'attività dell'iposi
che torna a secernere FSH e LH in modo da riprendere un nuovo ciclo ovarico.
Il corpo luteo può essere salvato da HCG, la gonadotropina corionica umana, che viene prodotta in caso di
fecondazione: in questa situazione continua la produzione di progesterone ed estrogeni e quindi non è possibile
intraprendere un nuovo ciclo ovarico.
La pillola anticoncezionale funziona come un corpo luteo articiale, impendendo l'inizio di un nuovo ciclo ovarico.
7.5
Gli altri cicli femminili
Ad accompagnamento del ciclo ovarico, l'organismo femminile presenta altri cicli periodici che hanno ripercussioni
importanti per la fecondazione.
Il ciclo più evidente è quello uterino, e per comprenderlo è necessario analizzare la struttura dell'utero.
L'utero è un organo muscolare a forma di pera lungo 7 cm, largo 4 e spesso 2,5. Il corpo è la parte espansa dove
si aprono le tube, il fondo è la parte superiore allo sbocco, mentre la cervice è la parte circolare che si apre nella
vagina.
Si riconoscono due strati in questo organo: l'endometrio, che è una mucosa, e il miometrio, che è uno strato
muscolare liscio.
Lo strato di endometrio è a sua volta suddiviso in uno strato funzionale e uno strato basale: il primo viene
sfaldato ad ogni ciclo mentre il secondo è deputato alla ricostruzione ciclica di ciò che viene perso.
Il ciclo uterino endometrico è quindi diviso in due fasi:
1. Fase proliferativa, sovrapposta a quella follicolare, termina con l'ovulazione: si ha la proliferazione dello strato
basale e la rigenerazione di quello funzionale
2. Fase secretiva, sovrapposta a quella luteinica, termina con la mestruazione: si ha l'aumento di spessore dello
strato funzionale in preparazione alla mestruazione.
Esiste anche un ciclo uterino miometrico in cui cambiano la frequenza e l'intensità contrattiva di questo muscolo:
durante l'ovulazione si hanno contrazioni ad alta frequenza (200/h) e bassa ampiezza mentre durante la mestruazione
se ne hanno a bassa frequenza (30/h) ma grande ampiezza.
Particolarmente rilevante a livello di fecondazione è il ciclo cervicale:
le ghiandole nella zona della cervice
secernono un muco le cui caratteristiche cambiano a seconda del momento del ciclo e ostacolano o favoriscono la
fecondazione.
8
La spermatogenesi
La spermatogenesi è il processo di formazione degli spermatozoi: a dierenza dell'ovogenesi, non inizia in età fetale,
ma con la pubertà.
Con la pubertà a livello dei testicoli si hanno dei cambiamenti strutturali che rendono possibile il processo,
cambiamenti che sono:
I cordoni seminiferi originano i tubuli seminiferi
Le cellule interstiziali si dierenziano in cellule del Leydig
Le cellule sostentacolari si dierenziano in cellule del Sertoli
Gli spermatogoni ricominciano a dividersi per mitosi diventando spermatogoni di tipo A
20
L'intero processo di spermatogenesi è intimamente legato alla morfologia delle struttura che lo interessano, in
particolare quella dei tubuli seminiferi.
I tubuli sono formati da due popolazioni cellulari: l'epitelio seminifero con le cellule spermatogene e le cellule
sostentacolari del Sertoli.
La parete dei tubuli è formata da una membrana basale cui sono collegate delle cellule mioidi di tipo muscolare:
questa parete separa il compartimento tubulare dal compartimento interstiziale creando così due ambienti totalmente
diversi.
Fuori dalla parete rimangono le cellule del Leydig, che nell'uomo sono deputate alla produzione di ormoni
androgeni.
Le cellule del Sertoli, interne ai tubuli seminiferi, concorrono a formare la barriera emato-testicolare attraverso
un sistema di giunzioni occludenti; questo suddivide il tubulo in due compartimenti, un compartimento basale tra
parete e giunzioni e un compartimento adluminale.
La barriera ematotesticolare separa cellule in momenti diversi della loro vita: nel compartimento basale si esegue
la mitosi, in quello adluminale la meiosi.
Il processo della spermatogenesi avviene in tre fasi di durata costante e caratteristica per ogni specie:
1. Fase di proliferazione: 16 giorni nell'uomo
2. Fase di meiosi: 24 giorni nell'uomo
3. Fase di spermiogenesi: 24 giorni nell'uomo
8.1
La fase di proliferazione
Durante la pubertà gli spermatogoni primitivi si trasformano in spermatogoni di tipo A, scuri e staminali, che sono
in grado di riprendere la spermatogenesi; quando gli spermatogoni di tipo A diventano di tipo A1 (si riconoscono
poichè di colore chiaro), inizia la prima mitosi.
Una volta iniziato il processo, una sequenza ssa di mitosi (nel caso dell'uomo sono 6) porta alla formazione
degli spermatociti primari: poichè nell'umano vi sono 6 mitosi, da ogni spermatogone si ricavano 64 cellule o meno.
In ogni clone di cellule derivanti da uno spermatogone intermedio non si divide il citoplasma ma restano dei
ponti necessari a garantire la sincronia dello sviluppo.
8.2
La fase di meiosi
Gli spermatociti primari sono ancora diploidi, ma alla ne di questa fase verranno prodotti spermatidi aplodi:
da ogni spermatocita primario che vi entra vengono dunque generato quattro spermatidi, sempre dotati di ponti
citoplasmatici per la sincronia.
8.3
La fase di spermiogenesi
In questa fase si passa da una cellula poco caratteristica a uno spermatozoo vero e proprio grazie a un drastico
rimodellamento cellulare.
Il nucelo diventa ora eterocromatico e dall'apparato di Golgi si crea l'acrosoma, un organo di penetrazione
paragonabile ad un lisosoma gigante che si pone in testa al futuro spermatozoo.
In totale uno spermatozoo sarà lungo circa 60
µm,
di cui 4-5 rappresentati dalla testa: la restante porzione è la
coda, suddivisa in
collo
pezzo intermedio
segmento principale
segmento terminale
21
8.4
Organizzazione spaziotemporale della spermiogenesi
Il processo di spermiogenesi è legato all'attività delle cellule del Sertoli che spingono verso il lume del tubulo le
cellule con il procedere della loro maturazione.
La traiettoria seguita dalle cellule nella spinta è quella del raggio del tubulo, in totale il tempo tra due spermatogenesi successive è costante per ogni specie e rappresentato da
1/4
del periodo totale che per l'uomo è quindi
pari a 16 giorni.
In ogni sezione frontale del tubulo dunque gli strati concentrici sono popolati da cellule allo stesso stadio di
maturazione, sincronia realizzata grazie ai ponti citoplasmatici.
Per garantire una produzione continua di spermatozoi le sezioni del tubulo non iniziano i cicli allo stesso tempo
ma secondo scadenze dettate dalla cosiddetta onda dell'epitelio seminifero: iniziano prima le porzioni periferiche e
via via no ad arrivare alla porzione centrale.
8.5
Controllo ormonale
Il controllo ormonale della spermiogenesi sfrutta gli stessi ormoni che controllano l'ovogenesi: LH stimola le cellule
del Leydig a produrre testosterone mentre FSH stimola l'attività delle cellule del Sertoli che dirigono di fatto l'intero
processo.
22
9
Le prime quattro settimane di sviluppo
Giorno
Evento
0
Fecondazione
1
Perdita corona radiata, prima divisione
Neuro
Cardio
2
3
Formazione della morula
4
Formazione della blastocisti
5
Schiusa della blastocisti
6
7
8
Formazione del disco embrionale
Formazione del sacco vitellino primario
9
10
Formazione del mesoderma extraembrionale
11
12
Formazione del sacco vitellino denitivo
13
Formazione del corion
I circ. utero-placentare
14
15
Inizio della gastrulazione
16
Membrana buccofaringea e cloacale distinte
17
Migrazione del mesoderma
Canale cordale
Placca neurale
18
19
Placca cordale
Separazione del mesoderma laterale
Canale dorsale
Doccia neurale
20
21
Formazione dei somiti
Tubo neurale
Inizio chiusura tubo
22
Battito cardiaco
23
24
Chiusura neuroporo ant.
Circ. sanguigna
25
26
Chiusura neuroporo post.
27
28
10
La prima settimana di sviluppo
Una volta avvenuta la fecondazione vi sono quattro settimane ricche di eventi che fanno diventare un'unica cellula,
lo zigote, un embrione umano riconoscibile.
La prima divisione dello zigote si ha dopo 24-30 ore dalla fecondazione: si passa dunque a uno stadio di due
cellule e connessa a questa divisione è la perdita della corona radiata.
Inizialmente lo zigote è molto più grande di una cellula umana tipica, pertanto si ha un processo di segmentazione
che porta ad aumentare il numero di cellule senza aumentare le dimensioni della struttura: il risultato nale è la
morula, un'organizzazione compatta che è riconoscibile a partire dal terzo giorno e che è formata da un numero
variabile tra 8 e 18 cellule.
Durante il quarto giorno la morula si trasforma in blastocisti e si ha la prima dierenziazione cellulare:
formano due gruppi cellulari di cui uno già destinato a non far parte dell'embrione.
La blastocisti, è dunque formata da:
23
si
All'esterno il trofoblasto, massa cellulare destinata a formare i tessuti extraembrionali
All'interno il nodo embrionale, destinato a formare sia l'embrione che alcuni tessuti extraembrionali.
Durante il quinto giorno si ha la schiusa della blastocisti: la zona pellucida viene digerita ad opera di enzimi secreti
dal trofoblasto e questo permette l'impianto, evento cruciale che si colloca a cavallo tra la prima e la seconda
settimana di sviluppo.
La fase di impianto è delicatissima ed è quella in cui la blastocisti penetra progressivamente la parete uterina
venendo a diretto contatto con le cellule materne.
Inizia tra il sesto ed il settimo giorno e si divide in due fasi: un aggancio a opera dei microvilli della blastocisti
e una penetrazione vera e propria.
L'inizio della fase penetrativa impone una seconda dierenziazione cellulare: le cellule del trofoblasto a contatto
con la parete endometrica si trasformano in sinciziotrofoblasto, formato da cellule polinucleate, mentre le altre
cellule diventano citotrofoblasto.
L'impianto avviene a metà della fase luteinica del ciclo ovarico, periodo di dominanza del progesterone: la pillola
del giorno dopo è una dose massiccia di estrogeni che inverte questa dominanza e impedisce alla blastocisti di iniziare
l'opera.
Nel nodo embrionale vi è anche un'altra dierenziazione: si forma uno strato detto ipoblasto.
Alla ne della prima settimana vi sono quindi quattro strati diversi nell'embrione:
1. Citotrofoblasto
2. Sinciziotrofoblasto
3. Ipoblasto
4. Nodo embrionale o massa cellulare interna
11
La seconda settimana di sviluppo
La seconda settimana è caratterizzata da due processi principali: la conclusione dell'impianto e la formazione di
disco embrionale, amnios e sacco vitellino.
La prima struttura a formarsi è il disco embrionale, a cavallo tra l'ottavo e il nono giorno; tra sinciziotrofoblasto
e nodo embrionale si formano cavità piene di liquido che conuiscono tra loro a formare la cavità dell'amnios che
verrà rivestita di amnioblasti ad opera dell'epiblasto. Si forma così un dischetto formato da epiblasto ed ipoblasto
compreso tra due cavità: sul versante uterino la cavità amniotica e sull'altro il blastocele.
Più o meno contemporaneamente alla formazione del disco embrionale si ha la formazione del sacco vitellino
primario: il blastocele viene rivestito ad opera dell'ipoblasto con una membrana, detta membrana di Heuser e a
questa struttura si da appunto il nome di sacco vitellino primario.
Tra il decimo e l'undicesimo giorno l'impianto è di fatto completato e nel sinciziotrofoblasto iniziano a formarsi lacune piene di sangue e uidi e si forma inoltre il mesoderma extraembrionale, cioè lo strato compreso tra
sinciziotrofoblasto e citotrofoblasto.
Al dodicesimo giorno di sviluppo le lacune si mettono in comunicazione tra di loro formando un sistema lacunare
che è la prima circolazione utero-placentare.
L'ipoblasto in questo stesso giorno prolifera formando il rivestimento denitivo del sacco vitellino ricoprendo la
membrana di Hauser.
L'ultima struttura a formarsi durante la seconda settimana di sviluppo è il corion, al tredicesimo giorno
Le cavità del mesoderma extraembrionale, formatesi durante il dodicesimo giorno, qui conuiscono formando il
celoma extraembrionale che circonda l'embrione in ogni sua parte escluso il peduncolo di attacco.
Questo celoma divide il mesoderma embrionale in due strati:
1. Somatopleura extraembrionale, che si accolla internamente ad amnios e citotrofoblasto
2. Splancnopleura extraembrionale, che si accolla al sacco vitellino
24
Il corion è il risultato dell'insieme di tre tessuti:
1. Citotrofoblasto
2. Somatopleura
3. Sinciziotrofoblasto
Alla ne della quarta settimana si hanno dunque le seguenti strutture:
Corion, formato da citotrofoblasto,sinciziotrofoblasto e somatopleura
Epiblasto, che darà origine all'embrione
Cavità amniotica
Sacco vitellino secondario
12
La terza settimana di sviluppo
La terza settimana presenta numerosi processi fondamentali:
1. Formazione dei tre foglietti germinativi e annessa gastrulazione
2. Inizio della neurulazione
3. Inizio dello sviluppo cardiocircolatorio
Il primo evento della terza settimana è la gastrulazione, che inizia al 15° giorno: cellule dell'epiblasto proliferano e
migrano dalla periferia verso l'asse centrale formando un ispessimento detto linea primitiva; l'ispessimento anteriore
della linea primitiva è detto nodo di Hensen.
La linea permette di denire le dimensioni dell'embrione, in particolare l'asse cefalo-caudale, le superci dorsale
e ventrale, e i lati destro e sinistro dell'embrione.
A partire dalla formazione della linea si hanno dei movimenti che portano alla formazione dei tre foglietti
embrionali:
L'endoderma si forma quando l'epiblasto si invagina attraverso la linea primitiva spostando l'ipoblasto
Il mesoderma si forma per seconda migrazione dell'epiblasto e quindi si trova sopra l'endoderma ma sotto
l'epiblasto
L'ectoderma si forma inne con la terza migrazione dell'epiblasto e quindi è lo strato superiore
Al 16° giorno l'asse cefalo-caudale diventa evidente poichè sono ora visibili la membrana cloacale posta al termine
della linea primitiva, e la membrana buccofaringea, posta all'altra estremità.
12.1
La notocorda
Il nodo di Hensen assume ora un ruolo fondamentale poichè da questo punto in avanti si ha il processo cefalico o
notocordale che darà origine alla notocorda.
La formazione della notocorda è un processo a tre stadi che occupa i giorni da 17 a 19: si ha la fase del canale
cordale, quella della placca e inne la fase della corda dorsale.
La prima fase si ha quando il processo cefalico si sposta in avanti e si cavita, la seconda si ha quando si fonde
all'endoderma formando il canale neuroenterico tra amnios e sacco vitellino, la terza ed ultima fase si ha quando la
placca cordale si ripiega a formare un cordone pieno: la notocorda.
La notocorda è una struttura fondamentale per lo sviluppo, in quanto serve come struttura assile per lo sviluppo
dello scheletro e in particolare della spina dorsale ma soprattutto induce la formazione del tubo neurale. Nell'adulto
i resti della notocorda sono i nuclei polposi dei dischi intervertebrali.
25
12.2
La neurulazione
Come il processo che forma la notocorda, anche la neurulazione attraversa tre stadi: si parla di placca neurale, di
doccia neurale e inne di tubo neurale.
Inizialmente, intorno al 17° giorno, si ha una placca cellulare posta sopra la notocorda, ma al 19° questa placca
assume una forma a ferro di cavallo e viene denita doccia, cioè incavo.
L'ultima fase è la chiusura delle estremità del ferro di cavallo con formazione di un tubo posto direttamente
sopra alla notocorda.
12.3
La suddivisione del mesoderma
Intorno al 19° giorno il mesoderma si presenta come suddiviso in cinque aree organoformative:
1. Mesoderma cordale, associato alla notocorda
2. Mesoderma parassiale, posto lateralmente al tubo neurale
3. Mesoderma intermedio, posto tra quello parassiale e le estremità embrionali
4. Somatopleura, posta sotto l'ectoderma
5. Splancnopleura, posta sopra l'endoderma
26
Figure 1: La suddivisione del mesoderma
Il mesoderma parassiale si ispessisce e comincia ad organizzarsi in raggruppamenti cellulari detti somitomeri,
molti dei quali si frammentano in somiti.
La formazione comincia alla ne della terza settimana e alla ne della quarta si avranno in totale 7 paia di
somitomeri e 37 coppie di somiti: 4 occipitali, 8 cervicali, 12 toracici, 5 lombari, 5 sacrali e 3 coccigei.
12.4
La circolazione primitiva
Alla seconda settimana l'embrione è nutrito passivamente dalle lacune del sinciziotrofoblasto ma ora tale sistema
non è più suciente: sangue, vasi e cuore derivano dal mesoderma splancnico e il processo inizia tra il 17° e il 18°
giorno.
Cellule mesenchimali del saccco vitellino si trasformano in isole sanguigne che originano in periferia degli angioblasti, che daranno vitra ai vasi, e all'interno degli emoblasti, cioè eritrociti primitivi.
13
La quarta settimana di sviluppo
Alla ne della terza settimana l'embrione è piatto e formato dai tre foglietti germinativi: la lunghezza è di circa
4mm e ha un aspetto simile a quello di tutti i vertebrati che hanno completato la gastrulazione.
27
Durante la IV settimana vi sono una serie di movimenti e di ripiegamenti che hanno come risultati:
1. Cambiamento della forma esterna dell'embrione
2. Delimitazione del corpo
3. Cambiamento di dimensioni e posizioni delle parti interne
4. Chiusura del tubo neurale
5. Evoluzione delle cinque aree organo-formative
L'espansione laterale dell'amnios è l'evento che scatena la delimitazione laterale del corpo embrionale: la cavità
espandendosi scende ai lati e forma le pieghe laterali avvolgendo l'embrione e trasformandolo in un tubo isolato dai
suoi annessi se non per un peduncolo.
Nella regione anteriore e in quella posteriore l'avvolgimento dell'amnios e la fusione delle pieghe laterali è
completa e viene data origine ad un intestino anteriore e uno posteriore.
13.1
Evoluzione dell'ectoderma
Già a partire dalla III settimana l'ectoderma si suddivide in due aree organo-formative: l'ectoderma nervoso, più
profondo, e l'ectoderma epidermico, più superciale.
L'ectoderma a destino nervoso chiude il tubo neurale a partire dal 21° giorno iniziando dalla zona centrale
dell'embrione e proseguendo verso le due estremità: queste si chiudono mediamente al 24° giorno (anteriore) e al
26° giorno (posteriore).
Il tubo neurale, ora completamente chiuso, diventa il primordio del sistema nervoso centrale: l'encefalo e il
midollo spinale.
Sopra ad esso vi è la cresta neurale, una popolazione di cellule destinate a migrare in regioni
diverse del corpo: queste si dierenzieranno in neuroni e cellule gliali, ma anche in melanociti e in derma della
faccia e del collo.
Riassumendo dunque l'ectoderma durante la IV settimana si suddivide in tre aree organoformative:
1. Ectoderma di rivestimento: epiteli, ghiandole e annessi cutanei
2. Tubo neurale: cervello, midollo spinale, nervo ottico, retina, parte dell'iposi
3. Cresta neurale: neuroni, cellule gliali, meningi, derma, muscoli dell'iride, cartilagini facciali, melanociti
13.2
Evoluzione dell'endoderma
Durante la IV settimana con la formazione delle pieghe laterali e longitudinali l'endoderma va a formare tre regioni:
l'intestino anteriore, quello medio e quello posteriore; ciascuna di queste porzioni non va a formare solo tratti
dell'apparato digerente ma anche altri organi.
1. Intestino anteriore: itestino branchiale, apparato respiratorio, esofago, stomaco, fegato, pancreas, cistifellea,
duodeno superiore
2. Intestino medio: duodeno inferiore, intestino tenue e crasso, colon ascendente e i primi due terzi del trasverso
3. Intestino posteriore: ultimo terzo trasverso, colon discendente e sigmoide, retto, vescica urinaria e urentra
primitiva
L'intestino branchiale, formato dall'intestino anteriore, originerà bocca, naso, orecchie, collo e faringe, tutte strutture
che passano prima per la forma di tasche branchiali.
28
13.3
Evoluzione del mesoderma
Alla ne della terza settimana il mesoderma è diviso in cinque aree organo-formative: cordale, intermedio, parassiale,
splancnopleura e somatopleura.
Il mesoderma parassiale forma i somitomeri che all'inizio della IV settimana sono 49 di cui 7 non frammentati
e gli altri divisi in somiti.
I somiti in questa fase embrionale si cavitano e si frammentano, dividendosi in due aree:
dermomiotomo e
sclerotomo.
Lo sclerotomo forma l'abbozzo di vertebre, scapole e coste, il dermiotomo viene invece suddiviso ulteriormente
in dermotomo, che origina il derma, e il miotomo, che origina muscoli delle vertebre, del tronco e degli arti.
29