Modulo di Virologia (canali 1 e 2), AA 2016/2017
Prof. Milena Grossi
Corso integrato Microbiologia e Virologia
Dal 11/01/2017 al 20/01/2017
Orario lezioni:
Tutti i giorni
Lun-Mer-Ven 11.00-13.00 Aula Sergi
Mar-Gio 9.00-11.00 Aula IV via dei Marsi
78
Modulo di Virologia (canali 1 e 2)
Corso integrato Microbiologia e Virologia
Libri di testo:
G. Dehò, E. Galli
“Biologia dei microrganismi”, Cap. 16. Casa Editrice Ambrosiana
2014. Per una visione generale
E
Alan J. Cann
“Elementi di Virologia Molecolare”
ed. 2006, Casa Editrice Ambrosiana
Per approfondire alcuni argomenti
Modalità esame del Modulo di Virologia
• L’esame è orale e sarà possibile sostenerlo nelle date di
appello pubblicate su Infostud dai titolari del corso integrato
Microbiologia e Virologia, prof Bianca Colonna per il canale 1
(A-H) e prof Maria Lina Bernardini per il canale 2 (I-Z).
• Per chi desidera la modalità scritta sono previsti 3 esoneri :
27 Gennaio 2017 e 17 Febbraio 2017
• Per partecipare agli esoneri è necessario prenotarsi sul
elearning2.uniroma1.it alla pagina relativa al modulo di Virologia
del corso integrato Microbiologia e Virologia.
Programma Modulo di Virologia
(canali 1 e 2) AA 2016/2017
Corso integrato Microbiologia e Virologia
•
•
•
•
•
•
•
•
•
•
•
Caratteri generali sui virus: composizione dei virioni; struttura del virione: il
capside (simmetria icosaedrica ed elicoidale), l’involucro pericapsidico
(envelope);
Criteri di classificazione dei virus e suddivisione in classi di replicazione: lo
schema di Baltimore.
I genomi virali.
Ciclo replicativo.
Retrotrascrizione.
Regolazione dell’espressione e replicazione dei genomi virali a DNA.
Regolazione dell’espressione e replicazione dei genomi virali a RNA.
Interazioni genetiche e non genetiche tra virus
Modelli di infezione e Interazione virus-ospite
Prevenzione e terapia delle infezioni virali.
Metodi di saggio: colture di cellule animali e controllo della moltiplicazione
cellulare, quantificazione delle particelle virali, misurazione delle unità infettive
virali, identificazione delle componenti virali.
La storia della virologia
Dimitri Iwanowski (1892) dimostrò che estratti di piante
di tabacco ammalate potevano trasmettere la malattia ad
altre piante anche dopo aver passato tali estratti attraverso
filtri di ceramica abbastanza selettivi da poter trattenere
anche il più piccolo batterio. Tuttavia non realizzò quanto
era importante questa osservazione
Martinus Beijerinick (1898) confermò ed estese i risultati
di Iwanowski sul Tobacco mosaic virus e fu il primo a
sviluppare il concetto di virus, a cui si riferì come
“contagium vivum fluidum” (un agente vivente solubile)
La storia della virologia
Freidrich Loeffler & Paul Frosch (1898) dimostrarono
che un agente simile era responsabile dell’afta
epizootica nel bestiame
Il primo virus umano identificato, nel 1901, è stato
quello responsabile della febbre gialla
La scoperta dei batteriofagi
Frederick Twort (1915) & Felix d'Herelle (1917) sono stati i
primi ad identificare i virus che infettano i batteri che
d'Herelle chiamò batteriofagi (mangiatori di batteri)
Negli anni 1930s e nei decenni successivi, Salvador Luria,
Max Delbruck e molti altri utilizzarono questi virus come
sistemi modello per studiare molti aspetti della virologia,
comprendenti la struttura dei virus, la genetica, la
replicazione etc.
La storia della virologia è la storia dello sviluppo di sistemi
e strumenti sperimentali che potevano permettere lo studio
dei virus, attraverso il quale si sono aperte intere nuove
aree della biologia.
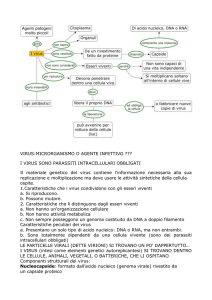
Proprietà generali dei virus
Parassiti intracellulari obbligati Mancano di tutte, o la
maggior parte delle informazioni genetiche che
codificano l’apparato necessario per la produzione di
energia metabolica e per la sintesi proteica
Le particelle virali (virioni) sono formate da acido
nucleico rivestito da un involucro proteico (capside). In
alcuni virus è presente una membrana pericapsidica
contenente lipidi e glicoproteine (envelope)
Il genoma virale consiste di DNA o RNA, ma non di
entrambi.
Il genoma virale guida la sintesi delle componenti virali
all’interno di una cellula ospite appropriata.
Proprietà generali dei virus
Le particelle virali sono prodotte come risultato
dell’assemblaggio di componenti preformati; le
cellule “crescono” grazie all’aumento controllato dei
loro componenti e si moltiplicano mediante il
processo di divisione. I virioni non crescono e non
vanno incontro a divisione
Dimensioni
Dimensioni sub-microscopiche 20-400 nm
From Flint et al. Principles of Virology (2000), ASM Press
Forme
Forme
Classificazione tassonomica
Gerarchia. La famiglia ha suffisso viridae. Il
genere ha suffisso virus. La specie.
Struttura del virione
Sistema classico di classificazione
• Natura dell’acido nucleico e
configurazione del genoma
• Simmetria del rivestimento proteico
(capside)
• Presenza o assenza di involucro lipidico
(envelope)
Genoma dei virus
Virus a DNA: tra questi, quasi tutti i virus animali
contendono DNA a doppio filamento, con l’eccezione dei
Parvoviridae (virus adeno-associati) e dei Circoviridae.
Genoma dei virus
Virus a RNA: quasi tutti i virus a RNA contengono RNA a
singolo filamento, con l’eccezione dei Reoviridae (p.e.
rotavirus) che contengono RNA a doppio filamento. I virus
ad RNA sono ulteriormente suddivisi in:
Genoma dei virus
• Virus con genoma a RNA a filamento positivo: cioè
genomi con la stessa polarità dell’RNA messaggero. I
retrovirus contengono due copie di RNA+
• Virus con genoma ad RNA a filamento negativo:
cioè genomi con polarità opposta a quella dell ’ RNA
messaggero. Tre membri di questa classe sono
sufficientemente correlati da essere inclusi nello stesso
ordine
dei
Mononegavirales
(rhabdoviridae,
paramixoviridae e filoviridae) Gli altri virus di questa
classe hanno un genoma segmentato (orthomyxoviridae
con
8 segmenti, arenaviridae con 2 segmenti e
bunyaviridae con 3 segmenti). Queste ultime due famiglie
sono uniche in quanto possiedono un genoma definito
ambisenso (cioè un genoma contenente RNA a polarità
sia + che -)
Funzioni delle proteine virioniche di
rivestimento
Protezione del genoma
•Assemblaggio di un rivestimento proteico protettivo e stabile
•Riconoscimento specifico e impacchettamento del genoma
•In molti virus, interazione con la membrana dell’ospite per formare
l’envelope
Trasporto del genoma
•Legame specifico con i recettori della cellula ospite
•Segnali specifici che inducono la liberazione del genoma all’interno della
cellula ospite (uncoating)
•Induzione della fusione dell’envelope con la membrana dell’ospite
•Interazione con componenti cellulari che permettono il trasporto del
genoma al sito appropriato per la replicazione
Struttura del virione
Capside (alcune volte denominato nucleocapside):
struttura proteica protettiva che circonda il genoma virale.
E’ costituito da subunità proteiche assemblate a formare
una struttura simmetrica ripetitiva. Le principali classi di
simmetria sono quella elicoidale e quella icosaedrica. Le
dimensioni del capside determinano in qualche modo la
quantità (quindi le dimensioni) del materiale genetico che
può essere impacchettato nella particella virale.
Nomenclatura
Subunità strutturale (protomero): le singole proteine che
costituiscono il capside
Unità morfologica (capsomero, simmetria icosaedrica):
la struttura più piccola visibile al microscopio elettronico,
formata dall’interazione di più protomeri
•Pentoni: capsomeri formati da cinque protomeri
•Esoni: capsomeri formati da sei protomeri
Simmetria elicoidale
Le dimensioni dei virioni con un capside a simmetria elicoidale sono
date in termini di diametro, che dipende dalle caratteristiche dei
protomeri, e di lunghezza, che dipende dalle dimensioni del genoma.
La simmetria elicoidale è definita da due parametri:
Ampiezza = diametro
Passo dell’elica (P) = distanza coperta da un intero giro di elica
P=m x r
m= numero di protomeri per giro d’elica
r= incremento assiale per subunità
From Cann Principles of molecular virology (2001). Academic Press
Simmetria elicoidale
La simmetria elicoidale è molto frequente
tra i virus vegetali
Simmetria elicoidale
I virus animali a simmetria elicoidale sono tutti provvisti di envelope
Numerosi virus patogeni per l’uomo sono caratterizzati da questa struttura: virus
dell’influenza (orthomyxovirus), i virus che causano la parotite epidemica e il
morbillo (paramyxovirus), il virus della rabbia (rabdovirus)
Virus della rabbia
From Cann Principles of molecular virology (2001). Academic Press
Simmetria Icosaedrica
L’icosaedro è un solido con 20 facce triangolari e 12 vertici;
È l’unico involucro chiuso che si può ottenere con protomeri identici
(il capside elicoidale è una struttura aperta);
È caratterizzato da 3 assi di simmetria rotatoria:
asse di simmetria 5 che passa attraverso ognuno dei dodici vertici,
sono possibili 5 rotazioni di 72°, ciascuna delle quali produce una
configurazione identica
asse di simmetria 3, che passa per il centro di ciascuna delle venti
facce, sono possibili tre rotazioni di 120°, ciascuna delle quali
produce una configurazione identica
Asse di simmetria 2, che passa per ciascuno dei trenta spigoli
dell’icosaedro, sono possibili 2 rotazioni di 180°, ciascuna delle quali
produce una configurazione identica
Simmetria Icosaedrica
Gli involucri più semplici sono formati da 60 protomeri, tre per faccia,
ognuno posto ad uno dei vertici.
L ’ insieme dei cinque protomeri attorno a ciascun vertice
dell’icosaedro costituisce un capsomero (in questo caso un pentone)
Solo i virioni più piccoli e più semplici hanno un capside composto da
60 protomeri, ed alcuni esempi si trovano tra i virus delle piante
Simmetria Icosaedrica
La maggior parte degli altri virus con capside a simmetria icosaedrica
hanno più di 60 protomeri
Numero di triangolazione (T). Numero di triangoli inscritti per faccia.
Il più piccolo numero possibile è 3, poi 4, 7, 9, 12 … quindi avremo
T=1 per l’icosaedro base
T=3 Tre triangoli inscritti per ciascuna faccia (sei emi-triangoli)
T=4 Quattro triangoli inscritti per ciascuna faccia etc…
From Flint et al. Principles of Virology (2000), ASM Press
Simmetria Icosaedrica
Principio di semi-equivalenza: quando la stessa unità strutturale o
protomero si organizza a formare sia pentoni che esoni, le diverse
interazioni stabilite nel formare i due tipi di capsomeri sono da
attribuire e differenze di configurazione della stessa catena
polipeptidica.
Poiché ciascun triangolo inscritto è formato da 3 protomeri, uno per
vertice, nei capsidi icosaedrici con T>1 il numero di protomeri sarà
60xT
Adenovirus T=25
HSV-1 T=16
Struttura del virione
Struttura del virione
Involucro pericapsidico (envelope): doppio strato
lipidico che circonda il capside di molti virus animali.
Deriva dalle membrane cellulari come risultato del
processo di gemmazione della particella virale. L’envelope
contiene anche proteine, codificate dal genoma virale;
spesso glicoproteine con un ruolo importante nel processo
di attacco/adsorbimento e entrata del virus nella cellula
ospite. La presenza dell ’ envelope conferisce minore
stabilità al virione.
VIRAL ENVELOPE
Al doppio strato lipidico derivato dalle
membrane cellulari sono associate proteine
virus-specifiche quali:
•Glicoproteine, suddivise in base alla loro
funzione in: glicoproteine esterne o di
superficie, proteine transmembrana e canali
di trasporto
•Proteine della matrice
From Flint et al. Principles of Virology (2000), ASM Press
From Flint et al. Principles of Virology (2000), ASM Press
Simmetria Icosaedrica
La maggior parte degli altri virus con capside a simmetria icosaedrica
hanno più di 60 protomeri
Pentone
Esone
From Flint et al. Principles of Virology (2000), ASM Press