TESSUTO MUSCOLARE
Ha la possibilità di contrarsi si usa per il movimento.
Tipi di tesuto muscolare che abbiamo già visto:
- Le cellule mioepiteliali si trovano tra membrana basale e ghiandole secretrici. Aiutano a secretare.
Sono cellule epiteliali con la caratteristica della contrazione.
- Periciti: si possono contrarre e sono appena fuori dalla membrana basale. Possono diventare cellule
muscolari lisce a tutti gli effetti.
- Fibroblasti: li troviamo quando c’è riparazione di una ferita; contraendosi fanno si che si colonizzi la
parte mancante.
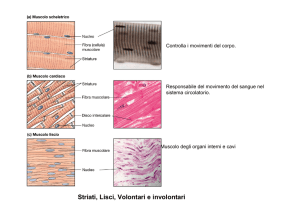
Ci sono due tipi di tessuto dal punto di vista morfologico:
Striato: ha delle bandeggiature.
- Viscerale (non c’è osso)
- Scheletrico (dove c’è osso)
Liscio: preposto al movimento involontario preposto alla vita. Sono controllati da sistema nervoso
autonomo o ormoni (ex. Apparato gastrointestinale, respiratorio)
Cardiaco: si contrae spontaneamente, ma si può avere controllo della frequenza del battito tramite
il sistema simpatico
SCHELETRICO
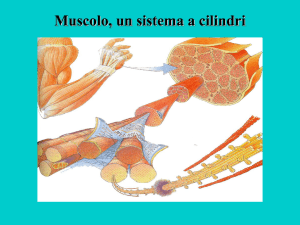
Sono muscoli abbastanza grossi. Attorno c’è del connettivo che permette di portare al muscolo dei vasi. C’è
un rivestimento connettivale (epimisio esterno). Dall’epimisio si dipartono dei setti (perimisio). Da questo si
distacca connettivo che separa le singole fibre muscolari.
La fibra muscolare è una cellula plurinucleata, lunghissima rivestita da endomisio (connettivo). Ha una
struttura di membrana esterna (tutto ciò che si distacca dal connettivo è rivestito da una membrana
esterna, parlo di membrana basale quando a toccarsi sono epitelio e connettivo).
Fibre sono strutture cilindriche compatte con un diametro costante (100 micron). È polinucleata (bisogna
mantenere un certo rapporto tra nucleo e citoplasma) e i nuclei si trovano appena al di sotto della
membrana, perché il citoplasma è tutto occupato da proteine contrattili. La fibra è rivestita da membrana
esterna a sua volta nell’endomisio.
Mioblasti derivano dal mesoderma (dà origine anche a tessuti mesenchimali). I mioblasti hanno un unico
nucleo. Le cellule si fondono a formare miotubi, che hanno ancora i nuclei centrali; sono centrali
normalmente, si spostano per via delle proteine. Poi queste cellule producono i filamenti che spostano i
nuclei in periferia.
Nuclei centrali= immaturità cellulare
Ci sono tante miofibrille a loro volta bandeggiate. Sono tutte in registro (ovvero in fase).
Bande scura: A. filamenti più spessi
Banda chiare: I. filamenti più sottili
Nella banda chiara c’è linea z.
Da stria z a stria z ho il sarcomero. I sarcomeri si susseguono e hanno lunghezza media attorno ai 3 micron.
Anche nella banda scura c’è una piccola banda scura (M) su un piccolo ambiente chiaro (zona H).
Tra le varie miofibrille ci sono i mitocondri.
Sincizio: cellula polinucleata derivata dalla fusione di più cellule nucleate.
L’unità morfologica è il sarcomero.
Sarcolemma (plasma lemma+ membrana basale). La membrana basale funge da colla affinchè le singole
fibre muscolari agiscano insieme. Le singole fibre sono innervate. È PAS positivo
Sarcoplasma
Reticolo sarcoplasmatico: contiene moltissimi ioni calcio
Mitocondri sono molto crestati; così aumenta la superficie e si ha maggiore produzione di energia.
Mioglobina: proteina globulare che lega ossigeno.
Citoplasma molto scarso.
Actina è una molecola che c’è in tutte le cellule, perché serve a muoversi.
Ciò che fa avvenire la contrazione è associazione di actina e miosina.
Stria M e Z servono per attracco.
Actina circondata da 3 filamenti di miosina. Miosina circondata da 6 filamenti di actina. Sono disposti in
modo molto ordinato.
La lunghezza del sarcomero può cambiare. Durante la contrazione si annulla la banda I e il sarcomero si
accorcia. Rimane costante la banda A (che contiene miosina), mentre le zone che contengono solo actina
(quindi le zone chiare)variano.
MIOSINA
Fatta da una zona fibrosa (di supporto) e teste globulari che hanno attività atp-asica, oltre che hanno sito di
legame per actina.
Si assemblano dalla parte c terminale (linea M) e vanno nelle direzioni opposte. Le teste sporgono per
prendere contatto con actine.
Alle teste sono associate catene leggere che regolano l’interazione con actine.
La zona H è solo fibrosa (non fuoriescono teste)
ACTINA
È una proteina globulare che si associa in filamenti stabilizzati da tropomiosina.
Troponina I,C(lega il calcio che elimina inibizione),T: bloccano il sito di legame di actina per miosina.
La testa di miosina tocca actina, si piega, tira e sposta actina, si stacca e va a interagire con un’altra, ecc...
la parte globulare di miosina cammina su quella di actina.
Con rigor mortis il muscolo diventa duro. Manca atp e si ha interazione statica di actina con miosina. In vita
atp stacca miosina da actina. Atp diventa adp e l’energia ricavata serve a risposare la testa, poi si rilascia
adp e si attende altro atp per far ricominciare il giro ( se si è morti, l’atp non arriva e il muscolo resta
contratto.)
MECCANISMO DI CONTRAZIONE
Ogni volta c’è lo spostamento di qualche sub unità di actina.
Quando lego ATP actina e miosina si staccano. L’idrolisi dell’ATP fa cambiare la conformazione della testa
della miosina.
Poiché serve molta energia (glicogeno è energia in tempi veloci) si ha molta ATP. Creatina lega ATP e così fa
in modo che possa essere accumulata. L’atp lo prende dal mitocondro.
Filamenti spessi: miosina
Filamenti sottili: actina filamentosa.
Ci sono proteine che permettono il mantenimento di questa struttura e delle reciproche distanze tra le
varie parti. Queste proteine sono il 25% di quelle che fanno parte della miofibrilla
TITINA: àncora filamenti spessi alle linee Z. Evita l’eccessivo stiramento del sarcomero.
NEBULINA: ancra i filamenti sottili alle linee Z e ne regola la lunghezza durante lo sviluppo
ADESINA, FILAMINA, ACTININA: mantiene coese alle zone di attacco
TROPOMODULINA: lega l’actina e si trova all’estremità libera dei filamenti di actina, per regolare la
lunghezza del sarcomero
MIOMESINA, PROTEINA C: servono di attacco per attacco della miosina
PLECTINA
ALFA/B CRISTALLINA
DESMINA: da qui si diparte actina
DISTROFINA: molto grande il suo gene. Nel caso della distrofia muscolare il gene è mutato.
Lega i Distroglicani (che possono essere mutati).
I distroglicani hanno una parta proteica.
Ai distroglicani è legata la laminina. I sarcoglicani sono associati ai distroglicani.
Se non abbiamo buona stabilità dei filamenti il sistema si sfascia e non si ha buona attività di contrazione
SARCOPLASMA E RETICOLO SARCOPLASMATICO
Il reticolo sarcoplasmatico è fatto da tubuli e cisterne collocate (nel muscolo scheletrico) nella zona di
passaggio tra banda A e I. La parte terminale dei tubuli è collegata e forma le cisterne terminali. Sono legati
al tubulo a T. Il tubulo a T è un’invaginazione della membrana della cellula: tanti formano il sistema dei
tubuli trasversali/sistema a T.
Tubulo T e le due cisterne terminali formano la triade, che si ha a livello di ogni tratto sarcomerico.
Quando si deve contrarre il muscolo, le molecole che servono devono essere reclutate in fretta. Il tubulo T
è la membrana della cellula invaginata a stretto contatto con reticolo sarcoplasmatico e viene
depolarizzata. Questa depolarizzazione viene trasmetta al reticolo sarcoplasmatico che è una riserva di ioni
calcio. Quando è depolarizzato libera calcio che si lega a troponina C che cambia troponina I che permette il
contatto tra actina e miosina. Si può dire che il calcio attiva una reazione a catena di troponina e
tropomiosina.
Questi fenomeni avvengono in millisecondi.
Sulla superficie del reticolo sarcoplasmatico c’è una calcio-atp-asi che pompa di nuovo il calcio all’interno
del reticolo.
Il calcio è mantenuto in soluzione perché legato a calsequestina: evita che il calcio precipiti, ha bassa
affinità (quindi il calcio si stacca facilmente per attaccare tropo miosina) facilmente) ma molti siti di legame.
CIRCOLAZIONE E INNERVAZIONE
Ogni singola fibra muscolare del tessuto scheletrico è innervata (con il cardiaco non è così).
In base alla quantità di innervazione si ha movimento più o meno preciso.
A livello di ogni muscolo si ha vascolarizzazione diversa.
Ci sono fibre:
Rosse
- Contrazione lenta
Posturali: non consumano grande quantità di energia
Sono isometriche (mantengono la loro lunghezza
Anaerobiche (bianche): usano la beta ossidazione dei lipidi e il glicogeno
Per contrazione veloce
Diminuiscono di lunghezza
Bianche
2b
Quelle contrapposte alle rosse sono le bianche 2b, sottoposte ad affaticamento
Le 2a hanno doppio metabolismo (aerobico e anaerobico)
Si possono distinguere istologicamente questi tre tipi di fibre. In ogni muscolo abbiamo tutti e tre i tipi in
proporzioni diverse a seconda della funzione del muscolo.
RIGENERAZIONE
Il muscolo se va incontro ad atrofia può rigenerarsi.
Atrofia: si ha diminuzione del volume delle fibre, perdita di massa.
L’attività è molto importante per evitarla.
Se uno taglia il nervo va incontro a atrofia.
Ipertrofia: il muscolo aumenta di dimensione (non aumenta il numero di fibre). Può prendere anche il
muscolo cardiaco.
Iperplasia: aumento del numero, ma nel muscolo si ha ipertrofia.
Nel muscolo ci sono cellule satellite che sono una sorta di cellula staminale del muscolo. Di solito sono tra
la membrana e le fibre. Non è facile vederle.
Queste cellule satelliti possono esprimere molecole per la regolazione miogenica:
MyoD
Mrf
La riserva di cellule staminali basta all’individuo per tutta la sua vita. Dopo un po’ le fibre muscolari si
usurano e quindi è normale che queste cellule vengano usate.
Distrofia di Duchenne: la distrofina non è resistente alle contrazioni della fibra muscolare. Si devono
continuamente inserire cellule satelliti nel muscolo, ma queste non possono bastare per tutta questa
domanda. I loro nuclei sono centrali e le fibre hanno dimensioni diverse. Finchè c’è riserva di cellule
staminali, l’individuo vive tranquillamente. Dopo un po’ le cellule staminali finiscono e si hanno problemi.
Ci sono altri tipi di distrofia che colpiscono le proteine che servono a tenere la struttura del sarcomero e
delle fibre.
LISCIO
Non è volontario. Il controllo nervoso qui è simpatico.
Non ha striature.
Le cellule sono fusiformi. Al centro hanno il nucleo e quindi sono più allargate. Il nucleo è unico.
Si ha sempre una componente connettivale molto importante, soprattutto collagene. Le cellule, anche se
sono singole, funzionano da scinzizio, funzionale appunto.
Con la contrazione il nucleo non è più ovoidale.
Si trova poco negli apparati respiratori, molto in quello gastroenterico.
Ci sono due strati con le fibre disposte perpendicolarmente.
Sono lunghe 100 micron e di diametro 20 micron. A volte la cellula diventa molto lunga , nel miometrio
(parete muscolare dell’utero. Questo serve per il parto).
Ci sono gap-junction che permettono alle cellule di comportarsi da scinzizio, non tutte le cellule sono
innervate (non sono innervate da motoneuroni, che troviamo solo nel muscolo scheletrico); quando arriva
la depolarizzazione questa è trasmessa tramite gap-junction e così si aprono i canali del calcio.
Caveole: sono vescicole che fungono da riserva di calcio. Derivano da cellule pinocitotiche.
Placche dense: si trovano sulla superficie della cellula e sul citoplasma. Qui si attaccano i filamenti di actina
(cfr linea Z nel muscolo scheletrico).
I filamenti non sono organizzati in modo ordinato nello spazio.
Contrazione: cellula diventa più corta e si inspessisce.
Muscolo scheletrico bipolare
Muscolo liscio polarità laterale
Caldesmon e calponina sono molecole regolatorie (mascherano siti di interazione tra actina e miosina) e
marcatori del muscolo liscio (cfr troponina nel muscolo striato).
Calcio (viene da vesciocole esterne e reticolo endoplasmatico liscio)si lega a calmodulina e attiva chinasi
della catena leggera della miosina (molecola regolatoria) che fosforila la catena leggera della miosina
cambia conformazione della miosina:
testa interagisce con actina
si srotola la parte arrotolata
Nel muscolo scheletrico il calcio lavorava sull’actina rimuovendo l’inibizione.
Il calcio si occupa anche di rimuovere calponina.
Canali del calcio: si trovano sulla superficie.
La cellula è attivata per via ormonale, nervosa (con neuro modulatori), tramite stimolazione meccanica che
parte quando si sente l’ambiente (a livello dello stomaco, quando si sente l’organo pieno, si ha movimento
per far defluire il contenuto).
CARDIACO
È striato, ma le cellule sono mononucleato ( o binucleate) e il nucleo si trova centralmente.
Si vengono a creare fibre muscolari, diverse da quelle scheletriche. Questa si può ramificare.
Si ha una zona in cui le cellule si giuntano una con l’altra, nei dischi intercalari, situati dove si trova la stria z
(non in tutte le cellule).
Si hanno tre sistemi di giunzioni all’interno dei dischi intercalari:
Desmosomi
Fasce aderenti
Gap-junction: servono per far passare la comunicazione della contrazione è un scinzizio
funzionale
Si ha una struttura come quella dello scheletrico.
I mitocondri sono crestati: la membrana interna è più espansa e si ha maggiore efficienza per gli enzimi di
membrana e quindi maggiore produzione di energia.
Lipofuscina: molecola che si accumula con la vecchiaia della cellula.
Sono cellule perenni che possono ricambiarsi minimamente.
Nei punti importanti alla giunzione dove si ha maggiore trazione, ovvero parallelamente alla linea Z
(componente trasversale ei dischi intercalari), si hanno desmosomi e fasce di adesione a cui si attaccano
filamenti di actina. Perpendicolarmente (componente laterale dei dischi intercalari) si trovano le gapjunction.
Si parla di diade e non triade.
La depolarizzazione dura per tempi più lunghi. Si hanno molti canali del calcio sulla superficie del
cardiomiocita. Anche qui abbiamo pompe del calcio per riportare il calcio all’interno.
L’impulso alla contrazione parte nel nodo seno atriale (atrio destro), un vero e preoprio pacemaker e da qui
si diparte nel resto del cuore.