LA COMUNICAZIONE CELLULARE
LE CELLULE SONO CAPACI DI COMUNICARE E DI COORDINARSI
La comunicazione fra cellule avviene in maniera analoga a quella
che si realizza fra gli individui, mediante segnali. Il segnale parte
da una cellula e arriva alla cellula destinataria che lo recepisce e
risponde in maniera adeguata.
Se il segnale proviene da una cellula segnalante lontana dalla cellula
target e quindi viaggia attraverso il circolo sanguigno, la
segnalazione si dice di tipo endocrino.
Se il segnale proviene da una cellula segnalante vicina alla cellula
target la segnalazione si dice di tipo paracrino.
Segnalazione paracrina
Se la cellula segnalante è anche cellula target, la segnalazione si
dice di tipo autocrino.
Esiste anche una segnalazione dipendente da contatto mediata da
segnali legati alla membrana della cellula segnalante. In questo caso
cellula segnalante e cellula target devono venire in contatto perché la
segnalazione avvenga.
SEGNALAZIONE NEGLI ORGANISMI
UNICELLULARI
La segnalazione nei lieviti può
essere utilizzata in condizioni
sfavorevoli per la riproduzione.
Mating-type a
Mating-type a
Le molecole segnalanti, dette anche ligandi, possono
essere:
NEUROTRASMETTITORI
ORMONI
FEROMONI
GLI ORMONI
Gli ormoni possono essere lipofilici e allora possono attraversare la
membrana e raggiungere il loro recettore all’interno della cellula
target oppure possono essere idrofilici e allora il loro recettore si
trova sulla membrana plasmatica della cellula target.
Come agisce un segnale che attraversa la membrana
L’esempio più semplice di come può agire
un segnale che si lega ad un recettore
situato nella membrana plasmatica è
rappresentato dall’apertura e dalla
chiusura di un canale proteico (canali a
controllo di ligando).
L’interazione ligando-recettore
apre il canale ionico
Come agisce un segnale che genera una risposta
all’interno della cellula: la trasduzione del segnale
Un esempio di trasduzione del segnale: recettori tirosinchinasici
Un esempio di trasduzione del segnale: recettori accoppiati a proteine G
In molti casi l’interazione ligando-recettore attiva una serie di proteine
nella faccia citoplasmatica della membrana che ha come risultato finale
la produzione di un secondo messaggero del segnale che va a scatenare
nella cellula altre reazioni.
Il meccanismo che produce il secondo messaggero è complesso e
coinvolge una serie di reazioni.
Struttura del recettore che si lega alla proteina G
Schema grafico della trasduzione del
segnale mediata dalla proteina G
trimerica
0. Prima che il ligando interagisca con il
recettore, recettore ed effettore sono
inattivi e la proteina G è trimerica e legata
a GDP.
1. Il ligando attiva il recettore
inducendone un cambio conformazionale
e consentendo il legame con la subunità a
della proteina G.
2. Il legame recettore-proteina G provoca
nella proteina G la dissociazione della
subunità a dalle subunità b e g
permettendo lo scambio fra GDP e GTP. 3.
La subunità a, legata a GTP, si lega
all'effettore e lo attiva, mentre il ligando si
dissocia dal recettore.
L'idrolisi del GTP provoca la dissociazione
della subunità a dall'effettore e la sua
riassociazione alle altre due subunità.
La trasduzione mediata da proteina G può essere irreversibilmente modificata da
tossine batteriche. La tossina prodotta dal batterio del colera (Vibrio cholerae) altera
la sottounità Ga rendendo impossibile la inattivazione dell’adenilato ciclasi. Questo
produce un massivo passaggio di acqua dal sangue nel lume intestinale con
l’induzione di una violenta diarrea che può portare anche alla morte.
EFFETTORI DELLE PROTEINE G: ADENILATO CICLASI E FOSFOLIPASI C
SECONDO MESSAGGERO
SECONDO MESSAGGERO
L'adenilatociclasi produce cAMP che funziona come secondo
messaggero. cAMP si lega alla proteinchinasi A, cAMPdipendente (cAPK), composta da quattro subunità, due
regolatrici (R) e due catalitiche (C). Il legame di cAMP alle
subunità regolatrici induce la dissociazione e la concomitante
attivazione delle subunità catalitiche che vanno a fosforilare i
corrispondenti
substrati.
La
fosfolipasi
agisce
sul
fosfatidilinositolo bifosfato producendo IP3, che apre i canali
Ca2+ IP3-dipendenti del reticolo endoplasmatico, e DAG che
attiva la PKC che, a sua volta, fosforila i corrispondenti substrati.
http://bcs.whfreeman.com/lodish5e/pages/bcsmain.asp?v=category&s=00010&n=13000&i=13010.01&o=|00510|005
20|00530|00540|00560|00570|00590|00600|00700|00010|00020|0
0030|00040|00050|01000|02000|03000|04000|05000|06000|0700
0|08000|09000|10000|11000|120
Fosfolipasi C
AZIONE DELL’AMP CICLICO
AMP ciclico
Chinasi cAMP-dipendente
cAPKs
Fosforilazione di enzimi (nei residui di serina e treonina)
Aumento attività catalitica
Diminuzione attività catalitica
Il secondo messaggero permette di
amplificare notevolmente il segnale
L’arrivo di un segnale esterno ad una cellula ha spesso come risultato finale
l’attivazione della trascrizione di un gene specifico
http://bcs.whfreeman.com/lodish5e/pages/bcsmain.asp?v=category&s=00010&n=13000&i=13010.02&o=|00510|00520|00530|00540|00560|00570|00590|0060
0|00700|00010|00020|00030|00040|00050|01000|02000|03000|04000|05000|06000|07000|08000|09000|10
000|11000|120
Proteine G e adenilato ciclasi
La stessa molecola segnale può
indurre risposte differenti che
dipendono dalle cellule bersaglio e
dai recettori.
A Real Example of Cell Communication - YouTube
Negli organismi unicellulari ogni cellula agisce e si divide
indipendentemente dalle altre. Il più importante fattore
limitante la sua indipendenza è rappresentato dalla
quantità di nutrienti presenti nel mezzo.
Esistono infatti precisi momenti, collegati a carenza di nutrienti, in cui queste
cellule comunicano fra loro e si coordinano. Questo avviene quando, ad
esempio, nel lievito (Saccharomyces cerevisiae), si differenziano le due
tipologie cellulari complementari (mating type) che si accoppieranno.
Quando mancano i nutrienti, l’ameba della specie Dictyostelium
discoideum mette in atto un meccanismo di aggregazione cellulare
con formazione di un organismo pluricellulare semplice, ma capace
di mantenere e proteggere spore.
Finché è presente nutrimento, queste amebe vivono
come organismi unicellulari. Se questo viene a
mancare, le cellule si dirigono verso un centro comune
(a) dove si aggregano e formano una specie di
mucillagine (b) da cui si allunga un peduncolo (c).
(a)
(c)
(b)
Dal momento in cui le cellule si
aggregano e formano il pluricellulare
nelle diverse regioni si
differenziano tessuti
specifici. Nel peduncolo
si forma un tessuto
di sostegno e all’apice si forma
un corpo fruttifero con le spore.
Come è stato dimostrato
sperimentalmente, il segnale
che induce l’aggregazione e
che viene emesso quando
le cellule vengono
affamate è l’AMP ciclico.
Slime Molds Time Lapse - YouTube
“…no more of a bag of amoebae
encased in a thin slime sheath..”
• La pluricellularità implica che le cellule stiano in comunicazione le une con
le altre in un sistema di rete complesso che coordina tutte le attività, dal
differenziamento al metabolismo. Le cellule comunicano con segnali
(molecole segnalanti) che partono da una cellula segnalante ed arrivano
alla cellula target che ha i recettori per ricevere tale segnalazione. Per
segnalare possono essere usate una gran varietà di sostanze come peptidi,
proteine, piccole molecole lipofiliche o derivati da amminoacidi.
• Quando la segnalazione arriva alla superficie della cellula esiste un
complesso meccanismo che converte la segnalazione esterna in una
segnalazione interna. Tale meccanismo si chiama trasduzione del segnale.
FORMAZIONE DEI TESSUTI
Come le cellule si integrano per formare tessuti: molecole di adesione,
giunzioni cellulari e matrice extracellulare
Come si presenta l’interfaccia matrice-cellula al microscopio elettronico
PRINCIPALI FAMIGLIE DI MOLECOLE DI ADESIONE
Le N-CAM e le proteine dimeriche E-caderine di una cellula formano legami omofilici con molecole
presenti sulla membrana della cellula adiacente. Le selectine, anch'esse proteine dimeriche,
formano legami eterofilici con una catena oligosaccaridica di una glicoproteina presente sulla
membrana della cellula adiacente. Anche le integrine interagiscono con molecole differenti, per
esempio le ICAM, mediante adesione eterofilica.
LE CADERINE
Tipo di caderina
E-caderine
P-caderine
N-caderine
Distribuzione cellulare
Embrione, tessuti epiteliali
non neuronali
Trofoblasto
Tessuto nervoso, tessuto muscolare
scheletrico e cardiaco
Attivazione delle caderine per mezzo
del calcio e riconoscimento omofilico
Le caderine servono per far aderire tra loro le cellule appartenenti allo stesso
tipo di tessuto. La porzione intracellulare delle caderine aggancia l’actina
citoscheletrica attraverso proteine (catenine) che fanno da intermediarie.
catenine
Una volta che le cellule si sono riconosciute come appartenenti allo
stesso tessuto attraverso le caderine, si instaura fra loro un più
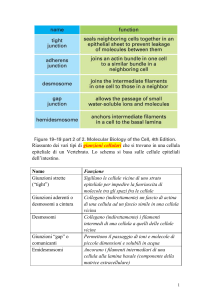
stretto rapporto mediante la formazione di GIUNZIONI. Le caderine
formano le GIUNZIONI ADERENTI e i DESMOSOMI. Nelle cellule si
possono poi formare GIUNZIONI GAP e GIUNZIONI TIGHT.
Intermediate
filaments
GIUNZIONI CELLULARI
Le giunzioni cellulari sono strutture che permettono
un’associazione a lungo termine tra cellule adiacenti.
1.
2.
3.
4.
GIUNZIONI TIGHT (Tight junctions)
GIUNZIONI ADERENTI (Adherens junctions)
DESMOSOMI (Desmosomes)
GIUNZIONI GAP (Gap junctions)
GIUNZIONI TIGHT
GIUNZIONI ADERENTI
DESMOSOMI E EMIDESMOSOMI
DESMOSOMA
EMIDESMOSOMA
DESMOSOMI
INTEGRINE
Questo tipo di giunzione mette in comunicazione diretta le cellule. Sono
infatti costituite da gruppi di canali, i connessoni presenti nelle due cellule
adiacenti e in asse fra loro. Ogni connessone è fatto da 6 copie della proteina
connessina.
GIUNZIONE GAP
GIUNZIONE GAP
SELECTINE: mediano l’interazione con
oligosaccaridi nel processo infiammatorio
C-Type Lectin Domain
P-selectin
EGF-Like Domain
Complement Regulatory Repeat
NH2
Transmembrane Domain
Cytoplasmic Domain
E-selectin
NH2
L-selectin
NH2
Leukocyte
Endothelial Cell
COOH
COOH
COOH
L-selectina: neutrofili, monociti, linfociti B e T
P-selectina: granuli a delle piastrine non attivate e
corpi di Weibel-Palade delle cellule endoteliali
(espressa costitutivamente nelle piastrine ed
indotta nelle cellule endoteliali)
E-selectina: cellule endoteliali e siero (espressione
indotta da citochine)
Rolling leucocitario nella
risposta infiammatoria
http://bcs.whfreeman.com/lodish5e/pages/bcsmain.asp?v=category&s=00010&n=06000&i=06010.01&o=|00510|00520|00530|
00540|00560|00570|00590|00600|00700|00010|00020|00030|00040|00050|0
1000|02000|03000|04000|05000|06000|07000|08000|09000|10000|11000|12
03&n