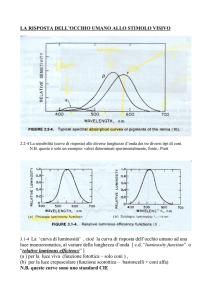
Lezione del 17/05/13
Prof.Anastasi
Fino adesso avevamo fatto uno schema cellulare della retina.
Possiamo dire che la retina è costituita verticalmente da 4 tipi di cellule, che dall’esterno all’interno sono:
-epitelio pigmentato;
-fotocettori;
- cellule bipolari;
- cellule gangliari.
Le gangliari sono le più interne di tutte; l’epitelio pigmentato è il più esterno. Badate che non sono le
uniche cellule, vedremo che ce ne saranno altre, ma per adesso consideriamo la catena verticale: questo
vuol dire che la luce deve attraversare questi strati! La luce arriva alla cornea, attraversa il cristallino e va a
colpire la retina. Questo che vuol dire? Vuol dire che colpisce lo strato delle fibre delle cellule gangliari, che
emettono un assone (ora vedremo dove va questo assone): colpisce quindi questo assone NON
MIELINIZZATO, trasparente; se fosse mielinizzato non arriverebbe luce.
Quindi: ASSONE NON MIELINIZZATO- CELLULE GANGLIARI- CELLULE BIPOLARI- FOTOCETTORI- EPITELIO
PIGMENTATO.
Nella retina abbiamo un asse ottico, cioè l’asse che congiunge il polo anteriore con il polo posteriore
dell’occhio. Questo però non è l’asse della visione discriminata, perché la visione discriminata è in una zona
depressa della retina, un infossamento della retina, che prende il nome di fovea.
La fovea si trova precisamente al centro di una zona iperpigmentata, la cosiddetta macula lutea.
Se io esamino il fundus dell’occhio di un paziente, cercherò come punto di riferimento la zona
iperpigmentata, 1/8 di grado lateralmente all’asse ottico, perché so che al centro c’è la fovea.
La fovea è la zona di massima discriminazione dell’immagine.
Se invece mi sposto lateralmente a questa… [Studente: “E’ il fuoco?” Prof: “ No, il fuoco è il cristallino.
Facciamo un esempio: io guardo la signorina, o guardo lui, o la signorina . Ma “guardo” cosa vuol dire?
Guardo tutti… Non basta avere la testa in questa direzione, vuol dire che l’immagine della signorina, o la
tua, mi cade….(non finisce la frase) …Guardo tutti perché se guardo lui passo per…(si porta la mano
all’orecchio), se guardo lei passo per maniaco sessuale…quindi è meglio non guardare nessuno, guardo il
mio dito!
Ma questo che vuol dire? Vuol dire far cadere la sua immagine, o la sua, nella fovea.
Guardare vuol dire “foveare”.
Il mio campo visivo è tutto a fuoco: il fuoco è legato alla lente, e la lente è il cristallino. Per mettere a fuoco
cosa fai?Modifichi la distanza focale o modifichi l’asse di curvatura. In questo caso modifichi l’asse di
curvatura].
Quindi “guardare” vuol dire mettere l’immagine nella fovea.
Se io mi sposto lateralmente all’asse ottico, circa di 1° e ½, ho un punto che si chiama disco ottico.
Il disco ottico è il punto in cui gli assoni di tutte le cellule gangliari della retina si portano dall’interno della
retina a costituire il nervo ottico, cioè il punto in cui nasce il nervo ottico. Immaginate tanti fili tangenti che
ad un certo punto trovano un buco e penetrano. Quindi dal disco ottico nasce il nervo ottico. Ma se io sto
parlando di assoni , parlo più di fotocettori? No! Quindi è un punto cieco!
(Studentessa: “Sono mielinizzati? “ Prof: “No. Fino a quando non fanno parte del nervo ottico non sono
mielinizzati. Quando sono tangenti sulla superficie interna della retina sì, sono senza mielina, quando
entrano nel nervo ottico prendono la guaina mielinica.
Questo è un test classico, lo riporta qualsiasi testo: avete da un lato una crocetta e un “-“ dall’altro, se
mettete due cerchietti , se li avvicinate ai vostri occhi, c’è un punto critico in cui voi ne vedrete uno solo, o
la crocetta o la lineetta, perché la crocetta ricade nella fovea, la lineetta ricade nel disco del nervo ottico,
quindi nel punto cieco. Quindi se lo spostate più in là, li vedete tutti e due, se lo portate ad una certa
distanza focale ne vedete uno, se li avvicinate ancora non vedete più niente.
Quindi io ho un punto cieco, un DISCO cieco, che è la papilla del nervo ottico.
Ragazzi, l’anatomia dell’endoretina ha due dati fondamentali: la fovea e la papilla del nervo ottico.
Dalla papilla del nervo ottico emerge l’arteria centrale della retina, che viaggia al centro del nervo ottico,
torna indietro un istante, ma la trovo all’interno del nervo ottico, con la vena centrale della retina.
Quando emerge, si divide fondamentalmente in due rami, uno che va verso l’alto e uno verso il basso, uno
superiore e uno inferiore; ciascuno di questi due rami si divide a forchetta, con due rami in alto e due rami
in basso; ciascuno dei due rami si divide a sua volta.
Dall’altra parte, la fovea come la riconoscete? La riconoscete perché vedete una zona iperpigmentata!
[Studente: “Cosa sono quella palline nello strato delle cellule…” Prof: “ Quelli sono i corpi cellulari delle
cellule gangliari, ne hanno fatto esplodere alcuni per farvi vedere i principali.”]
Anticipiamo una cosa: quando andate a fare gli strati della retina, il vostro libro dice che essi sono 10 strati,
ma noi finora abbiamo detto che i tipi cellulari sono:
Cellule pigmentate
Fotocettori
Cellule bipolari
Cellule gangliari.
Il primo strato qual è? Quello più esterno, strato dell’epitelio pigmentato.
Poi ci sono i coni e i bastoncelli, ma quando parlo di questi io mi riferisco al loro articolo esterno.
Poi parlo di una cosiddetta membrana limitante esterna, un addensamento mucopolisaccaridico, un
sottilissimo strato trasparente perché le fibrille sono organizzate ortogonalmente per evitare distorsioni.
Dopo c’è lo strato dei granuli esterni, sono i nuclei dell’articolo interno. Quando vai a fare un preparato di
microscopia ottica il citoplasma non lo vedi, vedi i coni e i bastoncelli perché sono alti, l’epitelio pigmentato
perché c’è un bel nucleo e una cellula scura, poi vedi i nuclei dei fotocettori che ci sono nell’articolo interno.
Poi c’è uno strato plessiforme esterno, che è la sinapsi tra fotocettore e cellula bipolare.
Poi vedi lo strato dei granuli interni, il nucleo delle cellule bipolari.
Poi lo strato plessiforme interno, la sinapsi tra le cellule bipolari e le cellule gangliari.
Strato dei corpi delle cellule gangliari.
Strato delle fibre del nervo ottico.
Membrana limitante interna, anch’essa un addensamento di connettivo di mucopolisaccaridi
estremamente organizzati in maniera tale che non ci sia distorsione e che si trova internamente allo strato
delle fibre del nervo ottico.
Quindi, riepilogando:
epitelio pigmentato;
articolo esterno dei coni e dei bastoncelli;
membrana limitante esterna;
strato dei granuli esterni;
strato plessiforme esterno;
strato dei granuli interno;
strato plessiforme interno;
cellule gangliari;
strato delle fibre del nervo ottico;
membrana limitante interna;
L’epitelio pigmentato è un foglietto di derivazione ectodermica, tutto il resto è neuroectodermico.
Kandel dice una cosa molto bella: fondamentalmente qual è la caratteristica della visione? Negli altri sensi il
recettore è in periferia; qua non è in periferia, perché questa è una struttura corticale, è un frammento di
corteccia che si è stratificato in maniera peculiare e si è posto all’esterno. E’ una struttura stratificata a mò
di corteccia, perché deriva dal neuroectoderma.
[Studente: “Professore, non ho capito. Le cellule dell’epitelio pigmentato guardano verso l’interno o verso
l’esterno dell’occhio?” Prof: “All’esterno!A ridosso dello strato vascolare”]
La luce quindi deve attraversare tutti gli strati; per questo ho detto che i mucopolisaccaridi sono disposti
ortogonalmente per evitare la distorsione e le fibre dei nervo ottico sono demielinizzate per essere
trasparenti.
[Studentessa: Professore, ma ho letto che i coni prendono sinapsi sia con cellule bipolari centri-on e centrioff, invece i bastoncelli no,ma non è vero!” Prof: “No, non è vero. Con tutte e due. Lei ha introdotto il
concetto di campo recettoriale. “
In questa catena verticale abbiamo in buona sostanza che più coni e più bastoncelli convergono attraverso
le cellule bipolari su una sola cellula gangliare.
Questo è chiamato campo recettoriale della cellula gangliare.
Possiamo avere sia cellule centro-on che cellule centro-off, sia sui coni che sui bastoncelli. Ma alla fine, il
campo visivo è quello che è determinato, in ultimo, dalle cellule gangliari, cioè o è centro-off o è centroon.
Questo che vuol dire? Noi fondamentalmente possiamo avere, sulla stessa cellula bipolare centro-on, più
coni; una cellula bipolare centro-on e una cellula bipolare centro-off, teoricamente, possono andare su
un’unica cellula gangliare che sarà o centro-on o centro-off. E’ lei che poi impone il sistema!
Questo però è un concetto un po’ più sofisticato. Il discorso viene fatto più che altro a livello di avere canali
centro-on e centro-off , come in effetti è, dalle cellule gangliari separate. Se io vado alla corteccia, avrò una
differente modalità di arrivo delle fibre M, delle fibre P, delle cellule delle fibre centro-on e delle cellule
delle fibre centro-off. Quindi diciamo che la cellula gangliare determina il centro-on o il centro-off.
Il mio discorso nasce da un concetto fondamentale: quando voi parlate di fovea, parlate della zona della
visione discriminata, cioè della massima nitidezza d’immagine.
Dire che “la zona è costituita esclusivamente da coni” è sbagliato: ci sono i coni, ma ci sono pure i
bastoncelli. E guardate che Kandel stesso cade in contraddizione. L’unico vero problema importante è che
la catena verticale è inclinata verso l’esterno, cioè la catena verticale non è a 90° sul piano delle cellule
dell’epitelio pigmentato, ma è inclinata all’esterno, quindi la luce va direttamente sul segmento esterno. Il
fotocettore è connesso alla cellula bipolare e alla cellula gangliare che si continua con le fibre nervose ma
questo sistema è inclinato, non è ortogonale come le altre zone, e questo permetterà all’asse luminoso di
arrivare direttamente sull’articolo esterno.
Quindi non diciamo che la fovea è il punto della visione discriminata perché ha più coni, perché vedi, io
posso foveare anche un oggetto in movimento, e per farlo io ho in ogni caso bisogno dei bastoncelli, perché
ho una forma che si sposta ed ho una visione grossolana: pur foveando, ho una visione che ha meno
dettagli di quanto non ne avrebbe se riguardasse un’immagine fissa.
[Studente: “Quindi la fovea ha meno strati?” Prof:”No, ha gli stessi strati, sono solo inclinati. Cioè, invece di
metter e tanti stecchini in posizione eretta, tu li inclini e la parte interna è quella che viene colpita subito
dalla luce.”].
[Studente:”Però professore, forse è l’immagine, però c’è una porzione di fovea che riceve più luce..” Prof:
Devi vederlo in 3d, qui c’è un piano solo. E in quel buco ad imbuto te ne fa vedere due soli ma in realtà sono
milioni…In buona sostanza, ci sono milioni di fotocettori che vengono colpiti dalla luce senza che questa
luce sia deviata dai corpi cellulari, perché confluisce tutta“].
[Studente: “Ma c’ è un maggior numero di fotocettori nella fovea?” Prof: “No, non è una macula! La
chiamano macula lutea perché è colorata, ma in effetti è una fovea, non c’è maggior densità recettoriale,
c’è solo inclinazione dell’asse verticale”].
Se torniamo adesso alle nostre caratteristiche di partenza, cominciamo, in base a quello che ha detto la
signorina, a capire un fatto fondamentale: cos’è il campo recettoriale?
Il campo recettoriale non è il campo di un bastoncello o di un cono, ma di un insieme n di coni o di
bastoncelli che possono centrare direttamente su una cellula gangliare. Diciamo che la convergenza dei coni
rispetto a quella dei bastoncelli, al contrario di quanto dicono molti testi, è minore, per cui abbiamo
un’immagine più nitida.
Ricordatevi sempre che la visione a colori non è una visione più bella, ma più ricca, perché la quantità di
sfumature presenti nell’arco della gamma di colori discriminati è enormemente maggiore di quella che va,
nella scala dei grigi,dal bianco estremo al nero: è almeno 11-12 volte superiore. Quindi è chiaro che noto
una serie di particolari che non noterei in bianco e nero.
Torniamo ai coni e bastoncelli. Ricordate che il cono ha una vera e propria continuità di membrana nei
dischi, mentre nel bastoncello i dischi sono proprio “fistulazioni”interne, uno splitting del bilayer, è lo strato
interno che si fistulizza, più o meno, con lo stesso meccanismo mitocondriale delle creste.
Tecnicamente, vengono prodotti circa 3 dischi ogni ora. Il neurone non va mai in apoptosi, è una cellula che
non va in ricambio cellulare quindi non troveremo delle cellule che serviranno poi a rinnovare le cellule che
ci sono, ma rinnovano sempre i dischi con una certa rapidità.
Il bastoncello ha una grande capacità di fotomoltiplicazione, recepisce una densità bassissima di energia, di
fotoni, per il semplice motivo che in una serie di 1000 fotoni disorientati,anche solo un fotone passa
attraverso il bastoncello. Nel cono è più difficile, perché la stretta forma a cono rende possibile che una
certa quantità di fotoni non colpisca nessun disco. Mentre per quanto riguarda i dischi dei bastoncelli, è
obbligatorio che un disco venga sempre colpito anche solo da un fotone. La capacità di fotomoltiplicazione
è notevolissima.
[Studente: “Un solo fotone è sufficiente per innescare il potenziale?” Prof: “ Lo innesca, perché ha una
grande capacità di fotomoltiplicazione”.Questo è dato dalla convergenza: tu hai più bastoncelli su una sola
cellula bipolare e alla fine più bastoncelli su una cellula gangliare].
I bastoncelli hanno una sensibilità elevata; la sensibilità dei coni è inferiore.
I primi hanno un’amplificazione elevata anche ai singoli fotoni, minore amplificazione, bassa risoluzione
temporale (sono poco indicati per la messa a fuoco).
I coni hanno alta risoluzione temporale, risposta rapida e sono necessari per osservare un oggetto in
movimento, che vedi bene alla luce, e male durante le ore notturne.
I bastoncelli sono più sensibili alla luce diffusa. I coni sono più sensibili alla luce diretta assialmente.
Abbiamo una bassa attività visiva dei bastoncelli,è presente nella fovea, ma in quantità minore rispetto ai
coni. Infatti se vai a vedere cosa dice Kandel, ti dirà che la fovea non è costituita solo da coni ma anche da
bastoncelli, in quantità minore. In ultimo, è acromatico, riceve solo la visione in bianco e nero.
Questo l’abbiamo fatto, la forma cis e la forma trans, la rodopsina che è sottoposta a stimolazione si divide
in opsina e retinale 11 cis.
[La rodopsina è una proteina di membrana con 7 domini transmembrana a α-elica. Si trova principalmente nelle cellule
a bastoncello della retina umana che permettono la vista in bianco e nero. Queste cellule hanno una forma allungata e
nella loro parte apicale hanno numerosi dischi di membrana con molte rodopsine, legate tramite una base di Schiff ad
un pigmento l'11-cis-retinale sensibile alla luce.
Nei Bastoncelli abbiamo inoltre numerosi canali per il Na + aperti in condizione di buio, perché nella cellula ci sono alte
concentrazioni di cGMP che tiene aperti i canali.
Quando la luce interagisce con l'11-cis-retinale, catalizza la sua conversione a tutto-trans-retinale, che determina un
cambiamento conformazionale nella "opsina" (parte proteica della rodopsina) trasformandola in opsina attiva. Questa
proteina attivata, attiva una seconda proteina, una proteina Gt, la trasducina, che a sua volta attiva una specifica GTP
Fosfodiesterasi, che scinde il cGMP in GMP e determina un abbassamento della concentrazione del cGMP: i canali per
il Na+ lungo la membrana della cellula a bastoncello si chiudono, si determina una variazione del potenziale di
membrana (iperpolarizzazione). La membrana essendo polarizzata non rilascia il GABA (γ-amminobutirrato, un
neurotrasmettitore inibitorio), in questo modo le cellule del nervo ottico non essendo inibite possono generare l'impulso
nervoso, che arriverà al nostro cervello tramite il nervo ottico (Wikipedia)].
La cosa che m’interessa è che voi sappiate che al buio avete una grande quantità di cGMP; il cGMP
direttamente si aggancia al versante interno dei canali voltaggio-dipendenti (Na+ soprattutto),
quindi mantenendo i canali aperti si mantiene una corrente di depolarizzazione che viene inibita dal
cGMP. L’11- tutto-trans riesce ad attivare, attraverso l’induzione di una Proteina G
fondamentalmente, la trasducina, una fosfodiesterasi specifica che esterifica il cGMP e ne riduce
quindi la concentrazione.
Al buio la massima quantità di cGMP tiene i canali Na+ aperti, quando arriva la luce il cGMP si
abbassa e i canali si chiudono, quindi vado in iperpolarizzazione. Questi canali quindi sono
fortemente legati al cGMP sul versante interno: non appena esterifico il cGMP (perché viene
attivata la fosfodiesterasi),abbassando la sua concentrazione automaticamente i canali Na+ si
chiudono e vado in iperpolarizzazione. Se ve lo ricordate, il passaggio intermedio che rende attivo il
cGMP è il passaggio della trasducina,il cGMP poi tiene aperto il canale. La fosfodiesterasi attiva
esterifica il cGMP, quest’ultimo non sarà quindi presente all’interno, non è più attivo sui canali e i
canali si chiudono. Siamo di fronte ad un meccanismo opposto a tutti gli altri meccanismi di
trasduzione, perché qui partiamo da una corrente di depolarizzazione in corso.
Ritorniamo al discorso di prima.
Cosa vuol dire campo recettoriale centro-on: vuol dire che ho una cellula gangliare che nel
momento in cui perviene la luce al centro del suo campo blocca i fotocettori che vanno in
iperpolarizzazione; non essendoci più il glutammato le cellule centro-on bipolari vengono attivate e
queste attivano la cellula gangliare centro-on. L’alone intorno al campo recettoriale centro-on è un
alone buio, scuro, perché ricordate che il vostro gioco di fotocettori è un gioco che esalta il
contrasto.
Ragionate: pigliate un muro bianco, guardatelo e ditemi cosa riconoscete. Niente!
Pigliate un muro decorato,scritto o con qualunque oggetto, ma che sia scuro…a quel punto
distinguo,percepisco, riconosco. Quindi il nostro sistema deve essere un sistema che gioca sul
contrasto luce-buio.
Cosa vuol dire centro-on? Non vuol dire solo “luce al centro”, ma “luce al centro e buio in
periferia”, sennò non funziona!
Con una gangliare io non piglio solo un fotocettore, piglio dei fotocettori che sono mirati su di te,
ma intorno io sto discriminando! Quindi avrò altri fotocettori, non posso parlare di campo
recettoriale di una bipolare o di un fotocettore, il fotocettore è sempre depolarizzato o
iperpolarizzato, è sempre buio o sempre luce; posso parlare di una gangliare, perché una gangliare
piglia un insieme e devo avere la luce al centro e buio alla periferia perché il sistema funzioni.
E la centro-off com‘è? Al contrario! E’ la cellula gangliare in cui il fotocettore non stimolato dalla
luce attiva delle cellule bipolari che hanno bisogno di un centro-off e quindi vanno alla cellula
centro-off..
Il centro buio deve avere un alone illuminato, sennò non funziona! Perché gioco sempre sui
contrasti, luce-buio, rosso-verde, giallo-blu…
[Studentessa: “Noi vediamo il bianco, ma è l’insieme di tutti i colori…” Prof: “Diciamo che la
lunghezza d’onda è la somma di tutte le lunghezze d’onda.” Studentessa: “Quindi il bianco lo
vediamo con i coni o con i bastoncelli?” Prof:”Il bianco lo vediamo con i coni, il contorno con i
bastoncelli. Se io vado a vedere qualcosa di bianco, per me per esempio il muro non ha contorni, ma
se io inserisco nel muro un quadro nero, questo ti fa capire perché in una foto mettiamo una cornice,
perché giochi sul contrasto. Se hai dei colori bui, questo capita anche in microscopia, metti un
riquadro bianco, perché in fondo è la stessa cosa avere un muro bianco e nero, non vedi niente…Il
bianco lo vedi come sovrapposizione di tutte le lunghezze d’onda; se io devo identificare ognuno di
voi, la mia visione bastoncellare mi mostra il vostro contorno, il contenuto lo mostrano i coni”]
[Studente: “Prof, ma la cellula gangliare, abbiamo il potenziale del recettore oppure…(l’audio è
poco chiaro) Prof:” No! Quando arriviamo alla cellula gangliare, il momento in cui abbiamo il cono
di emergenza dell’assone, subito dopo il cono di emergenza abbiamo il potenziale di integrazione,
d’innesco. Dalla gangliare in poi hai il potenziale, fondamentalmente l’insieme fotocettore-cellula
bipolare per me è il vero recettore, e posso avere un recettore centro-on e un recettore centro-off”]
[Studente: “Gangliare comunque somma…” Professore: Gangliare comunque somma, sì”].
[Studentessa: “Ma quindi il campo recettoriale centro-on, per esempio, è dato da quello della cellula
gangliare a cui però arrivano anche impulsi delle centro-off adiacenti!” Prof:”Esatto! Per capire
meglio questa cosa dobbiamo risalire alla mancanza di 3 tipi di cellule: abbiamo le cellule del
Muller, cellule in verticale, che nutrono la retina, sono estese per tutta la retina tranne che a livello
dell’epitelio pigmentato; abbiamo poi le cellule amacrine e le cellule orizzontali.
Le cellule orizzontali sono cellule poste tra più articoli interni, o meglio, tra più espansioni
sinaptiche dei fotocettori. Per esempio, mettiamo che si trovino tra due fotocettori, e che abbiamo
una centro-on da un lato e una centro-off dall’altro(possono essere anche a distanza): la cellula
bipolare ha bisogno del glutammato per essere attivata, quindi il glutammato attiva la cellula
orizzontale. La cellula orizzontale emette,invece, un inibitore, un iperpolarizzatore sull’altro
fotocettore [Studente:”Inibizione centro-periferia..]
Se io ho qui un fotocettore e qua un altro fotocettore, su questo fotocettore mi arriva
improvvisamente la luce, questo recettore non ha più glutammato. Non avendo glutammato, non
attiva più la cellula orizzontale; non attivando la cellula orizzontale, quest’ultima non inibisce
l’altro fotocettore. Quindi, se io qua ho luce, vado in iperpolarizzazione, quindi la centro on parte, la
centro off accanto si blocca, questo non iperpolarizza più, l’altro scarica, quindi si blocca la centroon dall’altra parte e attiva la centro-off di là. Se io qua ho bianco, qua ho nero.
[Studente: “Ma la centro-off non sarebbe già disattivata all’arrivo della luce?”]
Qua mi arriva la luce, ho il mio fotocettore, diventa iperpolarizzato, la mia centro-off è bloccata e la
centro-on è attiva. Andiamo accanto: questo iperpolarizza, nel momento in cui non iperpolarizza
più, non è quindi stimolato, questo fotocettore è libero di scaricare; se scarica, blocca le centro-on e
attiva le centro-off. Quindi hai un maggior contrasto centro-periferia, l’inibizione centro-periferia.
Il campo non può essere solo luminoso o solo buio, deve esserci un centro luminoso e un alone
buio, o un centro buio e un alone luminoso. Ora funziona perché non appena io iperpolarizzo, non
comunico più l’attivazione della cellula orizzontale, la cellula orizzontale non più attivata mi lascia
libero un altro fotocettore. Se io ho un fotocettore particolarmente attivo, ne ho un altro meno
attivo.
[Studentessa: “Ma questo all’interno dello stesso campo recettoriale o riguarda anche gli
altri?Prof:”No, nello stesso campo”].
Sul punto in cui ho la massimo quantità di luce o di colore, ho la massima iperpolarizzazione, la
massima attivazione dei centri-on e la massima inibizione dei centri-off. Nella parte periferica ne
arriverà di meno, quindi queste sono meno stimolate, queste mi stanno scaricando. Se fossero
iperpolarizzate non scaricherebbero più, ma se blocco l’inibizione queste periferiche
scaricano,inibiscono le centro-on e attivano le centro-off.
Quindi qui ho massima attivazione delle centro-off, qui delle centro-on, qui ho luce, qui no.
Se io ho una centro-on e una centro-off, mettetele insieme a più livelli, una viene scaricata e quindi
viene inibita l’orizzontale. Facciamo al contrario:qui ho buio quindi ho glutammato. Il glutammato
attiva le centro-off, inibisce le centro-on ma attiva anche l’orizzontale. L’orizzontale inibisce i
fotocettori accanto, ma se il fotocettore accanto è inibito cosa accadrà? Ho il massimo centro-on e
niente centro-off. Ed ecco che ho realizzato l’inibizione centro-periferia. Ho il massimo contrasto:
vedo bene se ho contrasto, vedo male se ho uniformità.
Pensate a quando vi trovate al sole,al mare, e guardate la sabbia, intanto vi brucia tutti i bastoncelli
che avete, avete senso di bruciore agli occhi e dovete chiudere gli occhi per alcuni secondi: in quei
secondi si rigenerano i bastoncelli ma, in ogni caso, non avete risoluzione perché vedete una massa
uniforme e tutti i fotocettori partono, avrete tutti i centri-on attivi e non vedrete più niente, solo un
massa di luce incredibile.
Affinchè la retina possa funzionare bisogna che ci sia contrasto: se da un lato favorisco le centroon, dall’altro devo esaurire le centro-off.
La cellula è depolarizzata: chi parte?Le centro-off; ma parte contemporaneamente la cellula
orizzontale, che blocca l’altro fotocettore. Non c’è più glutammato, quindi chi parte? La centro-on.
Il campo recettoriale sarà al centro luminoso, in periferia buio, o al contrario.
Alla stessa maniera lavorano le amacrine, né più nè meno. Lo stesso meccanismo che mettete in
funzione tra fotocettori e orizzontali, lo mettete in funzione tra bipolari e amacrine.
Se io ho una bipolare centro-on, inibirà l’amacrina che bloccherà la centro-off dell’altro lato e
viceversa.
Quindi orizzontali e amacrine lavorano alla stessa maniera!
L’occhio è un fine analizzatore che gioca sul contrasto. Questo ce lo portiamo a livello dei
genicolati, della corteccia, tutto il sistema gioca sul contrasto!
Se io qui ho una catena di fotocettori bipolare e gangliare, e poi ho una catena lì e lì, se io ho la
massima stimolazione qui mi attiva questo sistema, questa quindi è una stimolazione diretta. Ma lì
funziona pure: la quantità di luce che mi sta arrivando sta funzionando attraverso le orizzontali
giocando con l’inibizione centro-periferia, in modo da completare l’immagine. Il segnale va sia per
via diretta che laterale. Cosa vuol dire diretta? Che diretta è la catena cellulare (quella che abbiamo
studiato, fotocettori-bipolari-gangliari). Cos’è la via laterale? Le cellule orizzontali e amacrine, il
sistema che mi serve ad amplificare il contrasto.
Se io avrò una zona di buio, con il contrasto percepirò anche la luce accanto; se io avrò una zona di
luce, percepirò il buio accanto, sennò avrò o tutto buio o tutto luminoso. E’ una via che esalta il
contrasto.
Quella che ci sta facendo capire di più la corteccia,devo dire che è la vista: per un soggetto,
parliamoci chiaro, la sordità è terribile, la mancanza di un braccio è terribile, ma la cosa peggiore è
la cecità. Ho sempre detto che i mali sociali per un soggetto sono la cecità o la perdita della
motricità..poi, se li perdi tutti e due sei proprio sfortunato…
Vediamo in quanti modi si può formulare la domanda:
Retina
Le cellule della retina (sono 7: epitelio pigmentato, fotocettori, cellule bipolari e gangliari,
cellule del Muller, amacrini e orizzontali)
Strati della retina: sono 10!
Meccanismo funzionale della retina: campo recettoriale, centro-on, centro-off,ecc.
Parliamo delle gangliari, anche se io un discorso ve l’avevo già fatto: avevamo parlato di X,Y, W.
Abbiamo quindi 3 tipi di gangliari, anche se Kandel, come moltissimi testi di neuroscienze, si
sofferma sempre su 2: le M (Magnicellulari o Y) e le P (Parvicellulari o X).
Le X sono quelle della discriminazione massima e dell’analisi, sono quelle maggiormente legate ai
coni.
Le M (o le Y)sono quelle dei dettagli grossolani di un oggetto; è come se dicessimo M epicritica, P
protopatica. E in più, il soggetto in movimento.
Le W sono quelle di cui non si conosce ancora bene la struttura, sono grandi, hanno una notevole
arborizzazione denditica, rispondono all’intensità della luce, ma spesso non sono tenute in
considerazione.
Le chiameremo M e P perché? Nel nucleo del genicolato abbiamo 6 strati: 2 sono magnicellulari e
4 sono parvicellulari.
Se due sono magnicellulari, cosa vi aspettate che arrivino? Le gangliari M o P? Le M!
Se gli altri 4 sono parvicellulari cosa vi aspettate che arrivino? Le P!
Queste arriveranno dopo il chiasma ottico, in cui c’è l’incrocio esclusivo delle fibre nasali, le fibre
temporali rimangono omolaterali. Le fibre dell’emiretina nasale di sinistra vanno a destra mentre le
fibre dell’emiretina nasale di destra vanno a sinistra. Le fibre temporali di destra rimangono a
destra, le fibre temporali di sinistra restano a sinistra.
Questo nasce soprattutto perché il campo visivo è capovolto, cioè quello che percepisco per le leggi
dell’ottica è capovolto.
A me però interessa che voi capiate che al genicolato arrivano le M omolaterali e le M contro
laterali, le P omolaterali e le P contro laterali.
In sostanza avremo un M omolaterale e un M contro laterale; 2 P omolaterali e 2 P contro laterali.
Non vi “sbarruate!” (Sbarruare…famoso italiano dell’800! Anzi, eravamo nel 700 quando papà mi
spiegava queste cose!)
Stavamo dicendo che:
- il primo strato del corpo genicolato è magnicellulare, e prenderà le fibre magnicellulari contro
laterali;
-il secondo strato prenderà le fibre magnicellulari omolaterali;
-il terzo strato prenderà le fibre parvicellulari omolaterali;
-il quarto strato prenderà le fibre parvicellulari contro laterali;
- il quinto strato prenderà le fibre parvicellulari omolaterali;
-il sesto strato prenderà le fibre parvicellulari controlaterali .
Per ognuno di questi strati avrete una ricostruzione retino topica: abbiamo già 6 mappe
retinotopiche.
Le W non arrivano qui, passano “o rittu!” Pigliano il primo svincolo a destra, scendono al braccio
congiuntivo, vanno al corpo quadrigemello superiore e da lì arrivano ai nuclei dell’area
pretettale.
Il 100% delle P arriva al corpi genicolato; ci andrà anche l’80% delle M (M sensibili agli oggetti in
movimento), mentre il 20 % restante andrà ai corpi quadrigemelli superiori, che coordinano il
movimento oculomotorio. Le W vanno al riflesso viscerale. Se mentre sto guardando te vedo una
mosca, muovo gli occhi e sono sicuro di averlo piazzato, ma non lo piazzo perché quando ho
calcolato l’angolo di apertura l’oggetto era lì ma poi si è mosso. Il 100% delle W (o fibre pupillari)
vanno ai nuclei dell’area pretettale, in cui i centri-on attiveranno i riflessi di ipercostrizione, i
centri-off attiveranno il riflesso di iperdilatazione.
Flavia D’Andrea