STRUTTURA E
PROCESSI VITALI
NEGLI ANIMALI
settima edizione
eldra p. solomon
University of South Florida, Tampa
Hillsborough Community College
charles e. martin
Rutgers University
diana w. martin
Rutgers University
linda r. berg
University of Maryland, College Park
St. Petersburg College
Edizione italiana a cura di:
barbara costa
Università degli Studi di Pisa
stefano parmigiani
Università degli Studi di Parma
giuseppe fiermonte
paola palanza
emilio hirsch
giorgio scarì
Università degli Studi di Bari
Università degli Studi di Torino
Università degli Studi di Parma
Università degli Studi di Milano
Titolo originale:
Eldra P. Solomon, Charles E. Martin, Diana W. Martin, Linda R. Berg
Biology – X edizione
Copyright © 2015, 2011, Cengage Learning
Struttura e processi vitali negli animali – VII edizione
Copyright © 2017 EdiSES s.r.l. – Napoli
(estratto da Biologia VII edizione, Copyright © 2017, EdiSES s.r.l. – Napoli)
9 8 7 6 5 4 3 2 1 0
2022 2021 2020 2019 2018 2017
Le cifre sulla destra indicano il numero e l’anno dell’ultima ristampa effettuata
A norma di legge è vietata la riproduzione,
anche parziale, del presente volume o parte
di esso con qualsiasi mezzo.
L’Editore
Fotocomposizione: Grafic&Design – Napoli
Stampato presso la:
Tipolitografia Petruzzi S.r.l.
Via Venturelli, 7/B
06012 Città di Castello (PG)
per conto della
EdiSES S.r.l. – Piazza Dante, 89 – Napoli
http://www.edises.it e-mail: [email protected]
ISBN 978-88-7959-949-8
Sommario
parte sette: Struttura e processi vitali negli animali
39
Struttura e funzioni degli animali: un’introduzione 815
40
Protezione, sostegno e movimento 836
41
Segnalazione neurale 854
42
Regolazione neurale 876
43
Sistemi sensoriali 905
44
Trasporto interno 930
45
Il sistema immunitario: la difesa interna 956
46
Scambi gassosi 985
47
Elaborazione del cibo e nutrizione 1004
48
Osmoregolazione ed eliminazione dei rifiuti metabolici 1026
49
Regolazione endocrina 1044
50
La riproduzione 1068
51
Sviluppo animale 1098
52
Comportamento animale 1118
Risposte R-1
Indice analitico I-1
III
Autori
Eldra P. Solomon ha scritto
molti libri d’avanguardia di
Biologia, Anatomia umana e
Fisiologia. I suoi libri sono
stati tradotti in più di dieci
lingue. Ha ricevuto un M.S.
all’Università della Florida e
un M.A. e Ph.D. all’Università della Florida del Sud. È
professore e membro della
Facoltà dell’Università della
Florida del Sud; insegna Biologia da 20 anni.
È una biopsicologa e una
biologa. Come psicologa è
specializzata in salute mentale, disordini da stress post–
traumatico, disordini dissociativi e gestione dello stress.
La sua ricerca si è focalizzata
sulle correlazioni tra lo stress,
le emozioni e la salute.
Ha presentato i suoi lavori in molte conferenze nazionali e internazionali. È stata citata più di 30 volte in attualissime pubblicazioni, incluso Who’s Who in America,
Who’s Who in Science and Engineering, Who’s Who in Medicine and Healthcare, Who’s
Who in American Education,
Who’s Who of American Women e Who’s Who in the
World.
Charles E. Martin è professore
emerito di Biologia cellulare
e Neuroscienze presso l’Università Rutgers. Ha ricevuto
un Ph.D. in Genetica dall’Università statale della Florida
e ha condotto un post-dotto­
rato in Genetica e Biologia
delle membrane presso l’Università del Texas di Austin.
Nel corso della sua carriera a
Rutgers ha insegnato Biologia generale così come ha curato corsi di Genetica e Biologia cellulare molecolare per
laureandi e laureati. Professore premiato per più di 30
anni, nel 2011 è stato nominato Professore dell’anno dal­
la “Molecular Biosciences Gra­
duate Student Association”.
La sua ricerca sulla regolazione genica dei sistemi
enzimatici delle proteine di
membrana in lievito e altri
funghi illustra la natura interdisciplinare delle scienze
della vita. È molto fiero delle
generazioni di laureandi, laureati e studenti di post-dottorato che hanno contribuito
alla sua ricerca e hanno iniziato floride carriere. Continua a essere scelto per l’insegnamento ed è grato per
le opportunità in tale ambito e nell’ambito della ricerca in quest’era che continua
ad essere la più eccitante delle scienze biologiche.
Diana W. Martin è professore
emerito e ex-direttore di Biologia Generale, Divisione del­­
le Scienze della vita, presso
Università Rutgers. Ha ottenuto un M.S. all’Università della Florida, dove ha studiato i cromosomi di specie
di piante correlate per comprendere le loro correlazioni evolutive. Ha ricevuto un
Ph.D. all’Università del Texas
a Austin, dove ha studiato la
genetica della Drosophila melanogaster e poi ha svolto attività di ricerca post-dottorato presso l’Università di Princeton.
Ha curato corsi di Biologia generale alla Rutgers per
più di 30 anni e ha iniziato a
scrivere libri dal 1988. È molto soddisfatta della sua scelta
di insegnare biologia al college poiché questa carriera
le permette di condividere il
suo entusiasmo per tutti gli
aspetti della biologia.
Linda R. Berg è vincitrice di un
premio per l’insegnamento e
autrice di libri. Ha ricevuto
un B.S. per l’insegnamento
delle Scienze, un M.S. in Botanica e un Ph.D. in Fisiologia vegetale dall’Università
del Maryland. La sua ricerca è
focalizzata sulle implicazioni
evolutive della biosintesi degli steroidi in vari organismi.
Ha insegnato per 17 anni presso l’Università del
Maryland e per 8 anni presso il College St. Petersburg in
Florida. Durante la sua carriera ha curato corsi introduttivi in Biologia, Botanica e Scienze ambientali a migliaia di studenti. All’Università del Maryland ha ricevuto numerosi premi per l’insegnamento e l’organizzazione.
Ha anche ricevuto riconoscimenti nazionali e regionali, incluso quello nazionale per l’innovazione dell’insegnamento
delle Scienze, quello da parte del “National Science Teachers Association Award for
Innovations in College Science
Teaching”, il “Nation’s Capital
Area Disabled Student Services Award” e il “Washington
Academy of Sciences Award in
University Science Teaching”.
Durante la sua carriera di
redattrice di libri scientifici,
è stata autrice e coautrice di
molti libri di testo per il college. Il suo modo di scrivere
riflette il suo stile di insegnamento e il suo amore per la
scienza.
Struttura e funzioni
degli animali: un’introduzione
I
gruppi animali sono estremamente diversificati e presenta-
39
no strutture corporee radicalmente differenti. Ad esempio,
si consideri quanto siano diversi l’elefante e l’uccello, non
solo nelle dimensioni, ma anche nella forma corporea e nello stile di vita (vedi fotografia). Tuttavia, nonostante le notevoli
differenze, tutti i gruppi animali condividono numerose caratteristiche, una delle quali è rappresentata dalle dimensioni corporee relativamente grandi.
Perché la maggior parte degli animali è più grande dei batteri, dei protisti e dei funghi? Molti biologi ritengono che la riovvero ai ruoli funzionali delle specie all’interno di una comunità. Mentre gli animali si evolvevano, altri organismi avevano
già occupato la maggior parte delle nicchie ecologiche disponibili. Le nuove specie, se volevano avere il predominio, dovevano allontanare un organismo dalla nicchia che aveva occupato
con successo oppure adattarsi a una nuova nicchia. La conquista di una nuova nicchia richiedeva un adattamento corporeo,
spesso basato su dimensioni maggiori. Poiché i predatori sono
in genere più grandi delle loro prede, una dimensione maggiore offriva più opportunità di procacciarsi il cibo.
Per poter essere più grandi dei competitori batterici e protistici, gli animali dovevano quindi essere pluricellulari. Si ricordi che le dimensioni di una singola cellula sono limitate dal
© Francois Gagnon/Shutterstock.com
sposta a questa domanda sia collegata alle nicchie ecologiche,
Dimensione corporea maggiore non significa cellule più grandi. Le cellule di un elefante adulto, quelle dell’elefantino e dell’uccello posato sulla
testa dell’elefante adulto hanno all’incirca tutte le stesse dimensioni.
L’elefante adulto è più grande dell’uccello perché i suoi geni specificano
che il suo corpo deve essere costituito da un maggior numero di cellule. Le cellule dell’elefantino continueranno a moltiplicarsi fino a quando
esso raggiungerà le dimensioni dell’adulto.
rapporto tra la sua superficie (membrana plasmatica) e il suo
volume (vedi Capitolo 4). La membrana plasmatica deve es-
concetti chiave
sere sufficientemente grande, in rapporto al volume cellulare,
da permettere il passaggio di materiale dentro e fuori la cellula, in modo da mantenere le condizioni necessarie per la vita.
In un animale pluricellulare, ogni cellula ha un rapporto superficie-volume sufficientemente grande da regolare efficacemente l’ambiente interno. Le singole cellule possono vivere, morire
ed essere rimpiazzate mentre l’organismo continua a conservarsi e a crescere. Il numero di cellule, non la loro grandezza, è
responsabile delle dimensioni di un organismo.
Negli organismi unicellulari, come i batteri e i protisti, la
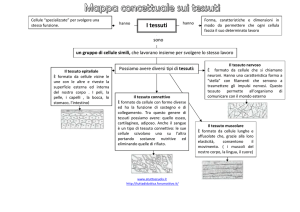
39.1 Le cellule formano i tessuti, vari tipi di tessuti formano gli organi, i tessuti e gli organi che lavorano insieme costituiscono
gli apparati. I principali tipi di tessuti in un animale sono quelli epiteliale, connettivo, muscolare e nervoso.
39.2 I meccanismi omeostatici sono principalmente sistemi di controllo a feedback negativo che mantengono un ambiente interno relativamente stabile.
39.3 La termoregolazione è un processo omeostatico che mantiene
la temperatura del corpo costante nonostante variazioni esterne (o interne) della temperatura.
singola cellula deve svolgere tutte le attività necessarie per la
vita. Si ricordi che i piccoli, semplici organismi unicellulari dipendono dalla diffusione per molti processi vitali, compresi gli
scambi gassosi e l’eliminazione dei prodotti metabolici di scarto. Uno dei motivi per cui possono essere piccoli è che non
hanno bisogno di complessi apparati.
In questo capitolo concentreremo la nostra attenzione sulla struttura e le funzioni di base degli animali. L’anatomia è lo
studio della struttura di un organismo. La fisiologia è lo studio
delle modalità di funzionamento dell’organismo. Si ricordi che,
nei sistemi biologici, struttura e funzione sono strettamen815
te correlate a ogni livello di organizzazione. Molteplici esempi di
questo principio basilare saranno riportati man mano che descriveremo i tipi e le funzioni dei tessuti e degli apparati degli animali. Inoltre, discuteremo l’importante concetto dell’omeostasi, usando come esempio la regolazione della temperatura corporea. Nei
capitoli successivi discuteremo come gli animali svolgono i processi vitali e come gli apparati di un animale complesso cooperano per mantenere l’omeostasi dell’organismo nel suo insieme.
39.1 TESSUTI, ORGANI E APPARATI
obiettivi di apprendimento
1 Confrontare la struttura e la funzione dei quattro tipi principali di tessuti animali: epiteliale, connettivo, muscolare e
nervoso.
2 Confrontare i principali tipi di tessuto epiteliale e descriverne le funzioni.
3 Confrontare i principali tipi di tessuto connettivo e descriverne le funzioni.
4 Mettere a confronto i tre tipi di tessuto muscolare e descriverne le funzioni.
5 Mettere in relazione la struttura del neurone con la sua
funzione.
6 Descrivere brevemente gli apparati di un mammifero e
riassumere le funzioni di ciascun apparato.
In un organismo pluricellulare, le cellule sono specializzate per
assolvere a compiti specifici. Dal Capitolo 1, si ricordi che le
cellule si organizzano a formare tessuti e i tessuti si associano
a formare organi come il cuore o lo stomaco. Gruppi di tessuti e organi formano gli apparati di un organismo complesso.
Miliardi di cellule possono essere organizzati a formare tessuti, organi e apparati di una farfalla, di un coccodrillo o di un
elefante.
Un tessuto consiste di un gruppo di cellule simili, strettamente associate, che possono svolgere funzioni specifiche.
I biologi classificano i tessuti animali in epiteliale, connettivo, muscolare e nervoso. La classificazione dei tessuti si basa sulla loro origine e sulla loro struttura. Ogni tipo di tessuto
è composto da cellule di grandezza, forma e disposizione caratteristiche; ogni tipo di tessuto è specializzato per svolgere
una determinata funzione o un gruppo di funzioni. Ad esempio, alcuni tessuti sono specializzati nel trasporto di materiali, altri nella contrazione, permettendo agli organismi di muoversi. Altri ancora secernono ormoni che regolano i processi
metabolici. Struttura e funzione sono strettamente correlate a
ogni livello organizzativo. Nella discussione sui vari tipi di tessuti, è sempre importante notare la correlazione tra la struttura e la funzione di ciascuno e come i sistemi biologici interagiscano continuamente.
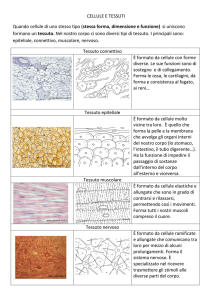
I tessuti epiteliali rivestono il corpo
e tappezzano le sue cavità
Il tessuto epiteliale (comunemente chiamato epitelio) è costituito da cellule strettamente adese che formano uno strato
continuo. Una superficie di questo strato è esposta, perché riveste il corpo (strato esterno della cute) o delimita una cavità,
816
CAPITOLO 39
come il lume (la cavità di un organo cavo) dell’intestino. L’altra superficie dello strato epiteliale è attaccata al tessuto sottostante tramite una membrana basale acellulare, composta
da fibre sottili e materiale polisaccaridico prodotto dalle cellule epiteliali.
Il tessuto epiteliale forma lo strato superficiale della cute e
il rivestimento dei tratti digerente, respiratorio, escretore e riproduttivo. Di conseguenza, tutto ciò che entra o esce dal corpo deve attraversare almeno uno strato di epitelio. Il cibo che
viene portato alla bocca e inghiottito non è realmente “all’interno” del corpo fino a quando non viene assorbito dall’epitelio
intestinale ed entra nel sangue. La permeabilità dei vari epiteli
regola ampiamente lo scambio di sostanze fra le differenti parti del corpo, come pure tra l’organismo e l’ambiente esterno.
Il tessuto epiteliale assolve a un gran numero di funzioni,
tra cui la protezione, l’assorbimento, la secrezione e la ricezione sensoriale. Lo strato epiteliale della cute, l’epidermide, riveste tutta la superficie corporea e la protegge da danni meccanici, chimici e batterici e dalla perdita di fluidi. Il tessuto epiteliale che riveste internamente il tratto digerente assorbe materiale nutritivo e acqua. Alcune cellule epiteliali sono organizzate in ghiandole che secernono prodotti come ormoni, enzimi o sudore. Altre cellule epiteliali sono recettori sensitivi e ricevono informazioni dall’ambiente: ad esempio, le cellule epiteliali delle papille gustative e del naso sono specializzate come recettori chimici.
La TABELLA 39-1 illustra i principali tipi di tessuto epiteliale, indica la loro localizzazione nell’organismo e descrive le loro funzioni (pagine 818–819). In base alla forma, possono essere distinti tre tipi di cellule epiteliali. Le cellule epiteliali pavimentose sono sottili e appiattite come le piastrelle di un mosaico. L’epitelio pavimentoso semplice riveste le pareti dei vasi
sanguigni e degli alveoli polmonari. Le cellule epiteliali cubiche sono piccoli cilindri che, visti di lato, possono assomigliare
a dei dadi; generalmente, ogni cellula cubica ha una forma esagonale in sezione trasversale e costituisce un ottaedro. L’epitelio cubico semplice riveste le pareti dei tubuli renali.
Le cellule epiteliali cilindriche assomigliano a sottili colonne o cilindri, se osservate di lato. Il nucleo è normalmente in
posizione basale. Viste dall’alto o in sezione trasversale, queste
cellule spesso appaiono esagonali. Sulla sua superficie libera,
una cellula cilindrica può essere provvista di ciglia, che si muovono in modo coordinato, facilitando il movimento di materiale sulla superficie del tessuto. La maggior parte del tratto respiratorio superiore è rivestita da un epitelio ciliato che rimuove particelle di polvere e altro materiale estraneo dai polmoni.
Il tessuto epiteliale può essere classificato anche in base al
numero di strati. L’epitelio semplice è composto da un singolo strato di cellule. È generalmente localizzato dove le sostanze vengono secrete, escrete e assorbite, o dove il materiale può
diffondere facilmente attraverso esso. Per esempio, un epitelio
pavimentoso semplice riveste gli alveoli polmonari. La struttura di questo sottile tessuto è adattata perfettamente a consentire la diffusione dei gas attraverso gli alveoli.
L’epitelio stratificato, composto da due o più strati, protegge i tessuti sottostanti. Per esempio, l’epitelio pavimentoso stratificato che costituisce lo strato esterno della pelle si desquama continuamente per le normali usure e lacerazioni, per
cui deve anche rigenerarsi continuamente. Le cellule dell’epitelio pseudostratificato appaiono falsamente disposte su più
strati. Sebbene tutte le cellule che lo compongono aderiscano alla membrana basale, non tutte arrivano alla superficie del
tessuto, dando quindi l’impressione di un tessuto costituito da
due o più strati di cellule. Una parte delle vie respiratorie è rivestita da un epitelio pseudostratificato provvisto di ciglia.
I rivestimenti dei vasi sanguigni e linfatici hanno una origine embrionale diversa rispetto al “vero” epitelio e vengono
denominati endoteli. Tuttavia, queste cellule sono strutturalmente simili alle cellule epiteliali pavimentose e possono essere comprese in questa categoria.
Le ghiandole sono costituite da cellule
epiteliali
Una ghiandola consiste di una o più cellule epiteliali specializzate nella produzione e nella secrezione di prodotti come sudore, latte, muco, cerume, saliva, ormoni o enzimi (FIG. 39-1).
Il tessuto epiteliale che riveste le cavità e i condotti del corpo
contiene spesso cellule specializzate nella secrezione di muco,
chiamate cellule caliciformi: il muco lubrifica queste superfici, offre protezione e facilita il movimento dei materiali.
Le ghiandole possono essere classificate come esocrine o
endocrine. Le ghiandole esocrine, come le cellule caliciformi e le ghiandole sudoripare, secernono i loro prodotti sulla
superficie epiteliale libera, generalmente attraverso un dotto.
Le ghiandole endocrine sono prive di dotti e rilasciano i loro
prodotti, denominati ormoni, nel fluido interstiziale (fluido
tissutale) o nel sangue; da qui, essi vengono poi trasportati alle altre parti del corpo dal sistema circolatorio. (Le ghiandole
endocrine sono discusse nel Capitolo 49).
Le cellule epiteliali formano membrane
Una membrana epiteliale consiste di una lamina di tessuto epiteliale e di uno strato sottostante di tessuto connettivo; alcuni esempi sono rappresentati dalle membrane mucose e sierose. Una membrana mucosa, o mucosa, riveste una cavità del
corpo che comunica con l’esterno, come il tratto digerente o
respiratorio. Le cellule caliciformi dello strato epiteliale secernono muco, che lubrifica il tessuto e lo protegge dalla disidratazione.
Una membrana sierosa ricopre una cavità del corpo che
non comunica con l’esterno ed è costituita da un epitelio pavimentoso semplice sopra uno strato sottile di tessuto connettivo lasso. Questo tipo di membrana secerne fluidi nella cavità
che riveste. Alcune tra le più comuni membrane sierose sono
la membrana pleurica, che riveste la cavità pleurica attorno ai
polmoni, e la membrana pericardica, che riveste la cavità pericardica intorno al cuore.
I tessuti connettivi sostengono
altre strutture del corpo
Quasi tutti gli organi del corpo hanno una struttura interna di
tessuto connettivo che li sostiene e li protegge. A confronto
con i tessuti epiteliali, i tessuti connettivi contengono relativamente poche cellule, immerse in un’abbondante sostanza intercellulare costituita da fibre microscopiche simili a fili sparse in una matrice allo stato di gel costituita da polisaccaridi secreti dalle cellule. La natura e la funzione di ogni tipo di tessuto connettivo sono determinate in parte dalla struttura e dalle
proprietà della sostanza intercellulare.
Ghiandole unicellulari
(cellule caliciformi)
Membrana basale
(a) Cellule caliciformi.
Sono ghiandole
unicellulari che
secernono muco.
(b) Ghiandole sudoripare.
Sono ghiandole semplici
con tubuli a gomitolo. Le loro
pareti sono costituite da un
epitelio cubico semplice.
(c) Ghiandole salivari
parotidee. Le ghiandole
composte, come le
parotidi, hanno dotti
ramificati.
Figura 39-1 Ghiandole
Una ghiandola consiste di una o più cellule epiteliali.
Il tessuto connettivo contiene generalmente tre tipi di fibre: collagene, elastiche e reticolari. Le fibre collagene, le più
numerose, sono composte dai collageni, un gruppo di proteine fibrose presenti in tutti gli animali (vedi Fig. 3-23b). I collageni sono le proteine più abbondanti nei mammiferi, costituendo circa il 25% della loro massa proteica totale. Il collagene è un materiale molto resistente (la carne è resistente a causa
del suo contenuto di collagene). La forza di trazione (capacità
di essere stirate senza rompersi) delle fibre collagene è paragonabile a quella dell’acciaio. Le fibre collagene sono ondulate e
flessibili e questo permette loro di rimanere intatte quando il
tessuto è sottoposto a stiramento.
Le fibre elastiche si ramificano e si anastomizzano per formare un reticolo. Esse possono essere stirate se sottoposte a
una forza e, quando questa viene rimossa, possono tornare alla loro forma e dimensione originali (come un elastico teso).
Le fibre elastiche, costituite dalla proteina elastina, sono un
importante componente delle strutture soggette a stiramento.
Le fibre reticolari sono fibre molto sottili e ramificate che
formano un delicato reticolo, il quale salda i tessuti connettivi
con i tessuti circostanti. Le fibre reticolari sono composte da
collagene e da alcune glicoproteine.
Le cellule dei vari tipi di tessuto connettivo differiscono
per forma e struttura e per i tipi di fibre e matrice che secerno-
Struttura e funzioni degli animali: un’introduzione
817
TABELLA 39-1
Tessuti epiteliali
Nuclei delle cellule epiteliali pavimentose
Epitelio pavimentoso semplice
Principali localizzazioni
Alveoli polmonari; rivestimento interno dei vasi
sanguigni
Ed Reschke/Getty Images
Funzioni
Passaggio di materiale dove non occorre protezione e dove la diffusione è la principale forma di
trasporto
Descrizione e commenti
Le cellule sono appiattite e disposte su singolo
strato
25 mm
Immagine al MO di epitelio pavimentoso semplice.
Nuclei di cellule epiteliali cubiche
Lume del tubulo
Epitelio cubico semplice
Principali localizzazioni
Rivestimento interno dei tubuli renali; dotti
ghiandolari
Funzioni
Secrezione e assorbimento
Ed Reschke/Peter Arnold/Getty Images
Descrizione e commenti
Singolo strato di cellule; la fotografia al MO
mostra sezioni trasversali di tubuli; vista di lato,
ogni cellula sembra un piccolo cilindro; alcune
hanno microvilli per l’assorbimento
25 mm
Immagine al MO di epitelio cubico semplice.
Cellula caliciforme
Nuclei di cellule cilindriche
Epitelio cilindrico semplice
Ed Reschke/Getty Images
Principali localizzazioni
Rivestimento interno della maggior parte del
tratto digerente e della parte superiore del tratto
respiratorio
Funzioni
Secrezione, soprattutto di muco; assorbimento;
protezione; movimento di strati di muco
Descrizione e commenti
Singolo strato di cellule cilindriche; complesso di
Golgi molto sviluppato; spesso ciliato; le cellule
caliciformi secernono muco
25 mm
Immagine al MO di epitelio cilindrico semplice.
Continua
818
CAPITOLO 39
TABELLA 39-1
Tessuti epiteliali (continua)
Epitelio pavimentoso stratificato
Principali localizzazioni
Cute; rivestimento interno della bocca; rivestimento interno
della vagina
Funzioni
Solo di protezione; poco o nullo l’assorbimento o il transito di
materiale; strato superficiale continuamente eliminato e rimpiazzato da quello sottostante
Descrizione e commenti
Diversi strati di cellule, in cui solo quelle più profonde sono
cilindriche e metabolicamente attive; la divisione delle cellule
profonde causa lo spostamento verso la superficie di quelle più
vecchie, che cominciano ad appiattirsi durante il movimento
Ed Reschke/Getty Images
Nuclei
50 mm
Immagine al MO di epitelio pavimentoso stratificato.
Epitelio pseudostratificato
Ciglia
Cellule epiteliali
Nuclei
Principali localizzazioni
Alcuni tratti dell’apparato respiratorio; dotti di molte ghiandole
Funzioni
Secrezione; protezione; movimento dello strato di muco
Descrizione e commenti
Ciliato, secernente muco o con microvilli; simile all’epitelio
cilindrico, tranne per il fatto che le cellule non sono tutte della
stessa altezza. Pertanto, sebbene tutte le cellule siano attaccate alla stessa membrana basale, il tessuto sembra essere
stratificato
Ed Reschke/Getty Images
Membrana basale
25 mm
Immagine al MO di epitelio cilindrico
pseudostratificato ciliato.
no. I fibroblasti sono le cellule del tessuto connettivo che producono le fibre e i complessi di proteine e carboidrati della matrice. Essi rilasciano nello spazio extracellulare le componenti
proteiche, che si organizzano a formare le fibre. Queste cellule
sono attive specialmente durante lo sviluppo tissutale e sono
importanti nella guarigione delle ferite. Nel tessuto adulto, il
numero di fibroblasti diminuisce ed essi cominciano a perdere la loro attività biosintetica. I macrofagi, le cellule spazzine
del corpo, si muovono generalmente attraverso il tessuto con-
nettivo, facendo pulizia di detriti cellulari e fagocitando materiale estraneo, inclusi i batteri.
Alcuni dei tipi principali di tessuto connettivo sono: (1)
tessuto connettivo lasso e denso; (2) tessuto connettivo elastico; (3) tessuto connettivo reticolare; (4) tessuto adiposo;
(5) cartilagine; (6) osso; (7) sangue, linfa e tessuti che producono le cellule del sangue. Questi tessuti variano enormemente nei dettagli strutturali e nelle funzioni che svolgono
(TABELLA 39-2).
Struttura e funzioni degli animali: un’introduzione
819
TABELLA39-2
Fibre collagene
Tessuti connettivi
Nuclei di fibroblasti
Ed Reschke/Getty Images
Fibre elastiche
Tessuto connettivo lasso
Principali localizzazioni
Ovunque l’esigenza di sostegno debba essere combinata con l’elasticità, come nel tessuto sottocutaneo (lo strato di tessuto sotto il
derma della cute)
Funzioni
Sostegno; riserva di fluidi e sali
Descrizione e commenti
Le fibre prodotte dai fibroblasti si immergono nella matrice semifluida; sono presenti altri tipi cellulari, come i macrofagi
50 mm
Immagine al MO di tessuto connettivo lasso.
Tessuto connettivo denso
Nucleo di
fibroblasto
Ed Reschke/Getty Images
Fibre
collagene
Principali localizzazioni
Tendini; molti legamenti; derma della pelle
Funzioni
Sostegno; trasmissione di forze meccaniche
Descrizione e commenti
Le fibre collagene possono essere disposte in modo regolare o
irregolare
25 mm
Immagine al MO di tessuto connettivo denso.
Fibre elastiche
Tessuto connettivo elastico
Principali localizzazioni
Tutte le strutture che sono in grado di espandersi e di ritornare alla
loro dimensione originale, come il tessuto dei polmoni e le grandi
arterie
Ed Reschke/Getty Images
Funzione
Conferisce elasticità
Descrizione e commenti
Fibre elastiche ramificate interposte tra i fibroblasti
50 mm
Immagine al MO di tessuto connettivo elastico.
Fibre reticolari
Tessuto connettivo reticolare
Principali localizzazioni
Struttura interna di fegato, linfonodi e milza
Funzione
Sostegno
Ed Reschke/Getty Images
Descrizione e commenti
Consiste di fibre reticolari intrecciate
50 mm
Immagine al MO di tessuto connettivo reticolare.
Continua
820
CAPITOLO 39
TABELLA39-2
Tessuti connettivi (continua)
Cellule riempite
con depositi
di grasso
Nuclei
Tessuto adiposo
Principali localizzazioni
Strato sottocutaneo; cuscinetti attorno ad alcuni organi interni
Funzioni
Riserva di grasso; isolamento; sostegno di organi come le ghiandole
mammarie e i reni
Descrizione e commenti
Gli adipociti sono inizialmente a forma stellata; le goccioline di
grasso che si accumulano all’interno determinano la tipica forma
rotondeggiante
50 mm
Immagine al MO di tessuto adiposo.
Condrociti
Lacuna
Sostanza
intercellulare
Cartilagine
Principali localizzazioni
Scheletro di sostegno nello squalo e nella razza; parte terminale delle
ossa nei mammiferi e in alcuni vertebrati; anelli di supporto nella
parete della trachea; estremità del naso; orecchio esterno
Ed Reschke/Getty Images
Funzione
Sostegno flessibile
Descrizione e commenti
Le cellule (condrociti) sono separate l’una dall’altra da una sostanza
intercellulare; le cellule sono alloggiate in lacune
50 mm
Immagine al MO di cartilagine.
Lacune
Canale di Havers
Matrice
Osso
Principali localizzazioni
Costituisce la struttura scheletrica nella maggior parte dei vertebrati
Funzioni
Sostegno e protezione di organi interni; riserva di calcio; attacco dei
muscoli scheletrici
Descrizione e commenti
Le cellule (osteociti) alloggiano in lacune; nell’osso compatto le
lacune sono immerse in lamelle, cerchi concentrici di matrice che
circondano i canali di Havers
50 mm
Immagine al MO di osso.
Globuli rossi
Globuli bianchi
Sangue
Principali localizzazioni
All’interno del cuore e dei vasi sanguigni del sistema circolatorio
Ed Reschke/Getty Images
Funzioni
Trasporto di ossigeno, materiali nutritivi, rifiuti e altri materiali; i
globuli bianchi rappresentano un’importante linea di difesa contro i
microrganismi patogeni
Descrizione e commenti
Consiste di globuli rossi, globuli bianchi e piastrine dispersi in una
sostanza intercellulare (plasma)
25 mm
Immagine al MO di sangue.
Struttura e funzioni degli animali: un’introduzione
821
Il tessuto connettivo lasso è il tessuto connettivo maggiormente diffuso nel corpo dei vertebrati. Insieme al tessuto adiposo, forma lo strato sottocutaneo (sotto la cute) che collega la
pelle ai muscoli e alle altre strutture sottostanti. I nervi, i vasi
sanguigni e i muscoli sono avvolti da questo tessuto. Esso funge
da riempimento tra le varie parti del corpo e serve da riserva di
fluidi e sali. Il tessuto connettivo lasso consiste di fibre orientate
in tutte le direzioni all’interno di una matrice semifluida. La sua
flessibilità permette alle strutture a esso connesse di muoversi.
Il tessuto connettivo denso, che si trova nel derma (strato
più profondo) della cute, è molto resistente, benché meno flessibile rispetto a quello lasso. Le fibre collagene sono predominanti. I tendini, le corde che connettono i muscoli alle ossa, e
i legamenti, i cavi che connettono le ossa tra loro, sono costituiti da tessuto connettivo denso in cui i fasci di collagene sono disposti secondo schemi definiti.
Il tessuto connettivo elastico consiste soprattutto di fasci di
fibre elastiche parallele. Questo tipo di tessuto si trova in quelle strutture che sono in grado di espandersi e di tornare successivamente alla loro originale dimensione, come il tessuto dei
polmoni e le pareti delle grandi arterie.
Il tessuto connettivo reticolare è composto soprattutto da
fibre intrecciate. Esso forma la struttura di supporto di alcuni
organi, come il fegato, la milza e i linfonodi.
Il tessuto adiposo è ricco di adipociti che immagazzinano grasso e lo rilasciano quando occorre combustibile per la
respirazione cellulare. Esso si trova nello strato sottocutaneo e
nel tessuto che protegge gli organi interni.
Il supporto scheletrico di un vertebrato è composto da cartilagine, o da cartilagine e osso. Si ricordi che la cartilagine
rappresenta il supporto scheletrico durante lo stadio embrionale di tutti i vertebrati, ma viene largamente sostituita dall’osso durante lo sviluppo, anche se permane in alcune strutture di
supporto. Nell’uomo, la cartilagine forma la struttura di supporto dell’orecchio esterno, gli anelli di supporto delle pareti delle vie respiratorie, l’estremità del naso, le estremità di alcune ossa e i dischi che fungono da cuscinetto tra le vertebre.
La cartilagine è solida ed elastica. Le sue cellule, dette condrociti, secernono attorno a se stesse una matrice dura e gommosa. Esse secernono anche fibre collagene, che si inglobano
nella matrice, rinforzandola. I condrociti trovano alloggio, singolarmente o a gruppi di due o quattro, in piccole cavità della
matrice chiamate lacune e sopravvivono utilizzando il materiale
nutritivo e l’ossigeno che diffondono attraverso la matrice stessa.
Il tessuto cartilagineo è privo di nervi, vasi linfatici e sanguigni.
L’osso, il tessuto principale dello scheletro dei vertebrati, è
simile alla cartilagine, essendo costituito essenzialmente da matrice. Le cellule ossee, chiamate osteociti, sono contenute all’interno delle lacune. Gli osteociti secernono la matrice e provvedono al suo mantenimento (FIG. 39-2). A differenza della cartilagine, tuttavia, l’osso è fortemente vascolarizzato, con un rifornimento cospicuo di sangue. Gli osteociti comunicano tra loro e
con i capillari mediante sottilissimi canali (canalicoli) che contengono i lunghi prolungamenti citoplasmatici degli osteociti.
Un tipico osso lungo ha uno strato esterno di osso compatto, che circonda uno strato interno di osso spugnoso. L’osso compatto è costituito da unità di forma affusolata, chiamate osteoni. All’interno di ogni osteone, gli osteociti sono disposti in strati concentrici, chiamati lamelle, formati dalla matrice. Le lamelle circondano il microscopico canale centrale,
chiamato canale di Havers, in cui passano capillari e nervi.
822
CAPITOLO 39
Le ossa sono straordinariamente leggere e forti. I sali di calcio dell’osso rendono la matrice molto forte e il collagene impedisce che la matrice divenga eccessivamente fragile. La maggior parte delle ossa ha una cavità midollare centrale, che contiene un tessuto spugnoso, chiamato midollo. Il midollo giallo è
costituito principalmente da tessuto adiposo, quello rosso consiste di tessuto connettivo in cui vengono prodotte le cellule del
sangue. L’osso è discusso in maggior dettaglio nel Capitolo 40.
Il sangue e la linfa sono tessuti circolanti che aiutano le
altre parti del corpo a comunicare e a interagire. Come tutti i
tessuti connettivi, consistono di cellule specializzate disperse in
una sostanza intercellulare. Nei mammiferi, il sangue è composto da globuli rossi, globuli bianchi e piastrine, tutti sospesi
nel plasma, che rappresenta la componente liquida e acellulare
del sangue. Nell’uomo e in altri vertebrati, i globuli rossi (eritrociti) contengono un pigmento respiratorio rosso, l’emoglobina,
che trasporta ossigeno. I globuli bianchi rappresentano un’importante linea di difesa contro i microrganismi patogeni (vedi
Capitolo 45). Le piastrine sono piccoli frammenti derivanti da
grandi cellule del midollo osseo. Nei vertebrati complessi, svolgono un ruolo importante nella coagulazione del sangue. Il plasma è costituito da acqua, proteine, sali e una grande varietà di
sostanze solubili, come gli ormoni, che esso trasporta da una
parte all’altra del corpo. Il sangue è discusso nel Capitolo 44.
Il tessuto muscolare è specializzato
nella contrazione
I movimenti della maggior parte degli animali sono la conseguenza della contrazione delle lunghe cellule cilindriche o affusolate del tessuto muscolare. Le cellule muscolari vengono
definite fibre muscolari a causa della loro lunghezza. Ognuna
di esse contiene numerose unità sottili, longitudinali e parallele, chiamate miofibrille. Due proteine, miosina e actina, sono
le principali componenti delle miofibrille e svolgono un ruolo
importante nella contrazione delle fibre muscolari.
Diversi invertebrati hanno tessuto muscolare scheletrico
e liscio. I vertebrati hanno tre tipi di tessuto muscolare: liscio,
scheletrico e cardiaco (TABELLA 39-3 a pagina 824). Il muscolo
scheletrico forma le grandi masse muscolari attaccate alle ossa. Le fibre del muscolo scheletrico sono molto lunghe. Ogni
fibra muscolare scheletrica contiene parecchi nuclei. I nuclei
hanno anche una posizione inusuale: sono localizzati immediatamente sotto la membrana plasmatica, lasciando libera la
parte centrale della cellula per le unità contrattili, le miofibrille. Questa disposizione sembra incrementare l’efficienza della contrazione. Con la contrazione dei muscoli scheletrici, si
verifica il movimento delle varie parti del corpo. Mentre le fibre muscolari scheletriche sono generalmente sotto il controllo della volontà, le fibre muscolari cardiache e lisce non si contraggono normalmente in modo volontario.
Il microscopio ottico mette in evidenza che sia le fibre cardiache che quelle scheletriche hanno bande trasversali alternate, chiare e scure, o striature, che cambiano le loro dimensioni
relative durante la contrazione. Le fibre muscolari striate possono contrarsi rapidamente, ma non possono rimanere contratte a lungo. Esse devono rilasciarsi e riposarsi momentaneamente prima di contrarsi ancora. (La contrazione muscolare è
discussa nel Capitolo 40).
Il muscolo cardiaco è il principale tessuto del cuore.
Quando il muscolo cardiaco si contrae, il cuore pompa sangue.
(a) Lo scheletro umano
consiste principalmente
di osso.
(b) Un osso è stato tagliato
in modo da mettere in evidenza
la sua struttura interna.
Osso
spugnoso
Lacuna
Osso
compatto
Prolungamenti
citoplasmatici
Canale di
Havers
Vaso
sanguigno
(c) Vasi sanguigni
e nervi decorrono
all’interno del canale
di Havers in ogni
osteone dell’osso
compatto.
Osteone
Matrice
Prolungamenti
citoplasmatici
Osteocita
(d) La matrice ossea è rigida e dura. Gli osteociti rimangono
intrappolati all’interno delle lacune, ma comunicano tra loro
mediante prolungamenti citoplasmatici che si estendono
attraverso canalicoli.
Figura 39-2 Osso
Le fibre che lo costituiscono sono unite tra loro alle estremità e
si ramificano e si ricongiungono, formando reticoli complessi.
All’interno di ogni fibra si trovano uno o due nuclei. Una caratteristica del tessuto muscolare cardiaco è la presenza di dischi intercalari, giunzioni specializzate che si trovano nella zona di contatto tra le fibre.
Il muscolo liscio si trova nelle pareti del tratto digerente, dell’utero, dei vasi sanguigni e di molti altri organi interni. La contrazione dei muscoli lisci è necessaria affinché questi organi possano svolgere determinate funzioni. Per esempio, la contrazione della muscolatura liscia della parete del
tratto digerente sposta il cibo lungo il tratto digerente. Quando la muscolatura liscia nelle pareti delle arteriole (piccole
arterie) si contrae, il vaso si restringe e la pressione sanguigna aumenta. Ogni fibra affusolata contiene un singolo nucleo centrale.
Il tessuto nervoso controlla i muscoli
e le ghiandole
Il tessuto nervoso è composto da neuroni, cellule specializzate nella ricezione e nella trasmissione dei segnali, e da cellule gliali, che sostengono e nutrono i neuroni, distruggono
Struttura e funzioni degli animali: un’introduzione
823