
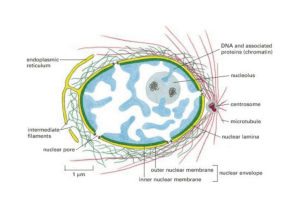
IL NUCLEO
Una cellula somatica umana possiede circa 4000 pori nucleari
Cisterna perinucleare
40-50 nm
La membrana nucleare esterna è continua con le
membrane del reticolo endoplasmatico per cui lo spazio tra le 2
membrane è direttamente collegato al lume del reticolo.
Anche funzionalmente la membrana esterna è simile a quelle del
reticolo endoplasmatico.
La membrana interna contiene invece proteine che legano
la lamina nucleare.
Molecole fino a 20.000 Da e senza segnali di
localizzazione
nucleare possono passare.
Dimensioni maggiori
richiedono SLN e importine.
Il poro nucleare
Canale attraverso cui piccole
molecole polari, ioni e macromolecole
possono passare dal nucleo al
citoplasma e viceversa.
Il diametro di queste strutture è di
circa 120 nm, la massa molecolare
attorno ai 125 milioni di dalton.
Piccole molecole con peso molecolare inferiore a 20 kDa
liberamente attraversano il poro in entrambe le direzioni.
La maggior parte delle proteine e degli RNA deve attraversare il
complesso del poro con un processo attivo, grazie al quali gli
RNA e le proteine vengono riconosciuti e trasportati
selettivamente
Il complesso del poro è una struttura dinamica
costituita da circa 30 proteine diverse chiamate nucleoporine disposte
in gran parte in simmetria ottagonale
Il passaggio attraverso il poro nucleare di molecole e aggregati
macromolecolari molto più grandi come le subunità ribosomali
dimostra che ci sono interazioni specifiche con le nucleoporine
in grado di modificare la sua permeabilità.
Il complesso del poro contiene circa 30 componenti proteici diversi e un PM di
120 MDa.
Il movimento attraverso il poro è regolato da una piccola proteina associata a
GTP chiamata RAN
La Làmina nucleare, è fatta di proteine filamentose appartenenti alla famiglia
dei filamenti intermedi Le lamine A B e C hanno un ruolo strutturale e
regolativo.
Le lamine di tipo A non sono espresse nelle cellule staminali ma solo nei
tessuti differenziati, ciò suggerisce un suo ruolo a stabilizzare l’espressione
genica tessuto-specifica.
Mutazioni del gene della lamina A sono responsabili della progeria
Mutazioni sui geni che codificano per le lamine (LMNA) sono stati associati a
diverse malattie genetiche come la progeria di Hutchinson-Gilford e la distrofia
di Emery e Dreifuss
MATRICE NUCLEARE
E’ visibile al ME come una trama fibro-granulare
diffusa fatta di tre principali componenti:
-La lamina nucleare
-Un reticolo fibro-granulare
-Residui nucleolari
Tra le proteine identificate ci sono
•le lamine,
•le nucleoplasmine (chaperones molecolari per la
formazione di nucleosomi,
ribosomi e la regolazione della trascrizione),
•actina e vimentina.
Alcune proteine di questa matrice si legano
al DNA mediante interazioni specifiche.
I cromosomi non sono distribuiti in modo casuale ma occupano
porzioni discrete definite territori cromosomici
Questo dimostra che al di là
dell’apparente disordine,
la disposizione del materiale
genetico nel nucleo è accuratamente
regolata.
Eterocromatina: più addensata,
trascrizionalmente inattiva,
a duplicazione tardiva. Corrisponde alle
zone centromeriche e telomeriche dei
cromosomi.
Sequenze ripetute e non conservate
Condensata e silente in interfase
Eucromatina: è costituita da una quota
minore, meno condensata e
trascrizionalmente attiva
e una gran parte a condensazione intermedia
e inattiva.
L’eterocromatina
può essere ulteriormente distinta in costitutiva e facoltativa.
L’eterocromatina costitutiva è costituita da regioni di DNA
altamente ripetitivo, costanti in tutte le cellule dell’organismo e
nel cromosoma si concentra principalmente a livello del
centromero e dei telomeri.
L’eterocromatina facoltativa può diventare condensata e
temporaneamente inattiva, inoltre può essere inattivata solo in
determinati tessuti oppure in determinati stadi dello sviluppo .
La cromatina è costituita da DNA e istoni
Gli istoni sono proteine basiche
ricche di lisina e anginina ed
altamente conservate.
Proteine non istoniche sono
associate alla cromatina
e ne regolano le funzioni
(enzimi della duplicazione del DNA,
della trascrizione e i fattori
di trascrizione)
La sintesi degli istoni necessari a formare la cromatina, viaggia insieme
alla duplicazione del DNA
Negli spermatozoi al posto degli istoni troviamo le protamine che consentono
una configurazione più addensata del DNA
Forma nativa
Fibra elementare
La sintesi degli istoni avviene in fase S
insieme alla sintesi del DNA
sotto controllo dei Corpi di Cajal
La formazione di nucleosomi
consente al DNA di accorciarsi
di 1/3
IL NUCLEOLO
È la fabbrica dei ribosomi
Morte cellulare programmata o apoptosi
A differenza della necrosi è un processo attivato
dalla cellula stessa
•Il processo può essere attivato da altre cellule, per es dai linfociti T citotossici
o da cause interne alla cellule come l’ipossia o le radiazioni, e da uno stress acuto
•Richede consumo di energia
•Uno degli eventi chiave è la fuoriuscita del citocromo c dal mitocondrio
che porta all’attivazione delle caspasi responsabili della degradazione
e morte della cellula.
Il nucleo si frammenta in modo caratteristico nel corso dell’apoptosi
Anche il DNA si frammenta in segmenti definiti di lunghezza.
In tutti i casi si attivano processi proteolitici mediati da proteasi chiamate caspasi
L’apoptosi serve nei processi morfogenetici e per eliminare cellule in eccesso o
dannose.
E’ implicata nella crescita tumorale: mutazioni geniche della proteina p53
consentono alle cellule tumorali di sfuggire all’apoptosi.
CORPI APOPTOTICI
Ciclo cellulare: 12-24 ore nelle cellule
attivamente proliferanti
Fase G1: 2-8 ore. Intensa sintesi di RNA e
proteine e accrescimento della
massa cellulare e attivazione delle vie
metaboliche per la successiva
sintesi di DNA.
Fase S: 7-10 ore. La cellula proliferante
duplica il proprio DNA.
Fase G2: 2-5 ore. Si svolgano i processi
metabolici richiesti per
l’esecuzione della divisione cellulare.
Fase M: 2-5 ore. Profase, metafase,
anafase, telofase, citodieresi.
•Degradazione
involucro
nucleare
+ciclina mitotica
= MPF attivo





![mutazioni genetiche [al DNA] effetti evolutivi [fetali] effetti tardivi](http://s1.studylibit.com/store/data/004205334_1-d8ada56ee9f5184276979f04a9a248a9-300x300.png)

![ESTRAZIONE DNA DI BANANA [modalità compatibilità]](http://s1.studylibit.com/store/data/004790261_1-44f24ac2746d75210371d06017fe0828-300x300.png)


