Da
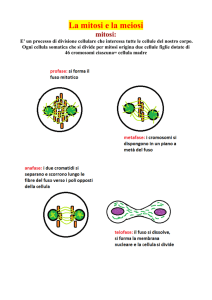
Generalità sulla mitosi
La mitosi è un processo, nell'ambito del ciclo cellulare, in cui la cellula deve suddividere il
proprio genoma in maniera tale che le cellule figlie abbiano, al termine, un ugual numero di
cromosomi. Ricordiamo che la cellula ha già raddoppiato il proprio DNA nella fase S del ciclo
cellulare. Nella prima fase della mitosi questo DNA si organizza sotto forma di cromosomi,
ciascuno formato da due cromatidi, bastoncelli che sono copie identiche l'uno dell'altro.
Successivamente i cromatidi si separeranno e verranno destinati ai nuclei delle cellule figlie.
É come se il nostro genoma fosse un romanzo, un libro composto da più capitoli, i cromosomi. La
cellula prima fa una copia completa di ogni capitolo e successivamente sudpide queste copie in
modo tale che ogni cellula abbia il suo "libro" completo, con tutti i capitoli di cui è composto,
necessari alla comprensione del romanzo.
Dopo l'ultima fase della mitosi, quasi sempre il citoplasma viene piso e si formeranno due cellule
indipendenti, ciascuna con il genoma identico e completo.
La mitosi nel dettaglio
La mitosi è un processo mediante il quale la cellula sudpide i propri cromosomi tra le cellule figlie,
in modo tale che ciascuna di esse abbia un corredo genetico completo.
Per
facilità
di
studio
viene
pisa
in quattro
fasi: profase, metafase, anafase e telofase.
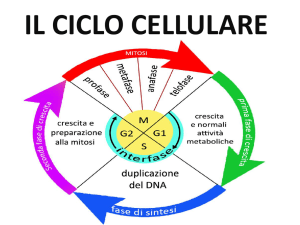
Ricordiamo che alla fine dell'interfase nel nucleo il genoma è già stato duplicato. Nella fase S
infatti, i filamenti, ancora sotto forma di cromatina, vengono trascritti in maniera completa.
Nella profase, che occupa circa il 60% della mitosi, i cromosomi si spiralizzano; i filamenti di
DNA, invisibili al microscopio ottico, si avvolgono attorno a particolari proteine associate ad esso,
dette proteine istoniche, e formano dei grossi bastoncelli (cromatidi) che rappresentano le due copie
del cromosoma. É a questo punto che i cromosomi risultano visibili al microscopio ottico; essi
hanno ancora un aspetto filamentoso.
Il cromosoma: è costituito da due cromatidi, copie dello stesso tratto di DNA
Mitosi, infatti, deriva dal greco mìtos, che significa filo. I cromatidi sono uniti in un'area
detta centromero. Il centromero è di vitale importanza per la mitosi; senza di esso il processo non
si completerebbe e la cellula morirebbe. Attorno al centromero è presente una struttura specializzata
detta cinetocore o cinetocoro, dalla quale si dipartono delle fibre proteiche, dette fibre del
cinetocore.
Queste fibre sono necessarie perché permetteranno ai cromosomi di ancorarsi ad una struttura di
fondamentale importanza per la mitosi: il fuso mitotico. Nelle cellule animali ed in alcune cellule
vegetali (tranne le Angiosperme, cioè le piante con fiore) vanno formandosi, appena al di fuori della
membrana che va via via dissolvendosi, due coppie di centrioli. Questi particolari corpuscoli sono
presenti a coppie: hanno una struttura identica ai corpi basali, fondamentali nella costruzione dei
flagelli e delle ciglia. Sono anch'essi costituiti da fascetti di microtubuli, nove triplette disposte a
raggio. Talvolta può essere presente una struttura proteica che si diparte da loro, disposta a raggio,
detta aster. I centrioli, appena vengono sintetizzati, si dispongono ad angolo retto: da essi prendono
forma le fibre del fuso. Mano a mano che queste fibre si formano, spingono i centrioli verso i poli
della cellula. La cellula, nel frattempo, cerca di allineare i cromosomi, tirandoli o spingendoli
tramite le fibre del fuso. Quest'ultimo conseguirà la sua tipica forma affusolata, simile ad una palla
da rugby, nella fase successiva.
Schema della profase: il nucleo si sta dissolvendo, i cromosomi sono ormai visibili; i centrioli si
stanno dirigendo verso i poli della cellula e stanno sintetizzando il fuso mitotico. (Per semplicità
sono state disegnate solo due coppie di cromosomi omologhi, con colori analoghi).
Nella metafase i cromosomi sono disposti tutti su di uno stesso piano immaginario, detto piano
equatoriale cellulare, posto al centro del fuso mitotico: i cromatidi sono orientati verso i poli della
cellula.
Schema della metafase: i cromosomi si sono disposti sullo stesso piano
//
Nella fase successiva, detta anafase, i cromatidi di ogni coppia si separano all'altezza del
centromero e migrano verso i poli cellulari lungo le fibre del fuso. Ogni cromatidio costituirà un
cromosoma indipendente. Anche i poli della cellula tendono ad allontanarsi, mentre la cellula stessa
tenderà ad allargarsi.
Schema dell'anafase: i due cromatidi di ciascun cromosoma si sono separati all'altezza del
centromero e si muovono lungo le fibre del fuso mitotico, verso i centrioli.
La fase finale della mitosi, detta telofase, vede la formazione di due nuovi involucri nucleari,
all'interno dei quali sono racchiusi i due gruppi di cromosomi che andranno verso la
despiralizzazione per formare la cromatina. Vanno costituendosi anche i nucleoli. Il fuso mitotico
tende a dissolversi: le molecole di cui è composto serviranno per assemblare il nuovo citoscheletro
delle cellule figlie. Al termine di questa fase ogni cellula possiede lo stesso genoma, cioè un numero
identico di cromosomi, ciascuno dei quali è costituito da un solo cromatidio.
Schema della telofase: si riformano i nuclei, che ora contengono il genoma completo della cellula.
Il fuso si sta dissolvendo e si riforma il citoscheletro. I cromosomi sono costituiti da un solo
cromatidio.
Per la cellula è arrivato il momento di pidersi completamente: la citodieresi.
Nelle cellule animali si forma un restringimento, all'incirca a metà strada tra i due nuclei, formato
da un anello di filamenti proteici. Questa strozzatura si accentua sempre di più, fino a separare di
fatto il citoplasma, con la conseguente formazione di nuova membrana plasmatica. La citodieresi
nelle cellule animali comincia quindi dalla membrana plasmatica e precede verso il centro.
Nelle cellule vegetali questo tipo di meccanismo viene impedito dalla presenza di una parete
cellulare rigida. Si forma una piastra cellulare lungo la linea mediana della cellula. Essa deriva
dalla fusione di vescicole contenenti polisaccaridi prodotti dall'apparato di Golgi. Mano a mano
queste vescicole si fanno più numerose ed ai loro bordi si forma prima una membrana cellulare che
va estendendosi verso i lati della cellula, fino a fondersi con la membrana preesistente;
successivamente ogni cellula figlia sintetizzerà la propria parete cellulare. La citodieresi nelle
cellule vegetali comincia quindi dal centro e procede verso la periferia.
APPROFONDIMENTO SUL CROMOSOMA:
cromosoma Nome
dato da W. Waldeyer nel 1888 ai piccoli corpi intensamente colorabili, in
genere di forma bastoncellare, visibili nel nucleo della cellula durante la mitosi. Secondo la teoria
cromosomica, dimostrata da T.H. Morgan con ricerche su Drosophila melanogaster, i c. sono le
strutture essenziali dell’eredità.
1. Caratteri generali
La struttura base è costituita da un filamento a doppia elica di DNA avvolto intorno agli istoni
(proteine basiche). Nella sequenza di basi del DNA è scritta tutta l’informazione genetica di una
cellula per la formazione di proteine; una sequenza di basi, che codifica una proteina o meglio una
catena polipetidica, prende il nome di gene. Quando il DNA si duplica, durante l’interfase del ciclo
cellulare, l’informazione genetica viene riprodotta fedelmente ed è trasmessa alle cellule figlie. Al
momento della mitosi il DNA è quindi già duplicato e il c. appare diviso in due elementi,
icromatidi, uniti a livello del centromero (fig. 1). Quando si completa la divisione cellulare ognuna
delle cellule figlie riceve uno dei due cromatidi. I cromatidi sono costituiti da un’unica molecola di
DNA a doppia elica molto spiralizzata. La spiralizzazione di questa molecola si realizza mediante
stretti avvolgimenti intorno alle proteine istoniche in strutture fondamentali chiamate nucleosomi.
Durante la metafase mitotica, i c. appaiono al microscopio ottico ben divisi nei due cromatidi ed è
possibile studiare e individuare ogni singolo c. in base ad alcuni parametri: a) lunghezza relativa dei
c.; b) presenza di una costrizione, situata sempre nella stessa posizione (costrizione primaria o
centromero); a seconda della posizione del centromero, che è considerato il centro cinetico del c. e
corrisponde al punto in cui il c. si attacca alle fibre del fuso durante l’anafase, si distinguono: c.
metacentrici o submetacentrici, con centromero in posizione mediana e submediana;c. acrocentrici,
con centromero subterminale; c. telocentrici, in cui il centromero è terminale (fig. 2). Il rapporto fra
le lunghezze dei due bracci di un c., misurate dal centromero, è detto indice centromerico; nel
metacentrico l’indice centromerico è pari a 1; c) presenza di costrizioni secondarie (studi citologici
e biochimici hanno rivelato che la costrizione secondaria dei c. è il sito dove sono localizzati i geni
che codificano per l’RNA ribosomale, cioè l’RNA che costituisce i ribosomi; questo RNA, appena
prodotto, si addensa a formare, insieme a una componente proteica, i nucleoli che rimangono vicini
alla costrizione secondaria e che per tale ragione viene chiamata regione dell’organizzatore
nucleolare); d) presenza di satelliti, formazioni sferiche terminali unite al c. con un sottile filamento.
Ulteriori dati sull’individuazione dei c. si ottengono con particolari tecniche di colorazione
(➔ bandeggio).
Il numero e la forma dei c. sono costanti per ogni specie di animali e di piante; negli organismi
diploidi il corredo cromosomico completo diploide (indicato con il simbolo 2 n) è rappresentato da
due serie di c. identici tra loro due a due, detti omologhi; per es. l’uomo ha 2 n = 46 c. (23 coppie);
il moscerino Drosophila melanogaster 2 n = 8 c. (4 coppie); il granturco Zea mais 2 n = 20 c. (10
coppie); il topo 2 n = 40 c. (20 coppie) e così via (per le eccezioni a questa regola ➔ poliploidia).
L’informazione genetica dei c. omologhi deriva per metà dal gamete maschile e per metà dal
gamete femminile. Durante il processo di formazione dei gameti (negli organismi a riproduzione
sessuale) il numero diploide 2 n dei c. viene, attraverso il processo della meiosi (o gametogenesi),
dimezzato a n cromosomi (numero aploide). All’atto della fecondazione tra i due gameti si
ricostituisce nello zigote il numero diploide.
2. Tipi di cromosomi
C. sessuali In molte specie di animali e in alcuni vegetali a sessi separati si osserva una coppia di c.
che differiscono nel maschio e nella femmina, e che sono appunto chiamati c. sessuali
(o eterocromosomi), mentre gli altri c. del corredo prendono il nome di autosomi. Si possono avere
diversi casi: la femmina ha due cromosomi uguali (XX) e il maschio due diversi (XY) oppure uno
solo (XO); in questo caso il sesso femminile è omozigote o omogametico, il maschile eterozigote o
digametico. Alcuni geni sono localizzati sui c. sessuali: nell’uomo appartengono a questa categoria,
detta dei caratteri ‘legati al sesso’, il daltonismo, l’emofilia e altri geni. C. giganti Detti
anche salivari, sono presenti nel nucleo delle grosse cellule delle ghiandole salivari delle larve di
alcuni Ditteri (in particolare Chironomus e Drosophila), si caratterizzano per: a) l’appaiamento dei
c. omologhi (sinapsi somatica), per cui il numero di c. risulta aploide; b) la notevole lunghezza,
dovuta al fatto che essi si trovano in uno stato di forte despiralizzazione; c) l’ammasso
eterocromatico ( cromocentro) nel quale sono riuniti tutti i centromeri e buona parte del c. Y; d) la
presenza di un’alternanza di bande chiare e scure che sono state interpretate come formate dalla
giustapposizione dei cromomeri allineati sui numerosi cromatidi appaiati (e per questa ragione i c.
salivari vengono detti anche politenici); si ritiene infatti che i c. salivari originino da un processo di
endomitosi, cioè da una serie di duplicazioni successive dei c. non seguiti da divisioni cellulari. In
particolari stadi della vita larvale di Drosophila è possibile individuare lungo i bracci di questi c. dei
rigonfiamenti detti anelli di Balbiani (o puffs) al cui interno si verifica una sintesi molto attiva di
RNA. C. a spazzola Nome dato da J. Rückert a particolari c., detti anche piumosi, presenti negli
oociti di alcuni vertebrati (specialmente anfibi), durante lo stadio meiotico del diplotene; vanno
perciò considerati bivalenti. Lungo i cromatidi, molto despiralizzati, si evidenziano i cromomeri, in
corrispondenza dei quali si formano due anse; poiché le anse sono molto numerose, i c. assumono
un aspetto piumoso. È stato dimostrato che lo stadio piumoso corrisponde a un periodo di attiva
sintesi di RNA. C. soprannumerari In numerose specie animali e vegetali alcuni individui
presentano un numero di c. diverso da quello normale e costante della specie a causa dell’esistenza
dei c. soprannumerari eterocromatici (detti anche c. accessori o B-cromosomi), i quali non sono
omologhi a nessun membro del corredo cromosomico della specie; tale fenomeno pertanto non va
confuso con i casi di ploidia (➔ poliploidia).
3. C. dei Procarioti
Nei virus il c. è rappresentato da una singola molecola di DNA o di RNA strettamente arrotolata. Il
DNA può essere a singolo filamento o a doppio filamento, può avere forma ad anello o essere
lineare; in alcuni batteriofagi (fago lambda) il c. ha doppia elica, forma bastoncellare e le estremità
sono ‘adesive’, per cui dopo che il virus è penetrato nella cellula ospite, il c. prende forma di anello.
Nei batteri la regione che contiene il DNA viene detta nucleoide, per analogia con il nucleo degli
Eucarioti. Nel nucleoide il DNA è strettamente attorcigliato, tanto da risultare escluso dalle
particelle citoplasmatiche per quanto non sia circondato da una membrana come nel nucleo degli
Eucarioti. Il c. batterico è stato studiato in Escherichia coli, che ha un unico c. circolare a doppio
filamento di DNA.
4. Malattie cromosomiche
Sindromi associate ad alterazioni della struttura o del numero dei cromosomi. Le più note
nell’uomo sono: la trisomia 13; la trisomia 18; la trisomia 21 (➔ Down, John Langdon Haydon).
CROMATINA:
cromatina Nome dato da W. Flemming (1879) a quella parte della sostanza di cui è costituito il
nucleo cellulare che si colora intensamente con i coloranti basici usati nella tecnica istologica
(ematossilina, blu di metilene, safranina, verde di metilene, fucsina basica). Durante la mitosi, la c.,
che si osserva nel nucleo interfasico come zolle intensamente colorate, ma che in realtà ha struttura
filamentosa, si organizza spiralizzandosi e forma i cromosomi. La basofilia della c. è dovuta
agli acidi nucleici che sono tra i suoi costituenti principali. La c. infatti è un complesso
nucleoproteico stabile comprendente DNA e proteine acide e basiche (principalmente istoni); a essa
sono associati alcuni enzimi e molecole di RNA. In particolare, l’unione degli istoni al DNA porta
alla formazione di nucleosomi, unità fondamentali della cromatina. I nucleosomi determinano
il tipico/">tipico aspetto a collana di perle della c. quando essa si osserva al microscopio elettronico,
dopo averla trattata con sostanze che distendono le ripiegature di ordine superiore.
Si possono distinguere due tipi di c., l’ eucromatina, che si colora meno intensamente e rappresenta
zone cromosomiche di despiralizzazione, e l’ eterocromatina, che si colora più intensamente e
rappresenta zone più condensate. I due tipi di c. hanno significato genetico diverso: nello stato
eterocromatico condensato il DNA è inattivo e non viene trascritto in RNA; si può perciò
considerare l’eterocromatina come lo stato inerte e l’eucromatina come quello attivo del DNA
cromosomico.
Alcune regioni dei cromosomi sono condensate in eterocromatina in tutte le cellule e vengono
indicate come eterocromatina costitutiva; altre regioni sono condensate solo in particolari cellule e
costituiscono la cosiddetta eterocromatina facoltativa. Nell’uomo l’ eterocromatina costitutiva si
distingue nei cromosomi mitotici, con colorazioni particolari, come una banda scura attorno al
centromero di ciascun cromosoma; in altri Mammiferi forma delle bande lungo i cromosomi o
occupa un intero braccio del cromosoma. Nell’interfase del ciclo cellulare, zone di eterocromatina
costitutiva si riuniscono a formare i cromocentri. Nei Mammiferi il numero e la disposizione di tali
cromocentri sono diversi nei vari tipi di cellule a seconda dello stadio di sviluppo. La maggior parte
delle zone di eterocromatina costitutiva contiene sequenze di DNA ripetute che, per ragioni di
ordine storico, vengono indicate come DNA satellite. Il significato della eterocromatina
facoltativa è certamente più chiaro rispetto alla costitutiva: essa riflette le differenti attività
genetiche di diversi tipi di cellule. Per es., le cellule embrionali ne possiedono poca, mentre cellule
altamente specializzate molta; questo fa pensare che, mentre la cellula si sviluppa, viene inattivato
in modo permanente un numero sempre più elevato di geni. La maggior parte delle conoscenze
sull’eterocromatina facoltativa deriva dalle ricerche sulla inattivazione di uno dei due cromosomi X
nelle cellule femminili dei Mammiferi, il cosiddetto ‘corpo di Barr’.
Gli istoni sono delle proteine basiche che presentano elevate quantità di aminoacidi basici come Lisina (Lys)
o Arginina (Arg). La carica fortemente positiva le rende capaci di legarsi fortemente al DNA e di neutralizzare
la sua carica negativa dovuta al gruppo fosfato . Vi sono cinque tipi di Istoni: H1, H2A, H2B, H3, H4. Il DNA
si avvolge attorno ad una struttura proteica della Nucleosoma, costituito da un ottamero di proteine, dove il
DNA compie un giro e tre quarti. L'ottamero è costituito da due copie di ciascun istone H2A, H2B, H3, H4 e
costituisce il cuore dell'unità di impacchettamento più elementare del DNA. Il doppio giro di DNA è poi
sigillato dall'istone H1. Ogni nucleosoma avvolge attorno a sé una lunghezza di DNA pari a circa 200
nucleotidi. Solo H1 consente ai nucleosomi di avvolgersi a spirale a costituire una struttura denominata fibra
da 30 nm dal suo spessore.