26/05/2013
Citoscheletro
Filamenti Intermedi
Coulombe PA, Wong P. Cytoplasmic intermediate filaments revealed as dynamic and multipurpose scaffolds. Nat Cell Biol. 6: 699‐706, 2004.
“I filamenti intermedi sono polimeri
forti ma flessibili che forniscono
sostegno meccanico alle cellule che
vanno dai batteri ai tessuti umani”
(Pollard et al., 2008)
I filamenti intermedi sono polimeri citoscheletrici codificati da una grande famiglia di geni espressi in modo differenziale che forniscono un sostegno strutturale cruciale al citoplasma e al nucleo degli eucarioti superiori. Perturbazioni della loro funzione spiegano diverse patologie determinate geneticamente in cui le cellule fragilizzate non sono in grado di reggere stress meccanici e non‐meccanici. Studi recenti chiariscono come il sostegno strutturale sia modulato per affrontare le necessità mutevoli delle cellule e rivelano un nuovo ruolo tramite il quale i filamenti intermedi influenzano la crescita e la morte cellulare mediante interazioni dinamiche con proteine non‐strutturali.
Adattato da Newsletter della Biomeda, Summer 1996, Volume 1, Issue 1
Herrmann H, Strelkov SV, Burkhard P, Aebi U. Intermediate filaments: primary determinants of cell
architecture and plasticity. J Clin Invest. 119:1772-1783, 2009.
I filamenti intermedi (IFs) sono i principali costituenti del citoscheletro e
dell’interfaccia col nucleo delle cellule animali.
Svolgono ruoli di importanza fondamentale nell’organizzazione degli
elementi strutturali.
A seconda del tipo cellulare, proteine morfologicamente simili ma
distinte biochimicamente formano filamenti ad elevata viscoelasticità
che svolgono molteplici funzioni nanomeccaniche.
Oltre al loro ruolo primario nella plasticità cellulare e alla loro funzione
già ben stabilita di ammortizzatori per gli stress cellulari, alterazioni
geniche recentemente identificate hanno elucidato che le alterazioni
strutturali degli IFs possono influenzare il loro coinvolgimento sia
nel signaling che nelle reti di geni regolatori.
In questo articolo evidenziamo le proprietà strutturali e funzionali
basiche degli IFs e ricaviamo un concetto su come le mutazioni possano
influenzare l’archittetura cellulare e perciò la costruzione e la fisiologia dei
tessuti.
Filamenti Intermedi - BUSB2011
1
26/05/2013
FILAMENTI INTERMEDI (1)
FILAMENTI INTERMEDI (2)
Hanno diverse proprietà carattereristiche che li distinguono dai microfilamenti e dai microtubuli:
Sono biochimicamente molto più eterogenei:
Esistono subunità dei filamenti intermedi diverse , ma correlate evolutivamente, che sono spesso
espresse in modo tessuto‐dipendente.
Hanno grande forze tensile; es:
Peli e unghie: consistono principalmente di filamenti intermedi delle cellule morte
NON hanno una polarità intrinseca come i microfilamenti e i
microtubuli, e le loro unità costituenti NON si legano ad un nucleotide.
Dato che non hanno polarità intrinseca non si conoscono
proteine motori che li usino come rotaie
Nonostante siano dinamici in termini di scambio di subunità, sono molto più stabili dei microfilamenti e i microtubuli dato
cha la velocità di scambio è molto più lenta.
NON si trovano in tutti gli eucarioti:
I funghi e le piante non hanno filamenti intermedi
Gli insetti hanno soltanto una classe, rappresentata da due geni che esprimono le lamine A/C e B (FI che sorreggono
l’involucro nucleare).
Caratteristiche dei Filamenti Intermedi
Stabili, resistenti
8 – 10 nm di diametro
Prominenti nelle cellule che debbono resistere a stress meccanici
Insolubili in soluzioni concentrate di sali e detergenti non ionici
IFs caratteristici e specifici per i iversi tipi di tessuti
Si possono sciogliere con urea (potente denaturante per le proteine)
FI: Struttura (1)
Ogni isoforma delle proteine dei FI ha una sequenza
aminoacidica caratteristica, ma tutti hanno un dominio a bastoncello fra i domini di testa e coda con dimensione
variabile, presenti alle due estremità. Il bastoncello è un “coiled‐coil” parallelo di due α‐eliche, di solito lungo circa 47 nm.
Come gli altri “coiled‐coils”, i domini a bastoncello dei FI hanno una ripetizione di sette AA, in cui il primo e quarto residui forniscono una riga continua di interazioni
idrofobiche lungo la l’interfaccia delle due eliche.
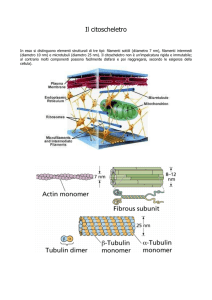
Filamenti intermedi
Struttura
FI: Struttura (2)
Zone di carica positiva e negativa si alternano lungo il
bastocello.
Quando sfalsate in modo corretto, queste zone forniscono
legami elettrostatici complementari per l’assemblaggio
dei filamenti.
Circa 20 residui altamente conservati ad ogni estremità del bastoncello sono essenziali per l’allungamento del filamento
mediante interazioni testa‐coda fra molecole dimeriche. Studi con proteine mutanti suggeriscono che queste parti del bastoncello contribuiscono alle associazioni laterali all’interno
dei filamenti.
Sequenze aminoacidiche indicano la presenza di tre
interruzioni nel “coiled‐coil”
Filamenti Intermedi - BUSB2011
2
26/05/2013
Didascalia della Fig. precedente:
Le proteine dei filament intermedi hanno domini testa e coda di dimensioni variabili che fiancheggiano un dominio centrale a bastoncello.
I domini a bastoncello consistono in un “coiled‐coil” di circa 310 residui e sono lunghi 46,5 nm.
Le lamìne hanno 42 residui addizionali nel bastoncello (λ).
I residui più importanti per l’assemblaggio si trovano all’inizio e alla fine del bastoncello.
I domini terminali differiscono in sequenza e dimensione da 6 a 1200 residui.
ETEROGENEITÀ MOLECOLARE DEI FILAMENTI INTERMEDI - 1
Mentre i filamenti di actina e i microtubuli sono polimeri di un unico tipo di proteina (actina e tubulina, rispettivamente), i filamenti intermedi (FI) sono composti da una gran varietà di proteine che sono espresse da tipi cellulari diversi.
Sono stati identificati più di 65 proteine diverse di FI, che sono state classificate in sei gruppi in base alle somiglianze di sequenze aminoacidiche.
Livelli di organizzazione ed assemblaggio dei filamenti intermedi 2
Filamenti intermedi
Polimerizzazione
Filamenti Intermedi - BUSB2011
Le proteine dei filamenti intermedi (IF) formano omo‐ ed eterodimeri con un dominio altamente conservato ad alfa‐elica e code e teste non elicoidali, che hanno dimensione e sequenza di aminoacidi variabili. Il dominio “core” centrale contiene tre elementi spaziatori non elicoidali. Un tetramero si forma mediante aggregazione sfalsata, antiparallela di due dimeri identici. I tetrameri si aggregano coda contro coda, formando un protofilamento; successivamente copie di protofilamenti si associano lateralemente formano una protofibrilla. L’associazione laterale di quattro protofibrille forma un cilindro con 10 nm di spessore.
3
26/05/2013
Livelli di organizzazione ed assemblaggio
dei filamenti intermedi (1)
http://www.ncbi.nlm.nih.gov/books/NBK26862/figure/A2984/
http://www.ncbi.nlm.nih.gov/books/NBK21560/figure/A5550/
Proprietà meccaniche dei polimeri di actina, tubulina e filamenti
intermedi
Lunghe subunità sfalsate: predominano i contatti laterali.
http://www.ncbi.nlm.nih.gov/books/NBK26862/figure/A2963/
Filamenti Intermedi - BUSB2011
Le reti composte da microtubuli, filamenti di actina o dei filamenti intermedi chiamati
vimentina, tutti alla stessa concentrazione, furono esposti a forze di taglio in un viscosimetro, e il grado di stiramento è stato
misurato. I risultati mostrano che le reti di microtubuli sono facilmente deformabili
ma si rompono (illustrato dall’asterisco
fucsia) e cominciano a fluire senza limite
quando stirati oltre il 150% della loro
lunghezza primitiva. Le reti di filamenti di actina sono molto più rigide, ma anche
esse si rompono facilmente. Viceversa, le reti di filamenti intermedi non solo si
deformano facilmente ma inoltre resistono
a elevati stress e tensioni senza rompersi; perciò essi sono ben adatti a sostenere
l’integrità della cellula. http://www.ncbi.nlm.nih.gov/books/NBK26862/figure/A2985/?report=objectonly
4
26/05/2013
Filamenti intermedi
TIPI
ETEROGENEITÀ MOLECOLARE DEI FILAMENTI INTERMEDI - 2
Adattato da: Cooper GM & Hausman RE: The Cell: A Molecular Approach”, 4th ed, ASM Press, Sinauer Associates
Omary MB. "IF-pathies": a broad spectrum of intermediate filament-associated diseases. J Clin Invest. 2009 Jul;119(7):175662.
L’ampia e
complessa
distribuzione
dei FI nei
tessuti umani
Omary MB. "IF-pathies": a broad
spectrum of intermediate filamentassociated diseases. J Clin Invest.
2009 Jul;119(7):1756-62.
ETEROGENEITÀ MOLECOLARE DEI FILAMENTI INTERMEDI - 3
• I tipi I e II consistono in due gruppi di cheratine, ciascuno consistente di circa 15 proteine diverse, che sono espresse nelle cellule epiteliali.
Ogni tipo di cellula epiteliale sintetizza almeno un tipo di cheratina I (acide) e un tipo di tipo II (basiche, neutre), che copolimerizza per formare filamenti.
Alcuni tipi di cheratine di tipo I e II (dette cheratine dure) sono usate per produrre strutture dure quali i capelli, le unghie e le corna.
Altri tipi di cheratina (cheratine molli) sono abbondanti nel citoplasma delle cellule epiteliali; nei vari tipi di cellule epiteliali sono espresse combinazioni diverse.
Adattato da: Cooper GM & Hausman RE: The Cell: A Molecular Approach”, 4th ed, ASM Press, Sinauer Associates
Filamenti Intermedi - BUSB2011
5
26/05/2013
ETEROGENEITÀ MOLECOLARE DEI FILAMENTI INTERMEDI - 3
TIPI DI FILAMENTI INTERMEDI ‐ 2
NEUROFILAMENTI
• I tipi I e II consistono in due gruppi di cheratine, ciascuno consistente di circa 15 proteine diverse, che sono espresse nelle cellule epiteliali.
Ogni tipo cellulare sintetizza almeno un tipo di cheratina I (acide) e un tipo di tipo II (basiche, neutre), che copolimerizza per formare filamenti.
Alcuni tipi di cheratine di tipo I e II (dette cheratine dure) sono usate per produrre strutture dure quali i capelli, le unghie e le corna.
Altri tipi di cheratina (cheratine molli) sono abbondanti nel citoplasma delle cellule epiteliali; nei vari tipi di cellule epiteliali sono espresse combinazioni diverse.
Queste tre proteine, NF‐L, NF‐M e NF‐H sono sintetizzate nei neuroni.
Non sono distribuite in modo uguale:
i corpi cellulari contengono soprattutto NF‐L e NF‐H.
gli assoni esprimono preferenzialmente NF‐M e NF‐H
Hanno residui carbossilici altamente carichi.
VIMENTINA
Questo filamento intermedi è espresso dai tessuti mesenchimali ed è tipica delle cellule primitive.
Le cellule in coltura ritornano ad esprimere vimentina anche se questo non è il loro filamento intermedio del tessuto maturo, forse come risultato della stimolazione a migrare e a dividersi rapidamente.
Adattato da: Cooper GM & Hausman RE: The Cell: A Molecular Approach”, 4th ed, ASM Press, Sinauer Associates
TIPI DI FILAMENTI INTERMEDI ‐ 3 Adattato da Smith and Wood:
Cell Biology,
Chapman & Hall, London, 1992
BUSB 2011 ‐
Filamenti Intermedi
32
Principali funzioni cellulare dei filamenti intermedi citoplasmatici -1
DESMINA
Questa proteina è espressa dalle cellule muscolari scheletriche,
aumentando negli stadi più tardivi di differenziamento, quando
l’espressione della vimentina cessa.
La desmina e la vimentina sembrano aiutare i microfilamente ad
allinearsi in registro nelle fibre muscolari scheletriche ed entrambe
rimangono come componenti minori nel muscolo scheletrico maturo.
La desmina è presente nella muscolatura liscia ed anche in cellule non
muscolari con attività contrattile, come i periciti, le cellule stellate del
fegato, le cellule mio-epiteliali e i podociti del rene.
E’ anche presente in altre cellule non muscolari come le cellule
endoteliali.
Quindi, l’espressione di desmina non è interamente muscolo-specifica.
Adattato da Smith and Wood: Cell Biology, Chapman & Hall, London, 1992
Principali funzioni cellulari dei filamenti intermedi citoplasmatici -2
Citoarchittetura. Nei motoneuroni, la crescita radiale dei processi assonali richiede la loro
interazione con neurofilamenti (blu chiaro) in modo da trovare la corretta stechiometria fra le subunità “light” (NF‐L), media (NF‐M) e “heavy” (NF‐H). I grandi domini della coda C‐terminale delle
subunità NF‐H (rosso) e NF‐M (arancione) sono iperfosforilati e si proiettano al di fuori della zona
centrale del filamento, così determinando la spaziatura fra I filamenti e il calibro dell’assone. Gli
abbondanti neurofilamenti interagiscono con I microtubuli meno abbondanti (blu scuro) e con I filamenti di actina subcorticali (verde scurto) mediante proteine linker del citoscheletro, quali la plectina e la BPAG1 (giallo). Un ruolo citoarchitteturale è stato inoltre dimostrato per le lamine
nucleari ed altri filamenti intermedi citoplasmatici quali la desmina e le cheratine. Filamenti Intermedi - BUSB2011
Sostegno meccanico: L’epidermide è un buon esempio per illustrare questa funzione che è condivisa dalla
maggior parte dei filamenti intermedi. I filamenti intermedi di tipo cheratina sono abbondanti nei cheratinociti, con un range fra > 10% del contenuto totale di proteine nelle cellule progenitrici basali fino a > 70% nelle cellule di differenziamento più avanzato. I cambiamenti nel colore dei filamenti riflettono l’espressione differenziale e la composizione delle cheratine nelle cellule basali, di differenziamento precoce e di differenziamento tardivo (la freccia indica il differenziamento). Le reti di filamenti di cheratina si estendono attraverso tutto il citoplasma dei
singoli cheratinociti e sono integrate fra cellule mediante collegamento alle giunzioni cellula‐cellula di tipo
desmosomi (punti rossi) e fra le cellule basali e la lamina basale mediante collegamento ad emi‐desmosomi
(punti gialli). Questa organizzazione massimizza il sostegno meccanico fornito dai filamenti di cheratina.
Coulombe PA, Wong P. Cytoplasmic intermediate filaments revealed as dynamic and multipurpose scaffolds. Nat Cell Biol. 2004 Aug;6(8):699-706.
Principali funzioni cellulari dei filamenti intermedi citoplasmatici - 3
Migrazione cellulare: Nel flusso sanguigno i linfociti resistono a stress emodinamici e meccanici a causa, parzialmente della loro rete di filament intermedi di vimentina che è organizzata in una configurazione tipo gabbia alla periferia del citoplasma. In seguito a chemotassi indotta da chemochine, ad esempio in siti di attiva infiammazione, i filamenti di vimentina rapidamente si spostano dalla periferia nucleare all’uropodo della cellula. Ciò è in parte reso possibile mediante fosforilazione sito‐specifica delle subunità di vimentina (illustrata da cambiamento del colore dei fila menti nella cellula a destra) ed è correlata con un ammorbidimento delle proprietà viscoleastiche del citoplasma, presumibilmente per permettere la flessibilità necessaria durante l’estravasione. Gli stessi principi generali sono alla base della capacità delle cellule epitelaili di migrare in un sito di ferita dopo il danno.
6
26/05/2013
Principali funzioni cellulari dei filamenti intermedi citoplasmatici - 4
Modulazione di segnale. I filament intermedi citoplasmatici possono legare e modulare l’attività di proteine di segnalamento, influenzando in questo modo il flusso di segnali extracellulari verso effettori terminali rilevanti all’interno della cellula. Sono illustrati due possibili meccanismi:
Sinistra: interazioni dei FI con recettori alla superficie della cellula quali Fas, modula la loro densità e funzione.
Destra: interazioni regolate fra i FI e una proteina adattatrice quale TRDD (rosa), vicino alla superficie cellulare, limita la disponibilità di questo adattatore ad un recettore legato ad un ligando, che è necessaria per trasmettere un segnale alla cellula. La regolazione di qualsiasi tipo di reazione, indicata qui da una variazione locale del colore del filamento (blu o rosso), può essere mediata da modificazioni post‐traduzionali dinamiche, associazione con altre proteine o differenze locali nella composizione delle subunità dei FI. Entrambi questi meccansimi possono partecipare alla regolazione della risposta delle cellule epitelaili a segnali pro‐apoptotici e altri eventi di segnalamento.
Filamenti intermedi
CHERATINE
CITOCHERATINE
Cheratine
Questo è il primo tipo di filamento intermedio a comparire durante la segmentazione dello zigote ed è già rivelabile allo stadio di 8 cellule. Più tardi, durante l’embriogenesi, la famiglia delle citocheratine è presente nelle cellule epiteliali.
Gli epiteli semplici, monostratificati o le cellule epiteliali in rapida divisione contengono le citocheratine di PM più basso, mentre gli epiteli complessi, con cellule altamente differenziate contengono citocheratine di dimensioni maggiori, con terminali aminico e carbossilico idrofobici. Questi interagiscono con una proteina citoplasmatica strettamente associata, la filaggrina, per formare un complesso insolubile ma molto malleabile che fornisce la barriera protettiva della pelle.
Le citocheratine sono anche associate ai desmosomi, le giunzioni che collegano le cellule epiteliali.
BUSB 2011 ‐ Filamenti Intermedi
39
Adattato da Smith and Wood: Cell Biology, Chapman & Hall, London, 1992
Coulombe PA, Omary MB. 'Hard' and 'soft' principles defining the structure, function and regulation of keratin intermediate filaments. Curr Opin Cell Biol. 14:110‐122, 2002.
Le cheratine costituiscono il maggiore sotto‐gruppo di proteine dei filamenti intermedi e rappresentano le proteine più abbondanti delle cellule epiteliali. Si presentano sotto forma di rete dinamiche di filamenti citoplasmatici di 10‐12 nm che sono eteropolimeri obbligati di cheratine di tipo I e di tipo II.
La principale fonzione delle cheratine è quella di proteggere le cellule epiteliali da stress meccanici e non meccanici che provocherebbero la morte cellulare.
Altre fonzioni emergenti includono ruoli nel segnalamento cellulare, nella risposta allo stress e nell’apoptosi, nonchè ruoli specifici e unici che sono cheratina‐ e tessuto‐specifici. Il ruolo delle cheratine in un gran numero di patologie della pelle, capelli, occhi, e fegato è ora stabilito e si combina bene con le prove sperimentali raccolte con modelli di topi transgenici.
I fenotipi associati a difetti nelle proteine della famiglia delle cheratine sono oggetto di modulazione significativa mediante ridondanza funzionale all’interno della famiglia e dei geni modificanti. I filamenti di cheratina subiscono regolazione complessa che coinvolge modificazioni post‐traduzionali e interazioni con loro stesse o con diverse classi di proteine associate. Filamenti Intermedi - BUSB2011
7
26/05/2013
Citocheratine: epidermide, peli, capelli, lana, zoccoli, artigli
CITOCHERATINE - 1
(N.B. Escludendo le cheratine di tipo “hair”, o “dure”, presenti nei peli,
zoccoli, artigli, ecc.
Gruppo più vasto e più complesso dei filamenti intermedi.
E’ composto di molti polipeptidi diversi, distribuiti nelle cellule normali degli epiteli e nelle cellule neoplastiche di origine epiteliale
La gel elettroforesi delle citocheratine estratte dai tessuti epiteliali e dalle cellule epiteliali in cultura ha rivelato una famiglia di 19 polipeptidi che variano in peso molecolare da 40 kD a 68 kD. In seguito, è stata inclusa nella famiglia delle citocheratine la citocheratina 20 (46 kD), nota anche come “citocheratina IT”.
Tutte le citocheratine sono state divise in due sottofamiglie, le citocheratine di tipo I, che sono più acidiche, e le citocheratine di tipo II, più basiche.
I membri delle subfamiglie di tipo I e di tipo II sono di solito espressi in coppia nei vari tipi di epiteli; ogni citocheratina di tipo I ha una correspondente citocheratina di tipo II.
La subfamiglia di tipo II consiste nelle citocheratine ad alto peso molecolare, dalla CK1(68 kD) fino alla CK6 (56 kD), che stanno entro il range di pH 7.8 a 7.2.
La subfamiglia di tipo I comprende le citocheratine a basso peso molecolare, dalla CK9 (64 kD) alla CK 19 (40kD), con pH da 5.4 a 5.2.
Le citocheratine CK7 (54 kD) e CK 8 (52.2 kD) sono considerate di tipo e carica elettrica intermedio, con pH 6.0‐6.1.
La citocheratina CK20 (46 kD) ha pH 6.1.
Adattato da Newsletter della Biomeda, Summer 1996, Volume 1, Issue 1
CITOCHERATINE - 2
CITOCHERATINE ‐ 3
Il contenuto in citocheratine dei vari tessuti dipende dal tipo di tessuto e dal grado di differenziamento delle cellule. Citocheratine primarie: compaiono negli epiteli normali nei primi stadi dello sviluppo
Citocheratine secondarie: espresse negli epiteli più differenziati.
Le citocheratine CK8 e CK18 sono le citocheratine primarie per gli epiteli semplici.
Le citocheratine CK5 e CK 14 sono primarie per gli epiteli stratificati; si trovano negli strati basali dei vari tipi di epiteli squamoso e non‐squamoso. Tutti gli altri membri della famiglia delle citocheratine sono secondarie; sono espresse negli strati soprabasali dei vari epiteli.
I polipeptidi di citocheratine sono di solito conservati nei derivati tumorali delle cellule epiteliali normali.
La definizione immunoistochimica del profilo di citocheratine dei tumori è estremamente utile perchè permette ai ricercatori di dimostrare l’origine epiteliale dei tumor indifferenziati, nonchè di risalire all’origine istologica dei tumori di origine epiteliale.
Il contenuto in citocheratine dei vari tessuti dipende dal tipo
di tessuto e dal grado di differenziamento delle cellule. Ci sono citocheratine primarie che compaiono negli epiteli normali nei primi stadi dello sviluppo, e citocheratine secondarie che sono espresse negli epiteli più differenziati.
Le citocheratine CK8 e CK18 sono le citocheratine primarie per gli epiteli semplici.
Le citocheratine CK5 e CK 14 sono primarie per gli epiteli stratificati; si trovano negli strati basali dei vari tipi di epiteli squamoso e non‐squamoso. Tutti gli altri membri della famiglia delle citocheratine sono secondarie; sono espresse negli strati soprabasali dei vari epiteli.
I polipeptidi di citocheratine sono di solito conservati nei derivati tumorali delle cellule epiteliali normali.
ESPRESSIONE DI FILAMENTI INTERMEDI
DELLA FAMIGLIA DELLE CHERATINE
IN DIVERSI TESSUTI EPITELIALI
Smith and Wood: Cell Biology, Chapman & Hall, London, 1992
Filamenti Intermedi - BUSB2011
http://jcs.biologists.org/content/124/24/4221/F7.large.jpg
8
26/05/2013
Appaiamento tra cheratine acide e basiche
http://en.wikipedia.org/wiki/Keratin
Desmina nel muscolo
http://www.cellsignal.com/products/image
s/4024_iff_md_071214.jpg
Filamenti intermedi
Questi filamenti intermediari di tipi III avvolgono il disco Z e fanno
contatti ulteriori con i circostanti dischi Z della stessa miofibrilla. L’allineamento dei filamenti di desmina con il sarcomero del muscolo è mantenuto da numerose proteine associate ai filamenti intermedi
(IFAPs, “intermediate filaments‐associated proteins”), che includono la schelemina nella linea M e la sinemina nel disco Z. DESMINA
http://www.ncbi.nlm.nih.gov/books/NBK21560/figure/A5561/
Architettura molecolare di un miocito, in cui sono evidenziate le proteine coinvolte nelle miopatie scheletriche e cardiache
La desmina è la principale proteina dei FI del muscolo. Interagisce con altre proteine per sostenere le miofibrille a livello dei dischi Z e forma una rete continua di citoscheletro che mantiene i rapporti spaziali fra l’apparato contrattile e gli altri elementi strutturali della cellula. La desmina mantiene l’integrità cellulare, la trasmissione di forza, e il segnalamento meccanochimico. Mutazioni in altre proteine sarcomeriche e citoscheletriche (plectina, miotilina, filamina, αB‐cristallina, «Z band alternatively spliced PDZ‐motif protein [ZASP]», e «BCL2‐associated athanogene 3 [BAG3]» provocano disordini neuromuscolari.)
Filamenti intermedi
VIMENTINA
Tragedy in a heartbeat: malfunctioning desmin causes skeletal and cardiac muscle disease Lev G. Goldfarb, Marinos C. Dalakas J Clin Invest. 2009; 119(7):1806–1813 Filamenti Intermedi - BUSB2011
9
26/05/2013
Vimentina
Vimentina: cellule di origine mesenchimale
The vimentin intermediate filament network (red) extends into the lamella where tetramers colocalize with the actin‐bundling protein, fimbrin (green). These fluorescent foci are specialized focal adhesions found in osteoclasts and macrophages called podosomes.
Vimentin is an intermediate filamentous cytosketetal protein found in cells of mesenchymal origin. Vimentin typically exists as a coiled‐coil dimer that interacts with another dimers to form a tetrameric complex that forms cytoskeletal protein sheets. Vimentin intermediate filaments are believed to be important cytosolic structures responsible for positioning organelles. It has been found to interact with the Golgi, ER, mitochondria and nucleus.
http://www.sigmaaldrich.com/life-science/metabolomics/enzyme-explorer/learning-center/structuralproteins/vimentin.html
http://web.wi.mit.edu/matsudaira/pub/fimbrin.shtml
Vimentina: cellule di origine mesenchimale
Filamenti intermedi
SISTEMA NERVOSO
http://www.sigmaaldrich.com/prodimages/c/c1801‐if.jpg
Le principali proteine
dei filamenti intermedi
dei neuroni e cellule
gliali
http://www.jci.org/articles/view/38003/figure/1
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bnchm&part=A569&rendertype=table&id=A578
Filamenti Intermedi - BUSB2011
10
26/05/2013
Didascalia figura precedente
Aspetti morfologici di aggregati proteici gliali e neuronali
nella malattia di Alexander e Charcot-Marie-Tooth
disease.
• A “typical” IF protein consists of an N‐terminal head region, a rod region that contains four α‐helical regions (helices 1A, 1B, 2A, and 2B), and a C‐
terminal tail region. Different isoforms of GFAP are shown: GFAP‐α and GFAP‐β are full‐length GFAP proteins; the mRNAs encoding these two proteins differ in the 5′ UTR. GFAP‐γ is encoded by an mRNA that has an alternate start site and is missing exon 1. The mRNAs encoding GFAP‐δ, GFAP‐ε, and GFAP‐κ are generated by alternative splicing of intron 7, with variable use of exon 7+, to create proteins with different C‐terminal tail sequences. Only single isoforms of α‐internexin, NFL, NFM, and NFH are shown. The gray areas in NFM and NFH contain multiple phosphorylation sites. Full‐length peripherin (Per58) is shown as well as Per61, a mouse isoform that retains intron 4 (96), and Per28, which is a mouse and human isoform that retains introns 3 and 4, resulting in a truncated peripherin
(97).
Liem RK, Messing A. Dysfunctions of neuronal and glial intermediate filaments in disease. J Clin Invest. 2009 Jul;119(7):1814-24.
Liem RK, Messing A. Dysfunctions of neuronal and glial intermediate filaments in disease. J Clin Invest. 2009 Jul;119(7):1814-24.
Didascalia figura precedente
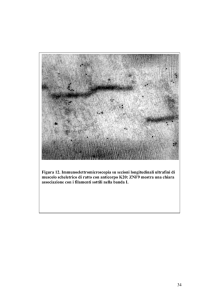
• (A) Rosenthal fibers concentrated in the astrocytic endfeet surrounding a blood vessel (V) in the brain stem of a 1‐year‐old child with Alexander disease. H&E stain, paraffin section (reproduced with permission from Elsevier [102]). Original magnification, ×62. (B) Rosenthal fibers surrounded by IFs in an astrocyte cell body from a 17‐month‐old child with Alexander disease, viewed by transmission electron microscopy (reproduced with permission of Wiley‐Liss Inc., a subsidiary of John Wiley & Sons Inc. [103]). N, nucleus. (C) Sural nerve biopsy from a CMT patient with an L286P mutations in NFL. The figure shows a giant axon with a cluster of organelles (arrow) and irregular whorls of neurofilaments. (D) Sural nerve biopsy of a CMT patient with an NFL del322C–326N mutation. The figure shows a fiber whose axoplasm consists almost exclusively of microtubules; note the loosening of the external myelin lamellae (panels C and D were reproduced with permission from Brain: a journal of neurology [79]). Scale bars: 1 μm (B); 2 μm (C and D). Filamenti intermedi
NEUROFILAMENTI
Principali funzioni cellulare dei filamenti intermedi citoplasmatici ‐2
Neurofilamenti
Confocal immunofluorescent image of mouse cerebellum labeled with Neurofilament‐
H (RMdO 20) Mouse mAb (green) and Calbindin Antibody #2136 (red). Blue pseudocolor = DRAQ5™ (fluorescent DNA dye).
http://www.cellsignal.com/products/2836.html
Filamenti Intermedi - BUSB2011
Citoarchittetura. Nei motoneuroni, la crescita radiale dei processi assonali richiede la loro
interazione con neurofilamenti (blu chiaro) in modo da trovare la corretta stechiometria fra le subunità “light” (NF‐L), media (NF‐M) e “heavy” (NF‐H). I grandi domini della coda C‐terminale delle
subunità NF‐H (rosso) e NF‐M (arancione) sono iperfosforilati e si proiettano al di fuori della zona
centrale del filamento, così determinando la spaziatura fra I filamenti e il calibro dell’assone. Gli
abbondanti neurofilamenti interagiscono con I microtubuli meno abbondanti (blu scuro) e con I filamenti di actina subcorticali (verde scurto) mediante proteine linker del citoscheletro, quali la plectina e la BPAG1 (giallo). Un ruolo citoarchitteturale è stato inoltre dimostrato per le lamine
nucleari ed altri filamenti intermedi citoplasmatici quali la desmina e le cheratine. 11
26/05/2013
Neurofilamenti
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bnchm&part=A569&rendertype=table&id=A578
http://www.encorbio.com/cpolyclonal/CPCA‐NF‐H.htm
Didascalia della figura sul citoscheletro degli assoni e dei dendriti
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=bnchm&part=A569&rendertype=figure&id=A574
I citoscheletri degli assoni e dei dendriti differiscono sia in composizione che in organizzazione.
Le principali differenze sono illustrate diagrammaticamente in questa Figura. Con una eccezione, tutte
le proteine citoscheletriche sono sintetizzate su polisomi liberi nel corpo cellulare, e successivamente
trasportate ai loro differenti compartimenti cellulari. L’eccezione è la MAP2, che è la principale proteina
associata ai microtubuli dei dendriti. Mentre una certa parte della MAP2 è sintetizzata nel corpo
cellulare, il mRNA per la MAP2 è specificamentente arricchito nel compartimento dendritico e si ritiene
che una frazione significativa sia sintetizzata in quel sito. Si crede che i microtubuli dei corpi cellulari,
dei dendriti e degli assoni siano nucleati nel centro organizzatore dei microtubuli (MTOC), e quindi
rilasciati e consegnati ai dendriti oppure all’assone. Nel dendrite, i microtubuli spesso hanno polarità
miste con sia estremità le “meno” che le “più” presenti nella parte distale rispetto al corpo cellulare. La
conseguenza funzionale di tale organizzazione è incerta ma potrebbe aiutare a spiegare come mai i
dendriti assumono una forma affusolata mentre si allontano dal corpo cellulare. Vice-versa, tutti i
microtubuli degli assoni sono orientati con l’estremità “più” in posizione distale rispetto al corpo
cellulare ed esibiscono una distribuzione uniforme lungo l’assone. Nonostante una certa quantità di
proteina tau possa essere identificata nei corpo cellulari e nei dendriti, i microtubuli assonali sono
arriccchiti in tau e la tau assonale è fosforilata in modo diverso. La MAP2 sembra essere assente
dall’assone. I neurofilamenti sono in gran parte esclusi dai compartimenti dendritici ma sono
abbondanti negli assoni di grandi dimensioni. Il distanziamento dei neurofilamenti è sensibile al livello
di fosforilazione. Sia i microtubuli che i neurofilamenti si fermano ed iniziano nell’assone piuttosto e non
proseguono nel corpo cellulare. I microfilamenti hanno un’organizzazione più dispersa e possono
essere di difficile visualizzazione nel neurone maturo. Essi sono principalmente abbondanti vicino alla
membrana plasmatica ma sono anche molto abbondanti nei terminali pre-sinaptici e nelle spine
dendritiche. GA: apparato di Golgi.
Glial Fibrillary Acidic Protein (GFAP)
Filamenti intermedi
GLIAL FIBRILLARY ACIDIC PROTEIN
http://encorbio.com/cpolyclonal/CPCA‐GFAP.htm
Filamenti Intermedi - BUSB2011
12
26/05/2013
Filamenti intermedi
NESTIN
http://en.wikipedia.org/wiki/Nestin_%28protein%29
http://www.pnas.org/content/98/8/4758/F1.large.jpg
http://en.wikipedia.org/wiki/Nestin_%28protein%29
http://en.wikipedia.org/wiki/Nestin_%28protein%29
Filamenti intermedi
DESMOSOMI, EMIDESMOSOMI
http://www.globalstem.com/human‐iPSC‐derived‐neural‐cells
Filamenti Intermedi - BUSB2011
13
26/05/2013
Filamenti intermedi
This schematic of the epidermis shows that proteins are expressed in a differentiationdependent manner. For example, in the case of desmosomal cadherins, desmoglein (Dsg) 1
and desmocollin (Dsc) 1 are patterned in a gradient, with the highest level in the superficial
layers and tapering off in the basal layers. Desmoglein 3 and desmocollin 3 show an opposite
pattern. Likewise, the armadillo family member plakophilin (PKP) 1 is concentrated in
junctions of the superficial layers, whereas PKP2 is found in desmosomes deeper in the
epidermis.
APPLICAZIONE IN ONCOLOGIA E BIOLOGIA DELLO SVILUPPO
http://www.nature.com/nrm/journal/v1/n3/fig_tab/nrm1200_208a_F3.html
DIAGNOSI DEI TUMORI UMANI IN BASE
ALLA LORO ESPRESSIONE DI FILAMENTI INTERMEDI - 1
Normalmente quando i pazienti con tumori sono trattati con radiazione o farmaci, la decisione sul tipo di trattamento è basata sulla diagnostica del tumore.
Per esempio, alcuni tipi tumorali respondono al trattamento con la radizione mentre altri sono resistenti.
Man mano i tumori crescono le caratteristiche strutturali del tessuto normale scompaiono e le singole cellule spesso perdono le loro carateristiche proteine di membrana, il che rende la loro identificazione molto difficile.
Tuttavia, tumori di origine sconosciuta possono essere diagnosticati mediante l’identificazione dei loro filamenti intermedi con anticorpi specifici. Questo perchè l’espressione dei filamenti intermedi è cellula‐specifica, ossia, ogni proteina di filamento intermedio è associata ad un tipo cellulare particolare.
Ad esempio, i tumori che esprimono citocheratine possono essere classificati come carcinomi (di origine epiteliale) e distinti dai sarcomi (di origine mesenchimale) che esprimono vimentina.
Filamenti Intermedi - BUSB2011
DIAGNOSI DEI TUMORI UMANI IN BASE
ALLA LORO ESPRESSIONE DI FILAMENTI INTERMEDI - 2
Il successo di questo metodo dipende moltissimo dall’assoluta specificità dell’anticorpo per un determinato tipo di filamento intermedio.
Un altro fattore critico per la diagnostica è che le cellule tumorali non sempre ubidiscono alle “regole” dell’espressione tissutale. E’ stato dibattuto vivacemente ed ora accettato che, al contrario delle cellule normali non cancerose, le cellule tumorali possono co‐esprimere due tipi di filamenti intermedi, ad esempio citocheratine e vimentina. Tuttavia, queste non co‐
polimerizzano in filamenti, ma esistono come reti separate.
Se le limitazioni di cui sopra sono tenute in considerazione, l’espressione dei filamenti intermdi dai tumori rimane un metodo per la loro diagnosi. (Adattato da: Smith and Wood: Cell Biology, Chapman & Hall, London, 1992)
14
26/05/2013
Immunohistochemical tumor differentiation (2)
Immunohistochemi
cal tumor differentiation (1)
BUSB 2011 ‐ Filamenti Intermedi
85
BUSB 2011 ‐ Filamenti Intermedi
86
FILAMENTI INTERMEDI & DIFFERENZIAMENTO
CELLULE DEI MAMMIFERI
Filamenti intermedi
COLLEGAMENTO CON ALTRE STRUTTURE CITOSCHELETRICHE
(Adattato da: Smith and Wood: Cell Biology, Chapman & Hall, London, 1992)
Didascalia della figura di Herrmann
Rappresentazione schematica dell’interazione fra I microfilamenti (MF), I microtubuli (MT) e I filamenti
intermedi (IF) che coinvolge la proteina plectina con I suoi domini terminali che legano l’actina (ABD) e
I microtubuli (MBD). Mentre il MBD della plectina non è ancora stato definito in termini molecolari, è
stato chiaramente visualizzato mediante microscopia elettronica. Per la BPAG1, un’altra plachina,
l’esistenza di un MBD è stato direttamente dimostrata. E’ stato dimostrato che il dominio carbossiterminale della plectina si lega ai Ifs direttamente, probabilmente mediato da un processo di
fosforilazione. In quest’ultimo contesto, si è visto che delle chinasi strettamente associate al
citoscheletro fosforilano la plectina in modo altamente specifico sia in vitro che in vivo. Per quanto
riguardano gli IF, è stato evidenziato un eterodimero vimentina (giallo)/nestina (blu). Il lungo carbossi.terminale non ad α-elica potrebbe ospitare sequenze di tipo sia MBD che ABD, così legando in modo
stabile gli Ifs ai MTs e/o ai MFs. Come si può inferire da lavoro di Prahlad et al. [1998] and Yoon et al.
[1998], le chinesino possono collegare gli Ifs ai MTs per facilitare le interazioni dinamiche. (X, Y e Z)
marcano altri potenziali motivi di legame all’interno del dominio pre-bastocello nell’amino-terminale
della plectina e nestina, rispettivamente. Il ABD e MBD della nestina sono are evidenziati da un punto
interrogativo che indica che I corrispondenti motivi di legame non sono ancora stati identificati. I
diametri relativi dei tre filamenti citoscheletrici sono stati disegnati in scala approssimata. I segmenti
“coiled-coil” della chinesina e della plectina sono stati disegnati in modo incompleto come indicato
dalla line atrattegiata doppia.
Herrmann H, Aebi U. Intermediate filaments and their associates: multi-talented structural elements specifying
cytoarchitecture and cytodynamics. Curr Opin Cell Biol. 12(1):79-90, 2000.
Filamenti Intermedi - BUSB2011
15
26/05/2013
Didascalia della figura di Herrmann, 2009:
Molecular organization of IF proteins.
(A) Schematic representation of the structural organization of an IF molecule.
Blue boxes represent segments; orange boxes represent IF-consensus motifs
(CD); and the yellow box represents an α-helical pre-coil domain only found in
vimentin-like IF proteins. L1, L12, and L2 are linker segments. As L2 is
predicted to form an α-helical segment, coil 2A and linker L2 are designated
as paired bundle (pb) in B. “Head” and “tail” represent non–α-helical
domains. The stutter (st) indicates a discontinuity in the heptad repeat
pattern of coil 2B (43). The blue box designated lamin-specific “insertion”
highlights a position in which an additional α-helical segment is found in
lamins and lower invertebrate cytoplasmic IF proteins.
(B) Model of the dimeric structures exhibited by human lamin A, vimentin,
and the epidermal pair K5/K14. Vimentin exhibits a pre-coil domain (PCD).
Moreover, a special feature of nuclear lamins, as opposed to cytoplastic IF
proteins, is that linker L1 in lamins is likely to be α-helical. The numbers in
parentheses indicate the number of amino acids in the respective non–αhelical domains. NLS, nuclear localization signal.
Lamina Nucleare & Lamìne
http://micro.magnet.fsu.edu/cells/nucleus/nuclearpores.html
http://www.nature.com/nrg/journal/v8/n2/fig_tab/nrg2041_F1.html#figure-title
http://www.abcam.com/index.html?pageconfig=resource&rid=11025&pid=10026
Lanctôt et al.. Dynamic genome architecture in the nuclear space: regulation of gene expression in three dimensions.
.
Nat Rev Genet. 8:104-115, 2007
Involucro nucleare (1)
Modello di assemblaggio delle lamìne
La scorsa decade ha visto un complete ripensamento della tradizionale visione dell’involucro nucleare come soltanto un involucro passivo per i cromosomi.
La convergenza di diverse linee di ricerca di base e clinica ha rivelato ruoli addizionali sia nel segnalamento che nella progressione mitotica. I polipeptidi delle lamìne formano dimeri in cui le regioni centrali ad α-elica si
avvolgono una rispetto all’altre. Ulteriore assemblaggio può coinvolgere
un’associazione testa-coda dei dimeri per formare polimeri lineari e associazioni
laterali di polimeri per formare filamenti.
http://www.ncbi.nlm.nih.gov/books/NBK9927/figure/A1328/
Filamenti Intermedi - BUSB2011
Sta diventando evidente che l’involucro nucleare definisce
non solo l’organizzazione nucleare ma anche quella del citoscheletro e in questo modo, integra sia l’archiettetura nucleare che quella citoplasmatica. (Stewart et al., Science, 2007)
16
26/05/2013
Lamina nucleare
La membrana nucleare interna contiene diverse proteine integrali, tra cui
l’emerina e il recettore per la lamìna B (LBR) che interagiscono con le lamìne
nucleari. Le lamìne interagiscono inoltre con la cromatina
Lamina Nucleare
(Scanning Electron Microscopy)
Lamina nucleare
http://userpage.chemie.fu-berlin.de/biochemie/agknaus/SNE.html
Involucro Nucleare (2)
L’involucro formato da due membrane è penetrate dai pori nucleari ed è continuo
con il reticolo endoplasmico. I ribosomi, che sono legati alla superficie citosolica
dela membrane del RE e alla membrana nucleare esterna, non sono evidenziati
Schema di una molecola di lamìna
La regione interna elicoidale è lunga circa 50 nm ed è fiancheggiata da domini non elicoidali.
I triangoli indicano siti di fosforilazione.
NLS: “Nuclear Localization Signal”
CaaX box: sito di modificazioni post‐traduzionali.
Foisner R. Inner nuclear membrane proteins and the nuclear lamina. J Cell Sci. 114:3791-3792, 2001.
Freccia: indica sito specifico di proteolisi della lamìna A.
(Smith & Wood, Cell Biology, Stanley Thornes, Cheltenham, 2nd edition, 1999).
Filamenti Intermedi - BUSB2011
17
26/05/2013
Modello di assemblaggio delle lamìne
I polipeptidi delle lamìne formano dimeri in cui le regioni centrali ad α-elica si
avvolgono una rispetto all’altre. Ulteriore assemblaggio può coinvolgere
un’associazione testa-coda dei dimeri per formare polimeri lineari e associazioni
laterali di polimeri per formare filamenti.
http://www.ncbi.nlm.nih.gov/books/NBK9927/figure/A1328/
Proprietà generali delle lamine nucleari
Le lamine nucleari sono state inizialmente descritte come componenti proteiche principali delle frazioni altamente resistenti al trattamento con i detergenti della “lamina” di nuclei di fegato di ratto e di eritrociti di pollo. In seguito, è stato visto che esse sono membri della famiglia di proteine dei Filamenti Intermedi.
I geni delle lamine si trovano in tutti i metazoi finnora osservati, ma sono assenti nelle piante e negli organismi unicellulari.
Le lamine sono suddivise in tipi A e B in base a omologie di sequenza.
Nei mammiferi sono stati caratterizzati due principali tipi di lamìna A (lamìne A e C) e due principali tipi di lamìne B (lamìne B1 e B2), oltre alle isoforme minori lamina A_10 e lamina C2 specifica delle cellule germinali. Mentre le lamìne B1 e B2–B3 sono codificate da geni diversi (LMNB1 e LMNB2, rispettivamente), le lamine di tipo A sono derivate per splicing alternativo da un singolo gene (LMNA).
Dechat T, Pfleghaar K, Sengupta K, Shimi T, Shumaker DK, Solimando L, Goldman RD. Nuclear lamins: major
factors in the structural organization and function of the nucleus and chromatin. Genes Dev. 22: 832-853,
2008
Lamina nucleare & Lamìne - 1
La lamina nucleare è compostada un gruppo di proteine chiamate lamìne A . B e C che hanno un PM di circa 60‐70 kD.
Il gruppo A è espresso sopratutto nelle cellule differenziate
Le lamìne B sono prodotte costitutivamente
Le lamìne C sono identiche ai tipo A. ad eccezione di un’estensione di ulteriori 90 AA, che deriva dllo splicing alternativo del prodotto genico nelle cellule dei mammiferi.
Si conoscono diversi tipi di lamìne B, tutti prodotti di geni diversi. (Smith & Wood, Cell Biology, Stanley Thornes, Cheltenham, 2nd edition, 1999).
Lamina nucleare
La membrane interna dell’involucro nucleare si trova vicino ad uno strato di filamenti sottili che circonda il nucleo ovunque tranne che all’altezza dei pori nucleari. Questi possono anche fungere da filamenti di stabilizzazione. Questa struttura è detta lamina nucleare. Ha le seguenti caratteristiche strutturali e funzionali: Consiste di "filamenti intermedi ", di spessore 30‐100 nm.
Questi filamenti intermedi sono polimeri di lamìne, con pesi molecolare 60‐75 kD .
Le lamìne di tipo‐A si trovano all’interno, verso il nucleoplasma. Le lamìne di tipo B si trovano vicino alla membrana nucleare (interna) e possono legarsi a proteine integrali all’interno della membrana. Le lamìne possono essere coinvolte nell’organizzazione strutturale del nucleo.
Esse possono giocare un ruolo nell’assemblaggio e disassemblaggio prima e dopo la mitosi. La loro fosforilazione provoca il disassemblaggio della lamina e provoca la disgregazione dell’involucro nucleare in vescicole. La defosforilazione rovescia questo processo e permette al nucleo di riformarsi. Se si iniettano anticorpi contro le lamine nelle cellule, il nucleo non si può riformare dopo la divisione. Quindi le lamine sono essenziali per il riassemblaggio. Lamìne A e B
Negli scorsi anni è diventato evidente che proteine della famiglia dei
filamenti intermedi, le lamìne A e B, non solo forniscono un’impalcatura
strutturale per il nucleo, ma sono inoltre essenziali per molti aspetti
delle funzioni normali del nucleo.
Conoscenze più approfondite sulle funzioni delle lamine sono state derivate
da studi del numero notoriamente elevato di mutazioni patogene nel gene
delle lamìne.
Questo review fornisce una panoramica aggiornata delle funzioni delle lamine nucleari,
enfatizzando il loro ruolo nell’epigenetica, organizzazione della cromatina, replicazione del DNA,
trascrizione e riparo del DNA. Inoltre, discutiamo prove recenti che supportano l’importanza delle
lamine nelle infezioni virali.
Dechat T, Pfleghaar K, Sengupta K, Shimi T, Shumaker DK, Solimando L, Goldman RD.
Nuclear lamins: major factors in the structural organization and function of the
nucleus and chromatin. Genes Dev. 22: 832-853, 2008
Lamina nucleare & Lamìne - 2
Le lamìne appartengono alla famiglia di proteine dei Filamenti Intermedi e presumibilmente tutte queste proteine sono evolute da un antenato comune.
Le lamìne hanno le caratteristiche molecolare strutturali tipiche della famiglia dei FI:
Una regione interna di PM 40 kD contenente circa 350 residui di AA che formano un dominio ad α‐elica a forma di bastoncino.
Questo dominio è fiancheggiato da regioni non‐elicoidali ad entrambe le estremità N‐ e C‐ terminali di PM 40 kDa e 20‐30 kDa, rispettivamente.
Le lamìne B differiscono dalle lamìne A in quanto hanno una porzione C‐terminale più piccola.
Questi dimeri si aggregano per formare lunghe fibre con diametro di circa 10 nm, che si associano per formare la rete della lamina nucleare.
(Smith & Wood, Cell Biology, Stanley Thornes, Cheltenham, 2nd edition, 1999)
Filamenti Intermedi - BUSB2011
18
26/05/2013
Lamina nucleare & Lamìne - 3
Le lamìne differiscono dalle proteine citoplasmatiche dei FI in quanto hanno un segnale di localizzazione nucleare, e una cosidetta “CaaX box” vicino al C‐terminale.
La box è il sito di tre successive modificazioni post‐
traduzionali:
(a) Isoprenilazione di un residuo di cisteina
(b) La rimozione proteolitica di tre residui di AA nel C‐
terminale
(c) La cabossimetilazione del residuo di cisteina isoprenilato.
Queste modificazioni aumentano l’idrofobicità del C‐
terminale e sono necessarie per indirizzare le lamìne di nuova sintesi verso la membrana nucleare interna.
Siti potenziali di fosforilazione sono presenti ad entrambe le estremità della regione elicoidale.
(Smith & Wood, Cell Biology, Stanley Thornes, Cheltenham, 2nd edition, 1999)
FRAMMENTAZIONE DELL’INVOLUCRO NUCLEARE
Lamina nucleare & Lamìne - 4
Le lamìne di tipo A sono soggette a proteolisi specifica che rimuove il C‐
terminale modificato post‐traduzionalmente, ma le lamìne B trattengono questa estremità modificate e si sa che legano un gruppo di proteine sulla faccia nucleare della membrana nucleare interna chiamata complesso p58.
La proteina p58 (PM 58 kDa) legante la lamìna B forma un oligomero con i polipeptidi p18.
Si ritiene che sia p58 che p18s siano proteine integrali della membrana nucleare interna.
Altre proteine del complesso includono una p58 chinasi, p34 e p150 (PM 34 e 150 kDa, rispettivamente) e le lamìne A e B che probabilmente sono tutte proteine periferiche di membrana.
Non si hanno informazioni specifiche su come le lamine del compleso interagiscono con la cromatina. Si sa che in vivo la lamìna A in particolare si lega alla cromatina e che queste interazioni coinvolgono i C‐terminali delle lamìne e le fibre cromatiniche di 30 nm.
(Smith & Wood, Cell Biology, Stanley Thornes, Cheltenham, 2nd edition, 1999)
ASSEMBLAGGIO DELL’INVOLUCRO NUCLEARE
DURANTE LA TELOFASE
Vescicole dell’involucro nucleare, generate dalla
rottura dell’involucro durante la profase, si
associano con i cromosomi in fase di decondensazione e quindi si fondono. I complessi dei subpori si riassemblano nei pori
nucleari, formando mininuclei individuali detti
cariomeri. Mentre la lamina nucleare si dissocia, l’involucro nucleare si frammenta in vescicole. Le lamìne di tipo B rimangono legate a queste vescicole, mentre le lamine A e C vengono rilasciate come dimeri liberi.
I cromosomi da essi racchiusi si decondensano
ulteriormente e la subseguente fusione degli
involucri nucleari di tutti i cariomeri all’altezza dei
poli del fuso forma un nucleo singolo contenente
un corredo completo di cromosomi. Il riassemblaggio della lamina nucleare non è indicato.
http://www.ncbi.nlm.nih.gov/books/NBK21704/figure/A3523/
Filamenti Intermedi - BUSB2011
19
26/05/2013
Rottura e ri‐formazione dell’involucro nucleare
durante la mitosi
Dissoluzione della lamina nucleare
La lamina nucleare consiste in una rete di filamenti di lamina. Nella mitosi, la Cdc2 ed altre proteina chinasi fosforilano le lamine, provocando la dissociazione dei filamenti in dimeri di lamina liberi.
http://www.ncbi.nlm.nih.gov/books/NBK9890/figure/A1377/?report=objectonly
LA FOSFORILAZIONE DELLE LAMINE PORTA ALLA DEGRADAZIONE DELL’INVOLUCRO NUCLEARE ‐1
LA FOSFORILAZIONE DELLE LAMINE PORTA ALLA
DEGRADAZIONE DELL’INVOLUCRO NUCLEARE -2
L’involucro nucleare è una estensione circondata da una doppia membrana del reticolo endoplasmico ruvido che contiene molti complessi dei pori nucleari. Il doppio strato lipidico della membrana interna è sostenuto dalla lamina nucleare, una rete di filamenti di lamine localizzato sotto la faccia interna dell’involucro nucleare. Le tre lamìne nucleari (A, B, e C) presenti nelle cellule dei Vertebrati appartengono alla classe dei filamenti intermedi
delle proteine del citoscheletro, che sono cruciali per sostenere le membrane cellulari.
Le lamìne A e C, che sono codificate dalla stessa unità di trascrizione e prodotte per splicing alternativo di un singolo pre‐mRNA, sono identiche, all’eccezione di una regione di 133 residui nel C‐ terminale della lamìna A, che è assente nella lamìna C.
La lamìna B, codificata da una unità trascrizionale diversa, viene modificata post‐traduzionalmente mediante l’aggiunta di un gruppo isoprenilico idrofobico vicino al C‐ terminale. Questo acido grasso viene incorporato nel foglietto interno del bilayer lipidico che forma la membrana nucleare interna, ancorando così la lamina nucleare alla membrana. Tutte tre le lamìne nucleari formano dimeri che contengono una sezione centrale bastoncellare ad α‐elica e domini di testa e di coda globulari; la polimerizzazione di questi dimeri mediante associazioni testa‐a‐testa e coda‐a‐coda genera i filamenti intermedi che compongono la lamina nucleare.
LA FOSFORILAZIONE DELLE LAMINE PORTA ALLA DEGRADAZIONE DELL’INVOLUCRO NUCLEARE ‐3
All’inizio della mitosi, il Mitosis Promoting Factor (MPF) fosforila residui di serina specifici in tutte tre le lamine, provocando la depolimerizzazione dei filamenti intermedi della lamina. I dimeri fosforilati di lamìne A e C vengono rilasciati in soluzione, mentre i dimeri fosforilati di lamìne B rimangono associati alla membrana nucleare mediante la loro ancora isoprenilica.
La depolimerizzazione delle lamìne nucleari porta alla disintegrazione della rete della lamina nucleare e contribuisce alla rottura dell’involucro nucleare in piccole vescicole. Ri‐formazione dell’involucro
nucleare
Il primo passo del riassemblaggio
dell’involucro nucleare consiste nel
legame di vescicole di membrana
ai cromosomi, che può essere
mediato sia da proteine integrali di membrana che da lamìne di tipo B. Le vescole allora si fondono, la lamina nucleare si riassembla e i
cromosomi decondensano. http://www.ncbi.nlm.nih.gov/books/NBK9890/figure/A13
82/
Filamenti Intermedi - BUSB2011
20
26/05/2013
Laminopatie
Mutazioni in uno dei geni delle lamine (LMNA) sono responsabili di diverse patologie umane, incluse una rara forma di distrofia muscolare, la distrofia muscolare di Emery‐Dreyfuss autosomica dominante (EDMD2) in cui le cellule muscolari contegnono nuclei estremamente fragili.
Anche mutazioni a carico della lamìna A sono state correlate ad una patologia, sindrome della progeria di Hutchinson‐Gilford (HGPS), caratterizata da invecchiamento precoce e morte durante l’adolescenza per infarto o ictus.
I nuclei deformi in questa patologia sono stati attribuiti ad una mutazione sinonima (mutazione che hagenerato un diverso codone codificante per lo stesso AA). Il cambiamento di sequenza di DNA ha alterato la modalità di splicing del trascritto genico, portando alla produzione di una proteina più corta.
Filamenti Intermedi
LAMINOPATIE
La sequenza di un gene quindi rappresenta un “doppio codice”, uno che dirige il machinario della traduzione ed un altro che dirige il macchinario di splicing.
(Karp, Biologia Cellulare e Molecolare, 3a ed)
Didascalia Fig.1 Worman
Figure 1
• The nuclear lamina. (A) The nuclear lamina is a meshwork of IFs localized
primarily to the nucleoplasmic face of the inner nuclear membrane (shown
schematically in red). The lamins interact with several integral proteins of
the inner nuclear membrane, including lamin B receptor (LBR), MAN1
(encoded by the LEMD3 gene), emerin, lamina-associated polypeptide 1
(LAP), LAP2β, small nesprin 1 isoforms, and SUNs. SUNs interact with
large nesprin 2 isoforms, integral proteins of the outer nuclear membrane,
which also interact with actin, linking the nuclear lamina to the cytoskeleton.
(B) In humans, 3 genes encode nuclear lamins. LMNA on chromosome
1q21.2 encodes the A-type lamins, with prelamin A and lamin C generated
by alternative RNA splicing being the major somatic cell isoforms. Prelamin
A has 98 unique amino acids and lamin C 6 unique amino acids at their
carboxyl terminus (gray striping). LMNB1 on chromosome 5q23.3–q31.1
encodes lamin B1, and LMNB2 on chro- mosome 19p13.3 encodes lamin
B2, the somatic cell B-type lamins. All the lamins have conserved α-helical
rod domains and variable head and tail domains preceding and following
the central rod domain. The nuclear localization signals are located in the
tail domain (indicated in red). Prelamin A, lamin B1, and lamin B2 have
carboxyl-terminal CaaX motifs, a signal for protein farnesylation.
Worman et al. Laminopathies and the long strange trip from basic cell biology to therapy. J Clin Invest. 119:1825-1836,
2009.
Worman et al. Laminopathies and the long strange trip from basic
cell biology to therapy. J Clin Invest. 119:1825-1836, 2009.
Figure 2
Schematic diagram outlining the
posttranslational processing of nucle- ar
lamins. Prelamin A and B-type lamins
undergo 3 sequential post- translational
processing steps. First, the cysteine of the
carboxyl-ter- minal CaaX motif is
farnesylated by protein FTase. Second, the
–aaX is clipped off. For prelamin A, this is
likely a redundant activity of RCE1 and
ZMPSTE24. Third, the newly exposed
carboxyl-terminal farnesyl- cysteine is
methylated by isoprenylcysteine carboxyl
methyltransferase (ICMT). Prelamin A
undergoes another step in which the
carboxyl-ter- minal 15 amino acids,
including the farnesylcysteine methyl ester,
are clipped off by ZMPSTE24 and
degraded, generating mature lamin A. In
the setting of ZMPSTE24 deficiency, the
final endoproteolytic cleav- age does not
occur, leading to the accumulation of a
farnesylated and methylated prelamin A.
ZMPSTE24 deficiency causes a severe
progeroid disorder, RD. In HGPS, an
alternative splicing event results in a 50–
amino acid deletion in prelamin A,
removing the site for the final
endoproteolytic cleavage step. Thus,
mature lamin A cannot be produced, and
cells accumulate a mutant prelamin A that
terminates with a farnesylcysteine αmethyl ester.
Filamenti Intermedi - BUSB2011
Worman et al. Laminopathies and the long strange trip from basic cell biology to therapy. J Clin Invest. 119:1825-1836,
2009.
Laminopatie (1)
21
26/05/2013
Laminopatie (2)
Worman HJ, Fong LG, Muchir A, Young SG. Laminopathies and the long strange trip from basic cell biology to therapy. J Clin
Invest. 119:1825-1836, 2009.
Filamenti Intermedi - BUSB2011
22