NEUROSCIENZE NELLA STORIA:
Fino a pochi decenni fa non era scontato che il cervello fosse alla base delle
ns abilità cognitive. Nell’antichità
concezioni diverse:
-Ritrovamenti di teschi che contengono fori abbastanza precisi dà idea delle
domande che si ponevano gli antichi e dei loro tentativi di risolvere eventuali
problemi
-Egiziani: il cervello veniva estratto dai cadaveri e non conservato come
invece gli altri organi interni
-Per molto tempo il centro affettivo è stato ritenuto il cuore. Il cervello era
considerato come una ghiandola (Aristotele)
-Ippocrate: il cervello ha maggior potere sugli altri organi
-Nel cervello ci sono due cavità (ventricoli), interpretati dal medico romano
Galeno come contenitori di un liquido che veniva spinto in varie parti del
corpo
concezione ghiandolare.
-Idea di fluido che viene spinto mediante propulsione/sistema idraulico (L.da
Vinci)
Il cervello assume grande importanza ma il suo funzionamento reale non era
chiaro.
- Cartesio (1600): grande influenza sul pensiero successivo.
motorio/sensoriale (inferiore)
vedeva due livelli
anima (superiore)
mente
concezione dualistica
cervello
riteneva che la ghiandola pineale fosse il centro dell’attività cerebrale.
- Gall (fine ‘700): idea della FRENOLOGIA
il cervello è diviso in zone,
ognuna controlla una caratteristica particolare della personalità. In base a
protuberanze esterne sul cranio individua quale tratto è più sviluppato
(PRINCIPIO DELLA LOCALIZZAZIONE DELLA FUNZIONE)
vs
concezione vigente di unitarietà della persona (vd.Flourens)
-‘700: scoperta elettricità
usata su organismi viventi.
Il sist. nervoso è capace di reagire a elettricità proveniente dall’esterno.
1
Esperimenti di Galvani:
DELL’ELETTRICITA’
IL
SIST.
NERVOSO
SEGUE
LE
LEGGI
- Jackson (1850): descrizione di episodi epilettici riguardanti il sist. motorio
contrazione muscolare che può restare localizzata o estendersi in tutto il
corpo. Se le funzioni fossero distribuite nel cervello, l’attacco avrebbe
interessato tutto il corpo; invece procedeva x zone (epilessia jacksoniana)
- Broca: studio su un paziente incapace di parlare
le funzioni sono
localizzate. Il paziente era incapace di produzione linguistica, non di
comprensione. All’esame autoptico: danno nella parte frontale (area di Broca)
- Wernicke: studia afasia x quanto riguarda comprensione del linguaggio
lesione nel lobo temporale
trovate due aree correlate al linguaggio che, una volta lese, presentano
sindromi diverse, speculari.
Esperimenti di stimolazione elettrica: a seconda della zona stimolata,
all’esterno si hanno movimenti diversi.
Progressi ottenuti grazie all’introduzione di nuove tecniche.
- Brodmann (inizio ‘900): cervello suddiviso in aree diverse, numerate, sulla
base di esame istologico.
- VS Lashley: esperimenti su ratti con lesioni in diverse zone del cervello
concluse che non c’erano effetti, o se c’erano dipendevano dalla
grandezza e non dalla zona della lesione. Si sbagliava!!!!!!
- Anni ’70: visione dell’interno del cervello quando l’uomo svolge attività
motorie, quando è sottoposto a stimoli esterni, quando è impegnato in
attività cognitive. Sistemi di neuroimmagine: il sangue si sposta verso l’area
più attiva in un certo compito.
Una funzione è suddivisa tra un numero finito di aree collegate tra
loro a formare un circuito.
A seconda della funzione un’area è più attiva di un’altra.
No separazione netta tra funzioni dell’uomo e dell’animale.
I comportamenti simili possono derivare da OMOLOGIA (parti simili) o da
ANALOGIA (stessa tecnica, parti diverse)
2
EVOLUZIONE DEL CERVELLO:
Il nostro cervello è il risultato di un’evoluzione durata millenni di varie classi di
vertebrati. Diverse parti si sono evolute diversamente a seconda
dell’uso.
Uccelli/Mammiferi
più sviluppati il telencefalo e il cervelletto (connessi ad
abilità sensoriali/motorie)
Il cervello ha dimensioni diverse a seconda della specie.
Uomo: sempre considerato in cima alla scala evolutiva. Oggi si preferisce
parlare di “albero”evolutivo.
Ci si chiedeva se le abilità cognitive umane fossero legate al fatto che il
cervello umano è il più grosso
NO
Si tentò di rapportare il peso del cervello al peso corporeo
a parità di peso
corporeo, a seconda della classe di vertebrati presa in considerazione, il peso
del cervello è diverso. L’uomo ha l’indice di cefalizzazione (quindi
evoluzione cerebrale) maggiore.
Animali a sangue caldo: omeotermia (capacità di mantenere la temperatura
corporea)
il cervello può lavorare al meglio in ogni condizione di
temperatura
funzioni cerebrali più elevate
Correlazione tra STRUTTURE e DIMENSIONE del cervello.
LO STUDIO NEUROSCIENTIFICO:
Si è notevolmente differenziato negli ultimi 20/30 anni.
4 tipi di specializzazione:
- NEUROLOGO: diagnosticare/curare malattie del sist. nervoso
- PSICHIATRA: diagnosticare patologie più complesse (disturbi dell’umore
e della personalità)
- NEUROCHIRURGO: intervenire su cervello/midollo spinale in caso di
tumore, aneurisma…
- NEUROPATOLOGO: vedere cambiamenti di tessuto connessi a una
determinata malattia
3
ELETTRICITA’:
Tappa fondamentale x la fisiologia: scoperta di ELETTRICITA’ ANIMALE.
Le tecniche di misurazione di elettricità xmettono di misurare attività del sist.
nervoso. Si può usare elettricità sui sistemi biologici.
Galvani
exp: gambe di rana separate dal corpo, con ancora attaccato
nervo sciatico (coinvolto in contrazione muscoli x movimento della gamba).
E’ possibile applicare elettrodi x stimolare il nervo e far contrarre la zampa.
Ma dopo aver appoggiato un troncone sezionato di nervo sciatico al tronco
sano dell’altro e viceversa, le 2 zampe si contraggono senza applicazione
di elettricità dall’esterno. Tra la parte sezionata e quella sana, all’interno
del nervo si genera una differenza di potenziale.
Proprietà di ECCITABILITA’ ELETTRICA possedute dal sist. nervoso,
endocrino e muscolare.
Nervi: fibre nervose che conducono informazioni sottoforma di impulso
elettrico.
Utilizzati elettrodi x lo studio dell’attività del sist. nervoso (movimento di
particelle cariche)
Se questi due elettrodi sono posti su qcs di biologico
che ha al suo interno delle cariche, registro d.d.p.
(differenza di potenziale)
Se appoggio gli elettrodi su un cranio umano, registro l’attività elettrica sotto
lo scalpo.
Elettroencefalogramma: valutare se il comportamento cerebrale è normale,
se l’individuo è in stato di veglia, sonno o coma, se c’è attività a seguito di
diversi stimoli sensoriali (potenziale evocato: risultante di attività di tutti i
neuroni che si trovano sotto l’elettrodo)
La cute è rappresentata nel cervello in modo organizzato, come se fosse una
mappa.
STIMOLAZIONE SUPERFICIALE: gli elettrodi possono essere usati non solo x
registrare ma anche x applicare corrente elettrica (di bassa intensità) e
4
stimolare i neuroni che si trovano al di sotto. Si induce una variazione di
corrente all’interno del cervello, poi si vede cosa succede all’esterno (es.
movimenti che possono sembrare volontari)
Esperimenti con l’uomo direttamente sulla corteccia durante esp.
neurochirurgici
costruzione dell’ homunculus motorio
(rappresentazione sulla corteccia cerebrale dei ns movimenti)
Sviluppo della tecnica
microelettrodi : arrivano al diretto contatto con una
cellula nervosa. Utilizzati su animali, entrano nel tessuto nervoso. Rendono
possibile registrare l’attività elettrica del neurone.
Micropipette: sottilissimi capillari di vetro riempiti di liquido (conduttore)
collegato a un filo metallico (che fa da elettrodo)
sistema di conduzione.
Possibile collegamento con un misuratore di potenziale: la presenza di cariche
indica attività neuronale.
stimolazione sensoriale
Un neurone può essere studiato durante
attività cognitiva
I microelettrodi possono anche generare elettricità direttamente sul tessuto
nervoso
risoluzione molto più elevata
Non ci sono terminazioni dolorifiche all’interno del cervello.
E’ possibile correlare una determinata funzione con una determinata
popolazione di neuroni.
Se, a causa di una lesione (nell’uomo o nell’animale) o di un’ablazione (solo
nell’animale), tolgo un’area responsabile di una funzione, quest’ultima
scompare.
Inattivazione con determinati farmaci
viene temporaneamente bloccata
l’attività dei neuroni x vedere quale funzione viene compromessa.
NEURONI E DIFFERENZA DI POTENZIALE:
Neurone = cellula nervosa
Le cellule degli eucarioti hanno una membrana plasmatica. I neuroni sono
altamente eccitabili
l’eccitabilità a livello di membrana permette la
trasmissione di impulsi.
5
più
Potenziale all’interno del neurone: -65 mV
deve rimanere
o meno costante xchè la cellula sia eccitabile.
- +
Il liquido intracellulare ed extracellulare hanno molecole
- +
diversamente concentrate
- +
Voltmetro: registra d.d.p. tra l’esterno e l’interno della membrana
diversa
distribuzione di cariche, fondamentale xchè una cellula sia eccitabile
Il potenziale della membrana si calcola ponendo due elettrodi sulla superficie
esterna della membrana
E’ FALSO!!! Uno deve essere posto all’interno
Dendriti (x ricevere info)
Neurone: corpo cellulare + ramificazioni
Assoni
o
neuriti
(x
trasmettere
info)
La membrana cellulare è la stessa x ogni cellula
Doppio strato fosfolipidico
Immerse nella membrana cellulare ci sono le proteine transmembrana che
permettono il passaggio selettivo di determinate sostanze (es. ioni)
Liquido intracellulare = liquido extracellulare
Ioni più importanti:
(distribuiti diversamente)
cellula,
K+
Na+
Cl-
A (anioni organici presenti
all’interno della
sostanze
organica
di
natura
o biochimica ad es.
DNA)
più concentrati
all’esterno
più concentrati all’interno
La membrana del neurone è permeabile ad alcuni ioni. Esistono canali di
membrana specifici x certi ioni. Possono essere passivi (sempre aperti) o
attivi.
6
K+ passa liberamente attraverso la membrana del neurone, ma è più
concentrato all’interno.
Concentrazione = numero di molecole x unità di volume
Osmosi = movimento dell’acqua lungo il suo gradiente di concentrazione
All’equilibrio il flusso netto di una sostanza che diffonde attraverso la
membrana è nullo:
concentrazione interna del soluto + sostanze organiche interne =
concentrazione esterna del soluto
Per evitare un’osmosi eccessiva, la membrana è impermeabile solo a certi
soluti.
I soluti abbassano la concentrazione dell’acqua all’interno della cellula!
Le cellule animali trattengono gli ioni Na+ (sodio) nel liquido extracellulare,
per cui è più concentrato all’esterno.
Interno
Esterno
K+
K+
AA-
Esterno
Interno
Esterno
K+
K+
-+
K+
K+
- - ++
A-
A-
-+
A-
A-
- - ++
Gradiente di concentrazione
all’interno
Interno
che sospinge all’esterno
respingono)
Migrazione di ioni: cariche che creano corrente.
Gradiente elettrico
che risospinge
(le
cariche+
si
Acqua: molecola polare
può legarsi con ioni ± presenti nel fluido intra ed
extracellulare. Gli ioni più piccoli vengono avvolti da nuvole d’acqua
Volume reale: ioni + nuvola d’acqua.
La permeabilità della membrana allo ione K+ è maggiore della permeabilità
allo ione Na+
Il canale che permette il passaggio degli ioni non si comporta come un
semplice tunnel ma è dotato di cariche che spingono lo ione dall’altra parte.
Alcuni canali sono selettivi
7
attivi:
funzionano
solo in
particolari sit.
passivi:
come tunnel. Possono essere chiusi da una particella
che blocca il passaggio.
I canali possono essere aperti/chiusi in diversi modi.
PERIODO DI REFRATTARIETA’: i canali si chiudono e non possono
riaprirsi x un certo periodo di tempo.
I canali sono mediatori di determinate funzioni che non possono essere
esercitate continuamente, altrimenti ci sarebbero continue stimolazioni.
Gli agonisti endogeni sono sostanze già presenti nel corpo che naturalmente
fanno aprire/chiudere i canali. L’interazione può avvenire anche con sostanze
penetrate dall’esterno (esogene) che assomigliano alle sostanze presenti nel
corpo e causano effetti simili
molti farmaci sfruttano questo principio,
sostituiscono una molecola che non c’è più o ne sostituiscono l’effetto. Le
proteine dei canali hanno la possibilità di cambiare la loro conformazione
quando entrano in contatto con queste sostanze.
L’eccitabilità è data da quello che succede a livello della membrana.
Versante intracellulare: cariche –
Versante extracellulare: cariche +
Utilizzando micorelettrodi e micropipette si è potuta registrare la d.d.p.
Circa –60millivolt: d.d.p. della membrana della cellula nervosa in situazione
di riposo (potenziale di riposo)
- Liberando cariche negative all’interno della cellula si ha
IPERPOLARIZZAZIONE della membrana
aumenta la negatività.
- Liberando cariche positive all’interno della cellula si ha
DEPOLARIZZAZIONE della membrana
diminuisce la negatività.
una
una
Il passaggio degli ioni dipende dalla permeabilità della membrana. Uno ione è
più permeabile quando può passare di più rispetto a un altro ione.
Lo ione Na+ passa più difficilmente di K+ a causa delle molecole d’acqua che
lo circondano.
Le cellule gliali sono permeabili solo al K+ (membrana di potassio)
8
Le cellule nervose sono permeabili a K+, Na+, Cl-.
Quando il gradiente di concentrazione e il gradiente elettrico generano
due flussi uguali ed opposti si ha una situazione di equilibrio.
La d.d.p. è di –75 mV per le cellule gliali. (= al potenziale di equilibrio del
potassio)
In una cellula nervosa, lo ione Na+ è più concentrato all’esterno e il gradiente
di concentrazione lo spinge verso l’interno, lo stesso accade con il suo
gradiente elettrico.
Nella membrana a riposo (-60 mV) i canali K+ sono molto più aperti e
numerosi dei canali Na+
K+ è molto più permeabile di Na+, è una
forza più importante del determinare il potenziale della membrana.
Il K+ è soggetto a due forze e tende ad aumentare la sua concentrazione
esterna. Il Na+ tende ad aumentare la sua concentrazione interna.
La situazione tenderebbe a stabilizzarsi, entra in azione la POMPA SODIOPOTASSIO: attacca 2 ioni K+ da fuori e 3 ioni Na+ da dentro e li scambia in
maniera diseguale (pompa elettrogenica). Per funzionare utilizza l’ATP
(fenomeno attivo). Se gli ioni non fossero distribuiti in modo diseguale non ci
sarebbe la d.d.p.
RIASSUMENDO…
La membrana plasmatica è permeabile ad alcuni ioni grazie alla presenza, in
essa, di canali ionici
Proteine che attraversano la membrana delimitando un lume interno che fa
passare gli ioni. Possono essere passivi o attivi, aperti, chiusi o inattivati.
Ai capi della membrana plasmatici del neurone esiste una d.d.p. che, a
riposo, è di circa –60 mV, interno negativo, esterno positivo. Causa: ineguale
distribuzione di carche + e – che riguarda solo quelle parti di liquido extra ed
intracellulare vicino alla membrana.
La d.d.p. della membrana a riposo è dovuta a:
- diversa concentrazione interna ed esterna degli ioni potassio (K+),
sodio (Na+) , cloro (Cl-) e degli anioni organici (A-)
- diversa permeabilità dei vari ioni
9
- genesi di due tipi di gradiente (di concentrazione e elettrico).
- presenza della pompa elettrogenica (proteina che permette di
scambiare ioni K+ e Na+ in modo che le loro concentrazioni all’interno
e all’esterno siano disuguali. Manda fuori 3 Na+ e fa entrare 2 K+)
CIRCUITO ELETTRICO EQUIVALENTE:
La situazione ai capi della membrana può essere paragonata ad un circuito
elettrico equivalente.
Molti canali aperti
il flusso netto di ioni che passa è molto forte
Na+ è spinto dai due gradienti ad entrare all’interno della membrana, ma la
sua permeabilità è minore rispetto a quella del K+.
Il potenziale di membrana tiene conto della permeabilità e della
concentrazione degli ioni (equazione di Goldman)
La membrana plasmatica, a seguito di eventi esterni, può cambiare il suo
potenziale
potenziale d’azione (+55 mV) (=potenziale di equilibrio x
il Na+)
La misura reale del passaggio di ioni è espressa dalla conduttanza.
Conduttanza totale: conduttanza del singolo canale x numero dei canali
Il potenziale di membrana dipende dalla conduttanza e dal potenziale di
equilibrio di ogni ione.
Il liquido extracellulare e il citoplasma sono buoni conduttori
possono
essere assimilati a un corto circuito (conduttore a resistenza = 0)
Le pompe sono assimilabili a generatori di corrente.
I tre ioni sono assimilabili a batterie (potenziale di equilibrio x i vari ioni)
La membrana plasmatica è isolante. Ha una capacità elettrica (di accumulare
cariche), che entra in gioco quando si creano variazioni del potenziale di
membrana. Ha perdite di cariche attraverso i canali.
ANATOMIA DEL CERVELLO:
Studio neuroanatomico dal pt di vista macroscopico e microscopico.
Confronto: - cervello del ratto è liscio
10
- cervello di altri mammiferi (gatto, uomo…) presenta dei solchi
cranica
tenderebbe ad aumentare più del volume interno della scatola
SOLCHI: permettono l’aumento della superficie della corteccia cerebrale
mantenendo un volume limitato nella scatola cranica. Sono dei ripiegamenti
della corteccia cerebrale, risultato dell’evoluzione. Delimitano anche aree che
corrispondono a funzioni diverse.
Il solco centrale separa la corteccia motoria (davanti) dalla corteccia
sensitiva (dietro). E’ uguale nelle scimmie e nell’uomo.
I solchi possono anche separare rappresentazioni diverse del corpo.
Suddivisione:
- rostrale (anteriore): faccia
- dorsale (superiore): cima del capo
x parte superiore della testa
- ventrale (inferiore): sotto la testa
- ventrale (anteriore)
- dorsale (posteriore)
x parte inferiore della testa
- caudale (posteriore): nuca
Uomo: torsione di 90° dell’asse della testa. Nomenclatura diversa dagli
animali (che hanno struttura longitudinale)
Parte MEDIALE del cervello: ciò che è PIU’ VICINO ALL’ASSE CENTRALE.
Parte LATERALE: ciò che è PIU’ LONTANO DALL’ASSE CENTRALE.
Connessioni tra zone diverse del cervello sono note negli animali ma non
nell’uomo, su cui certi esperimenti non si possono fare.
MENINGI: lamine che avvolgono il cervello
- DURA MADRE: la più esterna
- ARACNOIDE: l’intermedia
Funzione di protezione
- PIA MADRE: aderente al cervello
Il liquido cefalorachidiano che scorre a livello delle meningi fa sì che il
cervello in pratica vi galleggi dentro
lo protegge dagli urti.
11
Il tessuto nervoso è molle
non facilmente studiabile
si usano sostanze
che pemettono l’indurimento del cervello (formalina). Procedimento di
fissazione: studio della struttura interna.
Cervello tagliato a metà: 2 sezioni. A occhio nudo si notano parti giallo chiaro
e altre giallo scuro: SOSTANZA GRIGIA e SOSTANZA BIANCA
Forte densità di cellule nervose
(corpi cellulari dei neuroni)
Fibre
Per studiare il cervello è necessario sezionarlo secondo determinati piani, per
ottenere fettine molto sottili (40-60 micron) x l’esame istologico:
1) Piano coronale: da dx verso sx
2) Piano sagittale: da davanti a dietro, attraverso la linea mediana della
testa
3) Piano orizzontale
1) ┴ 2)
3) ┴ 1) e 2)
1) parte dx e parte sx del cervello sono pressoché identiche
convenzione:
la sezione tagliata va guardata da dietro x stabilire qual è la dx e quale la sx.
2) convenzione: guardare il cervello di profilo dalla parte di sx. Alla nostra sx:
sezione anteriore. Alla nostra dx: sezione posteriore.
3) convenzione: guardare dall’alto x individuare parte dx e sx.
TAC: possibile vedere alcune porzioni del cervello. E’ una sezione virtuale del
cervello dell’individuo.
I tessuti possono essere sottoposti a colorazione x renderne visibile la
struttura interna. Le fettine vengono immerse in coloranti speciali; alla fine
vengono disidratate, chiarificate
possibile evidenziare le cellule sullo
spazio extracellulare.
Le cellule si possono aggregare dando vita a strutture.
Cellule nervose: corpo cellulare + prolungamenti ( non evidenziabili con tutti i
coloranti)
Metodo di Golgi (colorazione attraverso sali): si vedono il corpo dei neuroni e
i prolungamenti.
12
La cellula è rivestita dalla membrana plasmatica che ha la funzione di dare
costituzione e protezione alla cellula.
Impermeabile ma attraversata
da
Fuori dalla cellula: liquido ricco di ioni
proteine con funz. di canali
ionici
Nel tessuto nervoso ci sono cellule non nervose (GLIA) di due tipi:
- OLIGODENDROCITI: si trovano nel cervello
stessa
- CELLULE DI SCHWANN: si trovano nella periferia
funzione
Producono mielina (avvolgimento scuro attorno al prolungamento del
neurone). La guaina mielinica avvolge i filamenti dei neuroni.
Le cellule gliali mandano i loro prolungamenti a contattare i prolungamenti a
contattare i prolungamenti della cellula nervosa. Formano un avvolgimento
che diventa una guaina formata da membrana plasmatica (isolante), che
accelera la trasmissione dell’impulso nervoso.
MIELINA: serie di foglietti di membrana che si avvolgono ripetutamente
attorno alle fibre nervose, non prodotti dalle cellule nervose ma dalle cellule
gliali. In mezzo non c’è nessun liquido, viene spremuto in direzione del corpo
cellulare della cellula gliale.
La guaina mielinica è formata da strati di acidi grassi.
- no comunicazione tra cellula nervosa e liquido extracellulare
- trasmissione più rapida dell’impulso nervoso (grande velocità di
.
conduzione)
Non tutte le fibre nervose sono mieliniche!
La mielina può essere visualizzata attraverso procedure istologiche (colorante
che evidenzia le fibre mieliniche e non i corpi cellulari)
Se c’è demielinizzazione della fibra nervosa
zone bianche
La mielina normalmente si ricrea, a meno di una malattia demielinizzante (es.
sclerosi multipla)
La stessa zona cerebrale può essere studiata con 3 metodi diversi (tre tipi
di colorazione che evidenziano componenti diverse)
morfologia
Fibre
Corpi cellulari
Corpi cellulari +
disposizione
prolungamenti
LA CELLULA NERVOSA:
13
Secondo Golgi il sist. nervoso è formato da una rete fatta di fili
(prolungamenti) e bottoni (corpi cellulari) e le cellule sono in continuità tra di
loro tramite i prolungamenti. Si sbagliava!!!!!!!!!!
Secondo Cajal ogni cellula nervosa è unica e isolata, ma in contatto con le
altre cellule nervose attraverso una zona dove sono giustapposte (no
continuità citoplasmatica). Aveva ragione!!!!!!!!!!!!!
Le cellule nervose sono unità anatomiche.
ORGANIZZAZIONE:
corpo cellulare + prolungamenti
funzione di
integrazione
DENDRITI
(pochi o molti)
ASSONE (o neurite o fibra nervosa)
(unico)
elemento ricevente
msg.
elevato
elemento di emissione del
(permettono alla cellula
nervosa di ricevere i
msg esterni provenienti
da altri neuroni)
DENDRITI
BASALI
(si originano dal
corpo cellulare)
Trasporta un potenziale
(potenziale d’azione o
impulso nervoso)
DENDRITI
APICALI
(si originano da
un dendrite più grande)
Possono essere portatori di variazioni di potenziale di
membrana dell’ordine di pochi millivolt.
Neurone: polarizzazione dinamica (il messaggio va in un’unica direzione)
DENDRITI
CORPO CELLULARE
ASSONE
Altri neuroni si mettono in contatto col neurone attraverso una sorta di
“bottoncini” che arrivano in contiguità col neurone (SINAPSI)
inibitorie
eccitatorie
14
depolarizzazione
iperpolarizzazione
Cono d’emergenza: punto in cui l’assone emerge dal corpo cellulare. QUI
nasce l’impulso nervoso.
Fibra nervosa:
la mielina è interrotta in alcuni punti
(NODI DI RANVIER)
L’assone ha una ramificazione finale x entrare in contatto con diverse parti
del dendrite o con diversi neuroni.
Principio della CONVERGENZA: su un unico neurone convergono le
terminazioni di più neuroni
Principio della DIVERGENZA: un unico assone, grazie allo sfioccamento,
riesce ad influenzare più neuroni.
SINAPSI: (è dove le cellule nervose comunicano tra loro)
Pt. di contatto tra il bottone finale dell’assone e il corpo/dendrite del neurone
sinaptico
Elemento pre-sinaptico
Elemento post-
DOTTRINA DEL NURONE: il neurone è un’unità
- ANATOMICA, isolata dagli altri ma c’è contiguità tra di loro (x
comunicazione)
- GENETICA, sa già nell’embrione che posto deve occupare all’interno
del tessuto nervoso, verso dove andrà a migrare, con quali cellule
dovrà avere contatto e quale tipo di informazione dovrà utilizzare con
altre cellule
- FUNZIONALE, ogni neurone ha la sua funzione (popolazione =
insieme di neuroni con uguale funzione)
- TROFICA, ogni neurone è caratterizzato da sostanze che gli
permettono di crescere (NGF). I neuroni devono essere sostenuti da
fattori di crescita. In particolari situazioni (traumi), entro certi limiti,
possono rigenerarsi.
15
Ai dendriti apicali/basali arrivano numerosi segnali (eccitatori e inibitori) da
altri neuroni. Questi segnali vengono trasformati in potenziali graduati, che si
sommano dal pt di vista temporale e spaziale, e poi integrati nel corpo
cellulare.
Il POTENZIALE D’AZIONE (risposta del neurone) nasce nel
cono d’emergenza dell’assone (dove ci sono più canali voltaggio-dipendenti).
Si propaga in una sola direzione (ortodromica: dal corpo cellulare alle
terminazioni, e NON antidromica!) con una conduzione di tipo saltatorio.
L’assone si ramifica nella sua parte terminale (divergenza)
il messaggio
raggiunge più neuroni.
N.B. Nei dendriti NON nasce il potenziale d’azione!!!!!!!!!!!!!!!!!!!!!!!!!
A seconda del neurone, la frequenza di scarica dei suoi potenziali d’azione
può variare.
I neuroni sono di vari generi:
- neuroni bipolari
- neuroni pseudounipolari
- neuroni multipolari (ampia arborizzazione dendritica)
I motoneuroni sono nel midollo spinale e servono x la contrazione muscolare.
Anatomia
Funzione
PROPRIETA’ ELETTRICHE PASSIVE DEL NEURONE:
Cambiamento del potenziale di membrana
inizio di possibilità di risposta
da parte del neurone.
Il potenziale di membrana può essere cambiato
Potenziale d’azione (impulso nervoso) è un fenomeno attivo!
Variazione del potenziale di membrana tramite introduzione di corrente:
- corrente capacitiva (carica la capacità di membrana): è trasportata
dagli ioni che si caricano sulle due facce della membrana.
- corrente ionica o resistiva (passa attraverso la membrana): è
trasportata dagli ioni che passano attraverso la membrana.
Risposta della membrana plasmatica
16
all’introduzione della corrente dall’esterno
Simulazione esterna
variazione del potenziale elettrico
COSTANTE DI TEMPO (τ) = il tempo impiegato a raggiungere il 63 % del
valore del potenziale finale.
Quando la membrana è stimolata, il processo di variazione di potenziale
impiega del tempo, maggiore o minore a seconda del tipo di fibra nervosa
Possibile generazione di una depolarizzazione/ iperpolarizzazione
successiva se la prima depolarizzazione/iperpolarizzazione dura nel tempo. Se
sono vicine, si possono sommare.
La membrana plasmatica ha delle resistenze interne (al passaggio delle
cariche) dovute al fluido intracellulare
Resistenza di membrana: somma delle resistenze dei canali ionici. A parità
di corrente è sempre la stessa.
La resistenza citoplasmatica aumenta all’aumentare della distanza.
COSTANTE DI SPAZIO(λ) : la distanza alla quale il potenziale è sceso al
37% del potenziale di membrana all’origine
Posso sapere quanto la
variazione di potenziale potrà propagarsi nello spazio lungo la fibra nervosa.
λ=
seguire il
r di membrana
r assiale
se è alta, le cariche la attraversano con
difficoltà. Le cariche tendono a
citoplasma e a non uscire attraverso i
canali
maggiore λ
maggiore τ
la depolarizzazione si propagherà nello spazio
aumenta la probabilità che una depolarizzazione
successiva si sommi alla precedente
La variazione di potenziale è passiva, soggetta a ritornare alla sua condizione
iniziale, ma può originare un impulso nervoso.
Τ = R x C (resistenza di membrana x capacità)
Fibre nervose molto piccole
Fibre nervose grandi
bassa velocità di conduzione impulso
alta velocità
“
“
“
17
CONDUZIONE ELETTROTONICA: variazione di potenziale lungo la fibra
nervosa dovuta a meccanismi passivi (flusso di cariche attraverso canali ionici
passivi)
POTENZIALE D’AZIONE:
Corrente Na+ entrante
Na+
³
depolarizzazione
²
¹
apertura dei canali
(voltaggio-
dipendenti)
I canali al Na+ voltaggio-dipendenti possono essere aperti da una variazione
di voltaggio della membrana. NON SONO PASSIVI!
Na+ tendono ad entrare x gradiente di concentrazione e x gradiente elettrico.
La membrana inverte il suo potenziale. Tende ad arrivare al potenziale
di equilibrio del sodio (+55 mV), non ci riesce xchè viene contrastato.
VISUALIZZAZIONE DEI COLLEGAMENTI:
Forte trauma
degenerazione della fibra nervosa. Il neurone legato
all’assone interrotto può andare incontro a morte. In altri casi, può
riprendersi e rigenerare la parte di assone degenerata.
Le fibre in degenerazione sono colorabili in argento, ma non si vedono più
con la colorazione x la mielina. Questi tipi di colorazione però non mostrano
tutti i tipi di collegamento. Ci sono quindi altri metodi.
TRASPORTO ASSONALE:
Altri sistemi permettono di studiare i collegamenti anche quando le fibre sono
intatte. All’interno della fibra nervosa ci sono dei meccanismi che assicurano il
trasporto di sostanze e organelli, in tubuli, all’interno dell’assoplasma:
- flusso assoplasmatico lento (no tubuli, x componenti strutturali)
- trasporti rapidi, sia in direzione anterograda (dal corpo cellulare
alla periferia: es. neurotrasmettitore portato dal corpo cellulare alle
terminazioni in vescicole) che retrograda (dalla periferia al corpo
cellulare: es. trasporto di materiali di degradazione al corpo cellulare
che li riutilizza, o di NGF, o di elementi negativi come virus o tossine)
18
Le particelle trasportate in direzione retrograda vengono dalle terminazioni o
dal liquido extracellulare.
Tracciante retroanterogrado: posso vedere se due strutture sono
collegate in maniera reciproca.
1
trasporto
Inietto la sostanza in A. Grazie al
attivo
trasportato
anterogrado
,
esso
viene
nella terminazione 1 a partire dal
corpo
A
B
cellulare del neurone.
La sostanza viene presa anche
dalla
2
terminazione del neurone e trasportata x
trasporto attivo retrogrado nel corpo
cellulare 2.
Accumulo sostanza in B.
La sostanza iniettata deve essere resa visibile xchè io possa vedere il
collegamento tra A e B. Il collegamento tra le due strutture viaggia nei due
sensi.
Riesco a riconoscere i corpi cellulari dalle terminazioni. Ho un livello di
risoluzione più alto rispetto alla degenerazione.
Queste sostanze danno una marcatura. Ci sono tempi diversi a seconda delle
sostanze che vengono utilizzate.
I sistemi a trasporto attivo permettono di studiare ad alta
risoluzione le connessioni tra le varie strutture del sist. nervoso.
STRUTTURA DEL SISTEMA NERVOSO:
Suddivisioni generali:
SISTEMA NERVOSO CENTRALE
SISTEMA NERVOSO PERIFERICO
Midollo spinale (in colonna vertebrale)
Tronco dell’encefalo + cervelletto
muscoli
+ proencefalo (diencefalo e telencefalo)
fibre che collegano il SNC
con la periferia (cute,
e articolazioni) in 2 sensi
19
(afferenti ed efferenti)
SISTEMA NERVOSO SOMATICO
Agisce attraverso
muscolatura scheletrica
SISTEMA NERVOSO AUTONOMO
controllo su organi interni
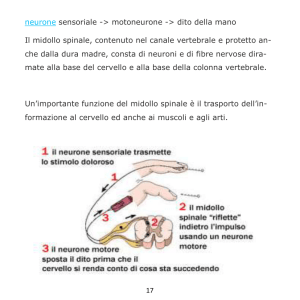
MIDOLLO SPINALE: riceve info sensoriali dalle periferia, determina la
contrazione dei muscoli che muovono le articolazioni (funzione sensoriale e
motoria) + integrazione di risposte sensoriali e motorie. Ad esso arrivano
fibre ascendenti/discendenti. E’ sede principale dei riflessi e dei meccanismi
che permettono la locomozione. E’ separato dalle strutture sovrastanti
TRONCO DELL’ENCEFALO (bulbo, ponte, mesencefalo): è primitivo ma
sede di funzioni importanti: controllo equilibrio, locomozione, dolore. Sede di
info sensoriali provenienti dalla testa (vista, gusto, udito), sede di centri che
regolano attività vegetative (respirazione, battito cardiaco…) e di zone che
xmettono di cambiare stati di coscienza (veglia/sonno)
CERVELLETTO: riceve info sensoriali, manda comandi motori. E’ un sistema
di controllo motorio + sede di meccanismi di apprendimento legati al
movimento.
CERVELLO= DIENCEFALO(TALAMO + IPOTALAMO) + TELENCEFALO
(CORTECCIA CEREBRALE + GANGLI DELLA BASE):
TALAMO: punto di passaggio di info che vanno verso la corteccia cerebrale.
Influenza l’eccitabilità generale della corteccia e gli stati generali di coscienza.
IPOTALAMO: coordina attività somatiche, vegetative e endocrine. E’ centro
di mantenimento degli equilibri (termico, dei liquidi…) interni al corpo.
Controlla l’ipofisi (ghiandola centrale dell’organismo).
CORTECCIA CEREBRALE: materia grigia dai 2 ai 4 mm con sotto sostanza
bianca. Riceve info soprattutto dal talamo e manda le sue uscite verso il
basso. Sede di percezioni sensoriali, dei programmi motori, di funzioni
cognitive (linguaggio, memoria, apprendimento) e emotività. Ha rapporti con
strutture sottocorticali e le varie aree della corteccia stessa, che
rappresentano diverse funzioni, hanno rapporti tra loro.
GANGLI DELLA BASE: struttura originariamente di tipo motorio.
PRINCIPI GENERALI DEL SISTEMA NERVOSO:
20
- La comunicazione nel sistema nervoso avviene attraverso la SINAPSI
(contatto tra terminazione assonale e neurone, usato anche tra cellule
nervose e effettori es. muscoli)
- Sistema di rappresentazione topografica: analisi o azione sul mondo
interno presuppone mappatura del corpo in varie strutture del sist.
nervoso (omuncolo: mappa corporea sulla corteccia cerebrale)
- Controllo nervoso sensoriale/motorio è di tipo crociato: la corteccia di
sx controlla la parte dx del corpo e viceversa.
- Suddivisione in modalità e sottomodalità (es. vista: analisi colore,
forma…)
SINAPSI:
Le cellule nervose comunicano tra loro tramite la sinapsi
membrane delle cellule.
vicinanza tra le
L’impulso nervoso arriva alla terminazione di un neurone (presinaptico)
Modificazione della membrana del neurone postsinaptico.
Sinapsi assosomatica: termina sul soma della cellula postsinaptica
“
Assodendritica: termina sul dendrite della cellula postsinaptica
“
Assoassonica: termina sull’assone “
“
“
L’impulso nervoso viaggia lungo l’assone e a livello della terminazione accade
qcs.
SINAPSI
ELETTRICHE
CHIMICHE
Sinapsi elettriche: le membrane sono molto vicine tra loro, l’elemento pre
e post sinaptico sono separati da uno spazio molto piccolo. Canali aperti a
contatto tra loro xmettono il passaggio di liquido plasmatico e di ioni tra le
due cellule
GIUNZIONI COMUNICANTI o GAP JUNCTION: una parte di
corrente passa dalla cellula pre alla cellula post sinaptica (flussi di corrente
bidirezionali) Nella sinapsi chimica invece avviene in una sola direzione!!!!
Le sinapsi elettriche hanno una velocità di trasmissione molto alta
molto
importante nelle reazioni di difesa. Spesso interconnettono intere popolazioni
di neuroni.
21
Sinapsi chimiche: es. SINAPSI NEUROMUSCOLARE.
Motoneurone: innerva i muscoli, xmette la contrazione.
assone
placca motrice
Viene rilasciato il NEUROTRASMETTITORE (es. acetilcolina x la sinapsi
neuromuscolare) nello spazio intersinaptico.
Recettori: proteine di membrana postsinaptica a cui si legherà il
neurotrasmettitore.
Vescicole sinaptiche: contengono il neurotrasmettitore che viene rilasciato.
Per esocitosi si fonderanno con la membrana postsinaptica e libereranno il n.
Cosa accade?
Arrivo del potenziale d’azione
aumenta la permeabilità della membrana
canali voltaggio-dipendenti x il calcio si aprono facendo entrare ioni Ca
segnale che xmette fusione di vescicole sinaptiche con la membrana e rilascio
del neurotrasmettitore
il n. si va a legare col recettore x l’acetilcolina (è
un canale normalmente chiuso. Si apre quando due molecole di acetilcolina si
legano su due siti specifici)
Attraverso lo stesso canale, entra Na+ ed esce K+
depolarizzazione nella
fibra muscolare
POTENZIALE DI PLACCA
Più acetilcolina viene liberata, più canali si aprono e maggiore sarà l’ampiezza
del potenziale.
ricaptata da membrana pre
dopo va disattivata
degradata
da
un
enzima
(acetilcolinesterasi)
si propaga in maniera elettrotonica: perde di intensità allontanandosi dalla
zona di stimolazione. Si propaga nelle zone adiacenti alla placca motrice,
dove esistono canali voltaggio-dipendenti per il Na che si aprono nasce un
potenziale d’azione nel muscolo, che si contrae.
SANGUE E CERVELLO:
Ci sono dei liquidi che bagnano il cervello. Riceve sangue. Il cervello, rispetto
al peso dell’uomo, è piccolo (2%) ma è molto rifornito di sangue (15% di
quello pompato dal cuore in un minuto) e di ossigeno
Circa il 20% di quello che entra
Porta al cervello ossigeno,
22
nei polmoni è inviato al cervello.
Utilizzato x respirazione cellulare
(mitocondri: possono formare ATP,
respiraz.
Molecola carica di energia)
chimiche,
glucosio e altre sostanze
nutritive. Porta via l’anidride
carbonica prodotta da
cellulare e le sost. di rifiuto
prodotte
da
reazioni
attraverso
venoso.
il
ritorno
Esiste un sistema arterioso che irrora tutto il SNC.
Il cervello può essere rifornito anche se in altre parti c’è un blocco, grazie a
un sistema particolare. Le arterie irrorano territori diversi del cervello. Se
avviene un danno a una di queste arterie, i territori cerebrali (indipendenti da
suddivisione funzionale) irrorati da essa vanno incontro a seri problemi.
Se avvengono danni circolatori
arrivano più
ICTUS:
componenti portate dal sangue non
- ISCHEMIA: chiusura di un vaso per 1) indurimento del vaso causato
dall’età 2)formazione di un trombo (grumo) che blocca la
circolazione in quel punto 3) compressione (tumore). Le cellule non
più irrorate muoiono
infarto (necrosi di una popolazione
cellulare)
- EMORRAGIA: 1)aumento di pressione che tende a rompere il vaso
2) aneurisma: pareti “erniate”, deboli. Se l’aneurisma è in posizione
operabile si può togliere, altrimenti si deve controllare. Crea un
versamento di sangue nella zona di rottura. Può essere riassorbito,
ma questo sangue può premere su vasi vicini e creare successiva
ischemia.
Il cervello è estremamente sensibile a diminuzioni di glucosio e di ossigeno.
Sistema venoso: vene profonde + vene superficiali
più grosse che confluiscono nella vena giugulare.
confluiscono in vene
LIQUIDO CEFALORACHIDIANO (o cerebrospinale):
Contenuto in cisterne all’interno dell’encefalo. Durante lo sviluppo embrionale
si evolvono
formazione dei VENTRICOLI
23
Liquido prodotto da strutture specializzate nel 3° ventricolo (nel diencefalo)
chiamate plessi corioidei. Il liquido viene mandato ai ventricoli laterali, poi
attraverso l’acquedotto (nel tronco dell’encefalo) e a livello del 4° ventricolo
(dietro il cervelletto). Si distribuisce all’esterno del SNC uscendo attraverso i
forami e scorre nello spazio sub-aracnoideo (al di sotto delle meningi).
Sbocchi: villi aracnoidei, fuoriuscite attraverso cui questo liquido può
essere convogliato nelle vene
la differenza tra secrezione del liquido e suo
assorbimento è uguale a zero.
Funzioni:
- Mantenere l’equilibrio tra il sangue e il liquido che bagna le
cellule. Liquido extracellulare
l. cerebrospinale
sangue
- Fare da barriera x il passaggio di alcune sostanze. Certe
sostanze possono passare tra il sangue e il cervello, altre no.
(BARRIERA EMATOENCEFALICA tra il sangue e il cervello)
- Attutire gli urti
BARRIERA EMATOLIQUORALE: tra il sangue e il liquido cerebrospinale
Sangue
liquido extracellulare che circonda i neuroni
(il liquido cerebrospinale fa da tramite)
sangue
Eccessiva secrezione /ridotto assorbimento di questo liquido
problemi: es.
idrocefalo: nel bambino c’è continua secrezione ma c’è un blocco a livello
dell’acquedotto
la testa si ingrossa.
Questo liquido dà info su possibili malattie del SN. Si può vedere facendo
asportazioni con un ago a livello lombare prelevando liquido che viene
esaminato (colore e concentrazione di sostanze
si deduce se c’è infezione)
MIDOLLO SPINALE:
24
Il midollo spinale è inserito nella colonna vertebrale. Suddivisione
corrispondente al num. delle vertebre. Da esso nascono prolungamenti che
escono dagli spazi tra le vertebre (nervi spinali). Ad ogni nervo è assegnato
un numero in relazione alla vertebra da cui esce.
Rigonfiamenti nel cordone
controllo nervoso più forte:
- cervicale
arti superiori
- lombare
arti inferiori
Zona interna del midollo: SOSTANZA GRIGIA (cellule nervose)
Zona esterna “
“
: SOSTANZA BIANCA (fibre)
E’ simmetrico, la sostanza grigia presenta le corna ventrali (o anteriori) e
dorsali (o posteriori). La sostanza grigia di dx e collegata con quella di sx.
La sostanza bianca è divisa in tre colonne (ventrale, dorsale, laterale) con
fibre diverse.
I nervi spinali sono il risultato dell’unione tra una radice dorsale e una radice
ventrale.
MIELOMERI: moduli del midollo forniti di 2 radici dorsali e 2 ventrali (unità
funzionale). Si formano durante l’embriogenesi. Ad ogni mielomero
corrisponde sulla cute un dermatomero, che fornisce l’innervazione al
corrispondente mielomero.
I DERMATOMERI si sovrappongono (regioni comuni)
se c’è un processo
infiammatorio di una radice dorsale che controlla una superficie cutanea,
questa può non perdere completamente il collegamento col midollo.
dolorifici
Dermatomeri
meno sovrapposti di
tattili
La lesione di una radice compromette un solo dermatomero, la lesione di un
nervo periferico comporta la perdita di più dermatomeri.
Possibilità di mappare la superficie corporea sul sistema nervoso.
Il midollo spinale è stato studiato con metodi istologici che rivelavano la
presenza di cellule e fibre, diverse come dimensione e come funzione
(citoarchitettonica: distingue le cellule)
25
Ci sono 3 tipi di neuroni disposti in
Corno dorsale
Zona intermedia
Corno ventrale
All’interno di ogni zona sono uguali. Ci sono delle sottozone.
Al midollo spinale arrivano fibre afferenti periferiche dalla cute, dai muscoli e
dalle articolazioni. Terminano in diverse zone della sostanza grigia facendo
sinapsi con i vari neuroni.
La classificazione di Rexed suddivide la sostanza grigia in lamine. Un’altra
classificazione la suddivide in settori.
Corno dorsale: lamine 1-5
volta divisa in sottozone.
zona sensoriale della sost. grigia, a sua
Zona intermedia: lamine 6-7
- nucleo di Clarke, nella porzione più mediale
- nucleo intermedio-laterale, nella zona toracica, sede dei neuroni del
sistema nervoso autonomo
- ci sono neuroni con caratteristiche integrative (interneuroni)
Corno ventrale: lamine 8-9
motoneuroni da cui partono fibre efferenti
(controllo del sist somatico). Ci sono anche interneuroni direttamente
collegati ai motoneuroni.
Il midollo spinale è una struttura molto primitiva. E’ un centro di controllo e
un punto di passaggio di informazioni.
Le info afferenti dalla periferia possono essere usate x:
1. semplici riflessi che si esauriscono a livello del midollo spinale (funzione
non cosciente)
2. ascendere fino all’encefalo e alla corteccia cerebrale (esperienza
cosciente della sensazione
percezione)
LA PERCEZIONE:
C’è una relazione tra lo stimolo esterno e la nostra percezione di quello
stimolo.
26
Due scuole di pensiero:
1. Scuola empirista: influenza di Comte, legata al Positivismo. Si può
studiare il comportamento umano (realtà misurabile) tramite leggi
usate x lo studio dei fenomeni fisici.
Cfr. esperienza precedente degli empiristi inglesi: le ns percezioni sono
direttamente misurate dalla ns esperienza sensoriale, il nostro cervello
è una somma di sensazioni. Modalità riduzionista.
Questa corrente di pensiero ha permesso la partenza della psicologia
sperimentale (studi psicofisici che mettono direttamente in relazione lo
stimolo con la percezione dell’individuo)
Ma il nostro cervello è una somma di sensazioni???
Molte percezioni sono costruzioni del ns cervello, sono tali xchè esistiamo noi.
Es. nel mondo fisico esistono onde, noi le percepiamo come luce o suono.
Mondo fisico misurabile
=
Mondo fisico percepito
Il nostro cervello è un organizzatore.
2. Scuola kantiana: nel cervello abbiamo categorie innate in cui
organizziamo le ns esperienze. Ha originato scuole che hanno
affrontato il rapporto stimolo/percezione in modo diverso dagli
empiristi, es. Gestalt (
il mondo esterno è un’organizzazione del
nostro cervello)
La stessa realtà fisica, cambiata rispetto al contesto, può dare origini a
percezioni diverse.
PSICOFISICA DELLA PERCEZIONE:
Lo stimolo deve essere considerato in relazione a:
1. modalità
2. intensità
3. durata
4. localizzazione
1. Lo stimolo esterno interagisce con qcs che si trova sulla superficie del
ns corpo. Noi percepiamo diverse modalità sensoriali (percezione
luminosa, acustica, etc)
27
Legge di Muller: lo stimolo interagisce con un recettore specifico, dal
quale dipende la ns sensazione. Il recettore deve essere colpito da uno
stimolo adeguato
la modalità della ns percezione non dipende
dal recettore, ma dalla parte del sistema nervoso attivata dopo il
recettore
Tramite tra la stimolazione esterna e il SN.
E’il SN che decide che tipo di percezione avrò.
2. Si può mettere in relazione uno stimolo esterno che ha una certa
energia con la percezione interna.
SOGLIA di percezione dello stimolo: 50% dei casi in cui lo stimolo
viene applicato ed esso dà origine ad una sensazione nel soggetto (a
volte lo stimolo è percepito, a volte no)
Può essere influenzata da:
- Aspettativa dello stimolo: se mi aspetto di ricevere uno stimolo,
la soglia di percezione dello stimolo si abbassa
- In altri casi la soglia di percezione si abbassa (es. fatica, uso di
farmaci…)
Modelli matematici:
- LEGGE DI WEBER: ΔS = KS
La variazione di intensità dello stimolo che mi produce variazione di
percezione è direttamente proporzionale all’intensità originaria. Se lo
stimolo iniziale è elevato, ci vuole un’elevata variazione xchè essa
venga percepita. Se lo stimolo iniziale è basso, verrà percepita una
bassa variazione dello stimolo.
- LEGGE DI FECHNER: I= k log S/S
(logaritmica)
- LEGGE DI STEVENS: I= k (S- S )ⁿ
(esponenziale)
L’intensità della percezione è correlata alla funzione esponenziale della
differenza dei due stimoli
Esistono relazioni tra intensità di stimolazione/percezione e sua
variazione.
3. Alcuni stimoli vengono applicati x breve tempo, altri costantemente. Ci
sono stimoli che è importante percepire immediatamente (es. stimoli
dolorifici), di cui è importante percepire l’inizio ma anche quanto
durano.
28
ABITUDINE: diminuisce la percezione dello stimolo anche se questo
continua ad essere applicato
4. E’ importante sapere dove uno stimolo viene applicato. Ci sono zone del
ns sistema recettoriale più o meno sensibili a un certo tipo di stimolo. E’
importante quanto si è in grado di discriminare, in senso spaziale, tra due
stimoli applicati contemporaneamente. L’ acuità discriminativa varia a
seconda della zona corporea. La distanza da tenere tra 2 stimoli xchè
vengano percepiti come distinti dipende dalla zona, cioè dalla densità di
innervazione, che è maggiore nelle porzioni distali (mani piedi bocca)del
corpo rispetto alle porzioni prossimali
Massima acuità: mani, piedi,
bocca, lingua.
PORZIONI DISTALI
MAGGIORE ACUITA’
La percezione è frutto della nostra elaborazione cerebrale!!!!!!!!!!!
I RECETTORI DEL SISTEMA NERVOSO:
Recettori del sistema sensoriale =
Recettori dei neurotrasmettitori !!!!
- Classificati in base a modalità (vd. Tabella pag. 408 Kandel nuova edizione)
Ci sono recettori che sono azionati da stimoli diversi (es. meccanocettori:
recettori sensibili a uno spostamento meccanico)
- Classificati in base al loro rapporto con la fibra sensitiva (pag 127 Rizzolatti)
I tipo: terminazioni periferiche di fibre sensitive afferenti (recettore
e fibra sensitiva in quel punto coincidono) es. Cellula del Pacini
II tipo: il recettore è una cellula modificata in contatto con la fibra
sensitiva afferente. Es. cellula del sistema dell’equilibrio
III tipo: (solo nella retina) in contatto con un neurone chiamato
cellula bipolare che è a sua volta a contatto con un altro neurone
(cellula gangliare) che è la fibra sensitiva afferente
Fibra sensitiva afferente
interneurone
recettore
TRASDUZIONE:
Come fa uno stimolo esterno a diventare info utilizzabile dal SNC?
29
Energie diverse che devono tradursi in un msg nervoso (di tipo
elettrico)
STIMOLO
VARIAZIONE DELLA
CONDUTTANZA DI
MEMBRANA
POTENZIALE
POTENZIALE
GENERATORE
D’AZIONE
Energia dello stimolo determina modificazione del recettore periferico
passaggio di ioni attraverso canali ionici che si aprono (depolarizzazione o
iperpolarizzazione)
Stimolo meccanico
stiramento della fibra periferica
apertura di
canali ionici
entrano ioni Na+
depolarizzazione a livello delle
terminazioni periferiche (potenziale generatore
si trasferisce in zona
vicina, nel primo nodo di Ranvier, dove ci sono canali voltaggio-dipendenti)
potenziale d’azione si trasmette x via aleatoria fino al midollo spinale
Se blocco meccanicamente il primo nodo di Ranvier, ho il potenziale
generatore ma non il potenziale d’azione
Nasce nella
Terminazione
è necessaria la
fibra sensitiva
Cellula del Pacini: ha una struttura accessoria
trasforma un recettore
a lento adattamento in un recettore a rapido adattamento. Se è presente, ho
un pot. generatore sia quando viene applicato lo stiramento sia quando viene
tolto.
Da usare se voglio conoscere la durata dello stimolo
Recettori I tipo: il potenziale generatore si trasmette in modo passivo x far
nascere il potenziale d’azione se si supera la soglia.
Recettori II tipo: il potenziale di recettore induce la liberazione di
neurotrasmettitore alla fibra afferente primaria, dove a livello del primo nodo
di Ranvier può nascere il pot. d’azione.
SISTEMA SOMATOSENSORIALE:
Meccanocettori: trasducono energia meccanica in energia elettrica
30
Sulla cute: reagiscono a pressione (leggera o profonda, singola o a
ripetizione, su cute pelosa o glabra…)
Parte superficiale e profonda
meccanocettori che segnalano vari tipi
di stimoli (recettori a contatto con bulbi piliferi, o con la parte +
esterna, o con la parte + profonda)
X stimolare questi ci deve essere una pressione + forte
- Terminazioni periferiche libere della cute
- Terminazioni che hanno contatto con cellule della cute (disco di
Merkel)
- Struttura a grappolo (corpuscolo di Meissner)
- Capsula che avvolge terminazioni libere (corpuscolo del Pacini)
- Terminazioni libere avvolte attorno al pelo
Struttura accessoria
modifica le proprietà del recettore
Corpuscolo di Meissner e di Pacini: rapido adattamento (sensibili a
stimolazioni ripetute/vibratorie)
Recettori di Merkel e corpuscoli di Ruffini : lento adattamento (sensibili a
infossamento continuo della cute)
Propriocettori: meccanocettori che portano info sullo stato dei muscoli e
delle articolazioni (statici o dinamici) es. fuso neuromuscolare (all’interno dei
muscoli) è un recettore complesso che contiene terminazioni afferenti di fibre
che porteranno info al midollo spinale
Nocicettori: terminazioni libere nella cute azionati da diversi stimoli
(meccanici, chimici, termici)
Termocettori: terminazioni libere nella cute
Per il freddo: fibre eccitate quando c’è uno stimolo a bassa
temperatura. Iniziano a rispondere sotto i 32 °C e aumentano la loro
risposta man mano che la temperatura diminuisce
la risposta del
recettore va in parallelo con la percezione del freddo. Alcuni di questi
recettori rispondono anche a stimolazioni particolarmente calde: oltre i
45 °C, quando i recettori x il caldo non rispondono più, iniziano a
rispondere quelli x il freddo
FREDDO PARADOSSO
Per il caldo: capaci di rispondere al riscaldamento tra i 34 e i 40 °C.
poi entrano in funzione i nocicettori termici (che rispondono al
riscaldamento eccessivo)
31
- Un recettore che porta una determinata modalità, può dare caratteristiche
abbastanza precise riguardo ad essa. ( Es. fibra acustica può rispondere a
delle frequenze. L’intervallo di frequenze a cui noi siamo sensibili dipende
dalle fibre acustiche. Ogni fibra acustica risp in un certo intervallo di
frequenza. La risp non è uguale x tutte le frequenze). Recettore + fibra
possono risp in un intervallo di energia, ma all’interno di esso l’intensità di
stimolazione che devo usare x avere la stessa risposta è diversa (ampiezza
dello stimolo). Per ogni recettore ho valori ottimali x frequenze anche molto
piccole.
- Intensità dello stimolo : corrisponde alla frequenza di scarica delle fibre
nervose (proporzionalità diretta)
Se si aumenta l’intensità dello stimolo, aumenta la frequenza di scarica:
STIMOLO INTENSO
ALTA FREQUENZA DI SCARICA
STIMOLO POCO INTENSO
BASSA FREQUENZA DI SCARICA
Se la frequenza di scarica è elevata, lo stimolo è ottimale.
- Durata: dipende da proprietà di adattamento dei recettori e delle fibre di
cui fanno parte
- Lento adattamento
frequenza di scarica ripetuta
- Rapido adattamento
scarica a inizio e fine stimolazione (risp a
stimoli di breve durata)
L’adattamento della fibra nervosa ci dà percezione della durata dello stimolo.
- Localizzazione: ogni neurone ha il suo campo recettivo
zona di cute che, stimolata, determina
variazione di scarica nel neurone ad
essa associata.
Convergenza anatomica: se su un neurone convergono i campi recettivi di +
neuroni, esso ha un campo recettivo + grande.
Nel campo recettivo un neurone può avere zona eccitatoria e inibitoria
Interneuroni inibitori
Campo recettivo eccitatorio
Campo recettivo inibitorio
32
Fibre periferiche nel sistema somatosensoriale hanno terminazione
specializzata in recettore. Arrivano al midollo spinale attraverso le radici
dorsali. Prima di entrare, ogni fibra afferente (mielinizzata) passa da un
ganglio (insieme dei corpi cellulari delle fibre afferenti primarie)
Dalla periferia vanno al midollo spinale (dal recettore al SNC),
trasportano l’info trasdotta verso il centro nervoso. Fanno sinapsi su un
neurone secondario.
Nervi spinali = parte afferente + parte efferente
X studiarle devo
x studiarle devo tagliare
tagliare la radice ventrale
la radice dorsale
Mettiamo elettrodi superficiali stimolanti e registranti (questi + vicini al
midollo spinale)
se stimolo il nervo dall’esterno, la corrente induce genesi
di pot. d’azione nelle fibre del nervo che viaggiano in direzione del midollo
spinale. Sono registrati dagli elettrodi registranti (la somma dei potenziali
d’azione che stanno passando rappresenta l’attività del nervo in quel
momento)
Onda α molto precoce + onde tardive successive
Ipotesi : 1. l’elettrodo stimolato ha causato potenziali d’azione a ripetizione
che generano le due onde
2. con la stimolazione, stimolo fibre di calibro diverso
diversa
velocità di conduzione
fibre che conducono a
maggiore velocità
fibre che conducono a
minore velocità
Allontano i 2 elettrodi
se è vera l’hp 1, non deve esserci differenza tra la
comparsa delle 2 onde. NON E’ VERO
Se la differenza temporale cambia
le onde tardive sono dovute a diverse
velocità di conduzione. E’ VERO
L’ipotesi 2 è vera!
Il nervo è costituito da fibre (di calibro diverso, che hanno diametro
diverso) che conducono a velocità diversa!
RIFLESSI: pag. 47 rizzolatti
33
Stimolazione della branca afferente delle radici dorsali
onde a livello
ventrale.
Renshaw: studi sul ritardo sinaptico (riflesso mono- o polisinaptico)
- FACILITAZIONE
Le afferenze sensitive influenzano una popolazione di motoneuroni
Se io stimolo elettricamente la fibra afferente A, posso registrare un certo
tipo di risposta nella fibra efferente
Eccita la popolazione di motoneuroni
che risponde con fibre efferenti
Se stimolo contemporaneamente l’afferenza A e B, la risposta
è più alta della somma dei 2 effetti singoli dovuti alla stimolazione
di A o B
a livello della radice ventrale l’onda registrata è più alta
Si sommano degli EPSP che, se ottenuti dalla stimolazione di una sola fibra
sensitiva, non riuscirebbero a raggiungere la soglia x generare il potenziale
d’azione.
Se le fibre sensitive sono stimolate insieme, l’effetto finale è
maggiore della somma degli effetti delle singole fibre.
A
B
Fenomeno che si ha se si stimolano
fibre appartenenti allo stesso nervo
muscolare.
Tanti motoneuroni vengono reclutati
x dare una determinata risposta.
- INIBIZIONE
Muscolo gastrocnemio (nel polpaccio) fa flettere il piede verso il basso.
Muscolo tibiale anteriore determina l’effetto opposto.
La radice ventrale del gastrocnemio non registra risposta se stimolo le fibre
sensitive provenienti dal tibiale anteriore.
Se stimolo insieme fibre sensitive provenienti da muscoli
opposti, la risposta è più bassa
questi due circuiti si inibiscono
reciprocamente
- OCCLUSIONE:
34
Stimolando insieme due fibre sensitive ( radicola A e B)
risposta
che rappresenta una sola onda.
Le 2 popolazione di neuroni hanno una sovrapposizione, ma in realtà la
somma xmette l’attivazione di un solo motoneurone in più.
Non c’è un effetto facilitatorio.
Una delle 2 fibre sensitive fa nascere il potenziale d’azione nella popolazione
controllata da entrambe
si impedisce la generazione di un altro potenziale
da parte dell’altra fibra (refrattarietà). La prima fibra porta già la popolazione
soprasoglia e quindi non è più influenzabile.
Facilitazione
Occlusione
eccitazione sottosoglia
eccitazione soprasoglia
effetto
dominante
Entrambe sono conseguenza di impulsi eccitatori sugli stessi motoneuroni
Riflesso POLISINAPTICO: x avere la risposta del motoneurone ci vuole +
tempo e l’onda a livello delle radici ventrali è + prolungata.
+ vedi pag. 59 rizzolatti
AFFERENZE DELLA SENSIBILITA’ SOMATICA:
Percezione
necessaria l’elaborazione della corteccia cerebrale (
cosciente). Dalle afferenze sensitive all’elaborazione corticale.
stato
Info che provengono dalla cute (tatto/pressione, carattere vibratorio,
propriocettivo, termico, dolorifico, prurito)
Arrivano al midollo spinale attraverso le radici dorsali
Le fibre afferenti hanno diverse terminazioni a livello del midollo spinale:
- le fibre Aδ e C (dolorifico/termico) terminano sulla lamina I e V.
- le afferenze propriocettive e tattili terminano nelle lamine III IV e in parte V
e VII
nucleo della colonna di Clarke
nucleo proprio
Corno dorsale: parziale aggregazione delle afferenze sensitive
ventrale
(riflesso)
quelle
che
arrivano
fanno
sinapsi
direttamente
direttamente
coi
nel
corno
motoneuroni
35
Fasci ascendenti
Lemnisco
mediale
bulbo
talamo
Antero
laterale
corteccia
localizzazione opposta rispetto a quella
delle fibre afferenti
SISTEMA LEMNISCO-MEDIALE:
Info tatto-pressorie e propriocettive provengono da tutto il corpo.
Le afferenze somatosensitive entrano nelle radici dorsali, poi:
1. circuito diretto
le afferenze sensitive viaggiano verso il bulbo
senza fare sinapsi nel midollo spinale. Si immettono direttamente nei
cordoni posteriori. - Nucleo gracile
fascicolo gracile
ricevono info
tattili
dagli arti superiori
- Nucleo cuneato
fascicolo cuneato e inferiori
2. circuito indiretto
le afferenze sensoriali fanno sinapsi nel corno
dorsale. I neuroni delle corna dorsali manderanno le fibre nei cordoni
posteriori
Ma c’è un’ ECCEZIONE!!!!!
Nella propriocezione, ci vuole distinzione tra:
- arti superiori
afferenze sensitiva che arriva al midollo cervicale,
arriva poi al nucleo gracile e cuneato
- arti inferiori
l’afferenza sensitiva fa un breve tratto nel fascicolo
gracile e termina nella colonna di Clarke. I neuroni della colonna di
Clarke fanno arrivare le fibre nel nucleo zeta (nel bulbo) e al cervelletto
Nucleo gracile
arti inferiori, modalità tattile
Nucleo cuneato
arti superiori, modalità tattile e
propriocettiva
Nucleo zeta
arti inferiori, modalità propriocettiva
Proprietà lemniscali:
i neuroni del sistema lemniscale (dei nuclei gracile e cuneato)
rispondono a un'unica submodalità
sono molto specifici
hanno campi recettivi molto piccoli
elevata acuità
discriminativa
rispondono costantemente all’applicazione dello stimolo
non c’è abitudine, anche se gli stimoli sono molto ravvicinati.
Hanno una risposta ad elevata sicurezza.
36
Il fascio lemnisco-mediale termina a livello del talamo (complesso ventrobasale, diviso in: - VPL: ventro-postero-laterale
- VPM: ventro-postero-mediale
qui terminano le info di
tipo somatosensoriale
i neuroni proiettano fibre verso
della testa
la corteccia somatosensoriale primaria
SISTEMA ANTERO-LATERALE:
Le afferenze sensitive entrano nel midollo spinale tramite le radici dorsali.
L’afferenza sensitiva primaria fa sinapsi nel corno dorsale del midollo.
Il sistema si incrocia subito nel midollo, non nel bulbo come nel
lemnisco-mediale.
Ha una prima tappa nel sistema retinale del tronco dell’encefalo
(spinoreticolare)
La seconda tappa è nel talamo (paleospinotalamica)
Termina nel complesso ventrobasale in neuroni posteriori (neospinotalamica)
Complesso posteriore del talamo: centro del dolore
I neuroni di questo sistema possono essere polimodali
Il sistema anterolaterale ha neuroni con campi recettivi grandi e si abituano
alla risposta dello stimolo.
Aspetto in comune dei due sistemi: percezione del tatto.
IL SIST. LEMNISCO-MEDIALE RISPONDE AL TATTO
(discriminativo)
EPICRITICO
IL SIST. ANTEROLATERALE RISPONDE AL TATTO PROTOPATICO (più
grossolano, dà info generali sulla localizzazione). Ma una sua componente
che serve x localizzare lo stimolo (la neospinotalamica) può portare anche al
tatto epicritico.
Il sistema anterolaterale è più antico di quello lemniscale. Con l’evoluzione si
è resa necessaria una maggiore raffinatezza x l’importanza del movimento.
Se c’è una lesione che interessa un sistema, c’è l’altro che interviene in
sostituzione parziale.
RIASSUMENDO….
Sistema colonne dorsali- lemnisco mediale:
Percorsi:
37
- tatto-pressione (arti e tronco) e senso di posizione (arti superiori):
a) diretto: fibre periferiche (afferenti primarie)
colonne dorsali
ipsilaterali
nuclei gracile e cuneato ipsilaterali
talamo controlaterale
b) indiretto: fibre periferiche
sinapsi con neuroni di II ordine nelle
corna dorsali
colonne dorsolaterali
nuclei gracile e cuneato
talamo controlaterale
- senso di posizione (arti inferiori):
Fibre periferiche
sinapsi con neuroni di II ordine del nucleo di Clarke
(zona intermedia)
colonna di Clarke
nucleo Z ipsilaterale
talamo
controlaterale
Sistema anterolaterale:
Dolore, temperatura e tatto grossolano.
Le fibre si incrociano immediatamente all’interno del midollo. Percorso
indiretto:
Fibre periferiche
sinapsi con neuroni di II ordine nelle corna dorsali
colonne antero-laterali controlaterali
sostanza reticolare e talamo
Talamo: nuclei specifici e nuclei aspecifici. Ricostruzione completa di tutta la
superficie corporea.
Testa/ faccia rappresentate nel tronco dell’encefalo.
TRONCO DELL’ENCEFALO:
Struttura abbastanza primitiva, ma diversa dal midollo spinale.
Implicato in attività motorie, coinvolto in funzioni vegetative, collegato con
encefalo superiore e con il midollo spinale. contiene numerosi aggregati
cellulari (nuclei) con funzioni diverse ed è attraversato da numerose fibre.,
soprattutto nel ponte.
Nervi cranici: portano modalità sensoriali di faccia/testa, legati ad alcuni sensi
dell’individuo.
Mesencefalo + ponte + bulbo
Da qui escono i nervi cranici
sensitivi
di moto
38
misti
sono numerati con numeri romani. Sono formati da diversi fascicoli
che poi si riuniscono in un unico nervo. Le fibre entrano a livello del tronco,
collegate a cellule sensoriali o motorie.
Nel tronco dell’encefalo sono 10 + 2
Nervo olfattivo ( nel proencefalo)
Nervo ottico ( nel diencefalo)
I nervi spinali sono sempre misti (afferenze + efferenze)
Movimenti oculari: determinati da muscoli oculari legati al globo oculare ma
non influenzati dal moto articolare (dunque più rapidi)
Movimenti oculari riflessi
Diversi circuiti
Movimenti oculari volontari
Fibre afferenti del nervo fanno sinapsi su determinate cellule del tronco
dell’encefalo.
Fibre efferenti nascono da cellule di tipo motorio presenti nel t.d.e.
I nuclei del t.d.e. sono chiamati con il nome dei nervi.
I nuclei sensitivi e motori dei nervi cranici hanno un’organizzazione
particolare che dipende da come essi si sono disposti nell’embriogenesi. Sono
disposti in colonne longitudinali che percorrono il t.d.e.
Nuclei motori: zona mediale.
Nuclei sensitivi: zona laterale
Trauma cranico
coma + perdita di alcuni riflessi (es. riflesso pupillare)
SOSTANZA RETICOLARE DEL TRONCO: i suoi neuroni hanno assoni molto
lunghi che interessano molte strutture del tronco dell’encefalo. Ha un aspetto
reticolato e ha caratteristiche diffuse. Ha influenza non specifica ( aspetti
motivazionali del sistema motorio, cambiamento di stato attenzionale, vari
stadi del sonno.)
noradrenalina
Produzione di neurotrasmettitori (catecolamine)
adrenalina
+serotonina
Dopamina
Queste sostanze intervengono in processi globali.
39
Funzioni che hanno a che fare con modulazioni.
Es. motoneurone silente: la sua attività parte quando riceve afferenze
Motoneurone in stato di facilitazione: il suo potenziale di riposo è più
basso del normale
può essere eccitato più facilmente
questa
facilitazione è dovuta ad uno stato comportamentale (motivazione
i
motoneuroni sono più pronti rispetto ad una situaz di riposo)
Locus coeruleus: collegamenti discendenti col midollo spinale +
collegamenti con l’encefalo e con il cervelletto. Produce prevalentemente
noradrenalina.
Nuclei del Rafe(nella posizione più centrale del tronco): producono
serotonina. Sono distribuiti tra il mesencefalo e il bulbo. Hanno varie
conessioni:
1) Discendenti dai nuclei posteriori: influenzano le corna dorsali
(info sensitive). Non hanno influenza sul sistema tattile o
propriocettivo, ma solo sul sistema dolorifico.
Producono
analgesia
alzano la soglia x la percezione del dolore. Bloccano
l’info dolorifica a livello delle corna dorsali.
2) Ascendenti dai nuclei anteriori: connessioni con corteccia
cerebrale
funzione attivante. Influenza di carattere
modulatorio. Possono cambiare lo stato di coscienza.
3) Con il cervelletto.
I nuclei del Rafe, come il locus coeruleus, hanno un ruolo nelle fasi del sonno
(fenomeno attivo). I sistemi catecolaminergici sono fondamentali.
Substantia nigra (parte dei gangli della base): i suoi neuroni producono la
dopamina che interviene in un circuito motorio che, se leso, determina il
morbo di Parkinson.
Sistema dopaminergico: produce dopamina. Nuclei che hanno influenza sul
sistema libico (primitivo dal punto di vista evolutivo, ha a che fare con le
emozioni). Influenzano anche il sistema motivazionale. Importanza nei
processi cognitivi e di consolidamento mnemonico.
Serotonina e dopamina
ruolo importante nelle forme psichiatriche
(psicosi, schizofrenia, disturbi dell’umore)
ALTRI NUCLEI:
Sul mesencefalo
4 rigonfiamenti: collicoli
inferiori(+ posteriori, tappa
del sist. uditivo)
Superiori (+ anteriori)
40
Collicolo superiore, diviso in
(fatto a strati)
Parte sensitiva
parte motoria
Parte sensitiva e motoria
- vi è rappresentata tutta l’info visiva + info somatosensoriale e acustica
- ci sono neuroni polimodali
rispondono contemporaneamente alle
varie info. Rispondono a stimoli congruenti tra loro (es. stimolo che
viene dalla stessa parte dello spazio sia visivamente che acusticamente)
- può ricevere queste info e fa partire movimenti riflessi che coordinano
movimenti della testa e degli occhi. Spesso interviene x via riflessa
(importante x controllo dei movimenti oculari)
Sostanza
grigia
periacqueduttale:
nel
mesencefalo,
all’acquedotto. Implicata in soppressione del dolore.
attorno
Nucleo rosso: struttura soprattutto motoria. Controlla i muscoli distali e
flessori
Oliva superiore: neuroni di tipo acustico
Nuclei pontini: stazione di ritrasmissione delle fibre che vanno dalla
corteccia al cervelletto.
Oliva inferiore (nel bulbo): tappa di passaggio delle fibre che vanno dal
midollo al cervelletto (fibre rampicanti)
FASCI DISCENDENTI: (importanti x controllo motorio)
1. SISTEMA VENTRO-MEDIALE (più vecchio). Controlla la muscolatura
assiale e prossimale:
- Tratto reticolo-spinale: nasce dalla sostanza reticolare
Componente
Componente
eccitatoria
inibitoria
(mediale)
- Fascio vestibolo-spinale: nasce dai nuclei vestibolari, va ipsi- e
controlateralmente sui muscoli estensori
Laterale
Mediale
- Via tetto-spinale: dal collicolo superiore, si incrocia e va nel
midollo cervicale controlaterale.
41
2. SISTEMA DORSO-LATERALE (più recente). Controlla la muscolatura
distale in modo fine:
- Nucleo rosso: le sue fibre si incrociano sotto il mesencefalo.
Termina sui motoneuroni che controllano muscoli distali.
- Fascio cortico-spinale: controllo diretto dei muscoli distali
SISTEMA VESTIBOLARE:
Si trova vicino all’apparato acustico. L’endolinfa può passare dall’uno all’altro
sistema. E’ collegato con la branca vestibolare del nervo VIII. E’ contenuto in
canali ossei, all’interno dei quali ci sono canali membranosi.
Recettori vestibolari: cellule modificate (II tipo)
alla loro estremità hanno
ciglia di diverse lunghezze, la più lunga è il chinociglio. Sono recettori
meccanici. Lo stimolo meccanico è uno spostamento delle ciglia dovuto a un
movimento relativo dell’endolinfa rispetto alla massa gelatinosa, che subisce
uno spostamento. Si trovano nei canali semicircolari, nell’utricolo e nel
sacculo.
Disposti su 3 piani dello spazio, perpendicolari tra loro. I recettori
si trovano a livello delle loro basi, nelle creste ampollari. Rilevano velocità
e accelerazione angolare
Le ciglia non sono direttamente immerse nell’endolinfa, ma nella massa
gelatinosa.
Aumento della frequenza di scarica delle fibre afferenti del nervo VIII:
spostamento delle ciglia verso il chinociglio
può liberare vescicole di
neurotrasmettitore, captato dai recettori delle fibre afferenti primarie
DEPOLARIZZAZIONE
Diminuzione della frequenza di scarica: spostamento delle ciglia in direzione
opposta al chinociglio
IPERPOLARIZZAZIONE
I neuroni del sistema vestibolare a riposo non sono silenti, producono un
certo numero di potenziali d’azione, hanno attività intrinseca.
Canali semicircolari (registrano i movimenti angolari della testa)
Orizzontale
Anteriore
Posteriore
(uno x lato di ogni tipo,
agiscono in coppia)
42
polarità diretta
verso il sacculo
e l’utricolo
di sinistra
di destra
fa coppia
con
di destra
di sinistra
polarità diretta in maniera contraria
Utricolo e Sacculo (sensibili alla posizione statica della testa nello spazio,
sensibili ad accelerazione e velocità lineare)
macula
orizzontale
macula
verticale
spostamento
Hanno ciglia immerse in una massa gelatinosa
che contiene cristalli di carbonato di calcio
(otoliti).
Spostamento
degli
otoliti
delle ciglia.
Equilibrio: risultato finale dell’azione dei recettori vestibolari
una risposta motoria che tende a ristabilire ordine.
produciamo
Le fibre afferenti primarie portano info nei NUCLEI VESTIBOLARI del
tronco dell’encefalo
Sono 4 per parte
superiore
riceve info di tipo
riceve
angolare
dei canali semicircolari
laterale
mediale
info dall’utricolo riceve
info
(info
angolari
di
inferiore
solo
influenze
tipo
angolare)
uscite verso i
uscite ai muscoli
oculomotori
alla
formazione
muscoli estensori
degli arti x ristabilire
l’equilibrio
e lineari
uscite al midollo
cervicale x riportare
il
capo
originale
+
reticolare
in
uscite
posizione
ai
muscoli
43
oculomotori
Es. movimenti oculari: movimento angolare verso sinistra
spostamento
occhi verso il lato opposto (destra). Retto mediale e laterale controllati da
nuclei oculomotore e abducente.
Necessario eccitare 2 nuclei e inibirne altri 2
controllo incrociato dei
muscoli oculari.
I nuclei vestibolari sono influenzati da afferenze vestibolari provenienti dal
labirinto.
CERVELLETTO:
Sopra il tronco dell’encefalo, nell’uomo in parte coperto dagli emisferi
cerebrali. Struttura abbastanza arborizzata. Coinvolto in funzioni motorie. Usa
l’info afferente x esplicare controllo sul movimento:
- Agisce da comparatore: riceve l’info sul programma motorio che
l’individuo vuole eseguire + info sensoriali su quello che avviene
quando questo movimento viene eseguito. Se durante l’azione ci sono
perturbazioni non previste
il cervelletto le confronta con la
situazione prevista
corregge il modello mentre l’azione sta
avvenendo (controllo e correzione della durata e della forza dei
movimenti)
- Funzione di apprendimento motorio e pianificazione del movimento.
Occupa la fossa cranica posteriore.
E’ formato da 3 LOBI:
1) anteriore (paleocerebello)
2) posteriore (neocerebello)
3) flocculo nodulare ( archiocerebello)
Nomenclatura funzionale:
1. vestibolo cerebello (flocculo nodulare)
collegato con
il vestibolo
2. cerebrocerebello (2 zone laterali)
collegato con
corteccia cerebrale
3. spinocerebello (centrale)
collegato con midollo
spinale
44
Organizzazione:
- corteccia cerebellare
- 3 paia di nuclei profondi (sotto la corteccia cerebellare)
nucleo del fastigio (collegato col verme)
nucleo interposto (formato da globoso e emboliforme)
nucleo dentato (collegato con zone laterali)
- Efferenze: ai nuclei profondi (cellule del Purkinje)
- Afferenze: dalla periferia e da tutti i livelli del SNC (sinapsi con
nuclei profondi e con la corteccia cerebrale)
Il cervelletto si trova al di sopra del ponte. E’ già abbastanza sviluppato
attraverso tutte le classi di vertebrati (l’apprendimento motorio esiste anche a
livelli bassi nella scala dei vertebrati)
Parte interna, centrale: verme
Due zone esterne, laterali: emisferi cerebellari
inibitorio
Rappresentazione schematica di una cellula del Purkinje:
unico neurone di uscita della corteccia cerebellare,
Vestibolo cerebello
influenza sui nuclei vestibolari superiore e mediale
Il cervelletto riceve afferenze vestibolari.
Cervelletto: non inizia il movimento, non è il centro fondamentale dei riflessi
Agisce come comparatore: confronta lo spostamento della testa con quello
che deve essere quello degli occhi, se questo non è giusto lo corregge
fa
assumere agli occhi la posizione giusta. Interviene perché il riflesso sia
accurato.
Serve x stabilizzare la testa (agendo attraverso i muscoli del collo)
Afferenze somatosensoriali: dal midollo nascono 4 fasci spinocerebellari
(nella sostanza bianca laterale, ad es. ne fa parte la colonna di Clarke)
+ info ricevute dal trigemino (testa/faccia)
Portano al cervelletto info
dalle diverse parti del corpo
sulle submodalità somatosensoriali
45
Zona del verme + parte di zona intermedia degli emisferi (cioè lo
spinocerebello) rispondono a stimolazione somatosensoriale. Qui arrivano
afferenze dal midollo spinale.Ci sono 2 rappresentazioni somatotopiche (di
tutto il corpo) opposte.
Corteccia cerebellare
Le afferenze spinocerebellari arrivano a
Nuclei profondi
Nucleo del fastigio: controllato direttamente da cellula del Purkinje. Manda
verso regioni del tronco dell’encefalo che influenzano il midollo spinale e
verso la corteccia cerebrale (aree 4 – 6
controllo volontario del
movimento). In particolare, manda ai nuclei vestibolari e alla formazione
reticolare al lato ipsi- e controlaterale
controllo dei motoneuroni che
controllano gli arti (controllo muscoli assialie prossimali)
Nucleo interposito: riceve da zona intermedia del cervelletto. Le sue uscite
terminano sul nucleo rosso controlaterale ( controllo sui muscoli flessori e
distali controlaterali rispetto al nucleo rosso) + corteccia motoria (4 – 6).
Influenza i processi di partenza del movimento.
La corteccia cerebrale influenza il nucleo dentato attraverso i nuclei pontini,
che ricevono da aree della corteccia e mandano msg verso gli emisferi
cerebellari. La corteccia informa il cervelletto sulle sue decisioni e sulle
conseguenze di una determinata decisione
risposta del cervelletto
Talamo
corteccia (riceve info di ritorno e può variare il suo
programma tramite uscite dirette verso il midollo spinale)
Conrollo ipsilaterale: il nucleo dentato di sx controlla il midollo spinale di sx.
Corteccia cerebellare: esternamente suddivisa in folia. Ha un suo
spessore, sotto c’è sostanza bianca (fibre)
Struttura interna della corteccia cerebellare:
Tre strati:
1) strato dei granuli (più profondo)
2) strato delle cellule del Purkinje
3) strato molecolare
1) contiene i granuli (eccitatori). Al suo interno c’è la cellula del Golgi
(interneurone inibitorio)
2) contiene solo le cellule del Purkinje (sempre inibitorie. Utilizzano in
neurotrasmettitore GABA. Hanno albero dendritico disposto su un piano
perpendicolare all’asse lungo del folium. Influenzano i nuclei profondi, sempre
eccitatori)
cellula a canestro(manda il suo
46
3) contiene due tipi di interneuroni inibitori
attorno
assone
cellula stellata
Entrate alla corteccia cerebellare:
- FIBRE MUSCOIDI
uniche afferenze
- FIBRE RAMPICANTI
del cervelletto
al corpo
cellulare della
cellula del P.)
Le fibre muscoidi fanno sinapsi con le cellule dei granuli.
I granuli mandano assoni allo strato molecolare
si dividono a T,
originando fibre parallele (all’asse lungo del folium), e perpendicolari al piano
delle cellule del Purkinje (influenzate dalle fibre parallele e dalle cellule dei
granuli). Le fibre parallele agiscono molto vicino al corpo cellulare. I granuli
sono anche influenzati in maniera inibitoria dalle terminazioni delle cellule del
Golgi.
Glomerulo cerebellare = fibre muscoidi + granuli + assoni delle cellule del
Golgi
Le fibre muscoidi portano info del midollo spinale, del tronco dell’encefalo,
delle zone superiori del t.d.e. (es. mesencefalo)
Le fibre che provengono dai nuclei pontini sono muscoidi!!!!!!
Le fibre rampicanti agiscono direttamente sulla cellula del Purkinje
(sul corpo cellulare o vicino). Vengono dall’oliva inferiore (riceve info dal
midollo spinale e dal nucleo rosso –fibre spino-olivari–)
La risposta del cervelletto deriva dai nuclei profondi (devono essere disinibiti
dal controllo delle cellule del Purkinje, che devono essere inibite
inibizione di una cellula inibitoria determina un’eccitazione)
La corteccia cerebellare è sempre uguale in ogni sua parte.
POSTURA:
Arrivo di segnali somatosensoriali che informano sui segnali dell’ambiente
esterno + previsioni di modifica del nostro equilibrio.
Meccanismi e feedback (retroattivo) e a feedforward (anticipatorio)
Segnali afferenti importanti x mantenimento della postura: somatosensoriali,
vestibolari, visivi
47
Riflessi che intervengono nel controllo della postura (dal sistema vestibolare
e dal sistema cervicale):
vestibolo-cervicali
modificano la
posizione
Segnali vestibolari
riflessi
della testa in seguito a
una perturbazione
vestibolo-spinali
Stimolazione di
articolazioni cervicali
riflessi
cervico-cervicale
arti, ha
impostano tono e
contrazione muscolare
degli arti dopo uno
spostamento della testa
sinergico con il
vestibolo-cervicale
cervico-spinale
agisce sugli
effetto opposto a quello vestibolo-spinale
a seguito di uno stesso tipo di stimolazione
es. Corpo che cade in avanti con la testa che rimane in asse rispetto al corpo
vengono stimolati i recettori vestibolari
Di solito c’è una combinazione dei due riflessi.
I sistemi ventromediali (vestibolo-spinale, tetto-spinale, reticolo-spinale)
hanno influenza sui muscoli di arti e testa.
PREPARATO DECEREBRATO:
Sezione perpendicolare all’asse lungo del tronco dell’encefalo:
1) diencefalo / mesencefalo
2) in mezzo ai collicoli (preparato intercollicolare)
3) mesencefalo / resto del tronco
L’effetto della decerebrazione è più forte in 3)
sezione più posteriore
Animale in rigidità estensoria (arti posteriori, arti anteriori e collo(opistotono))
E’ la “caricatura” di un preparato fisiologico, sta in piedi in una situazione che
esagera la sit. fisiologica.
Decerebrazione anemica: chiusura dei rami della carotide
non arriva più
sangue alla parte anteriore del tronco dell’encefalo
preparato decerebrato
48
Rigidità estensoria: aumento del tono dei muscoli estensori dovuto a:
- rigidità α (alfa)
- rigidità γ (gamma)
interrotta la branca afferente del riflesso da
stiramento (esagerazione del riflesso da stiram.)
Taglio delle radici dorsali del midollo spinale toracico e lombare
rigidità arti
Uscite del cervelletto: inibitorie (cellule del Purkinje)
Se è tolto, si toglie un’inibizione
si esalta rigidità estensoria
Fuso stirato
scarica fibra afferente Ia.
Se taglio le radici dorsali tolgo le afferenti Ia.
Muscolo in contrazione (coattivazione dei motoneuroni α e γ)
motoneuroni γ
eccitazione del fuso (che sarebbe contratto)
I motoneuroni γ, in rigidità estensoria da decerebrazione, sono più facilitati
del normale
eccitano di più il fuso
le fibre afferenti Ia hanno
frequenza di scarica maggiore
terminano sui motoneuroni α
maggiore contrazione del muscolo estensorio.
Se ledo il nucleo di Deiters (vestibolare laterale, ha anche componente
inibitoria)
perdita della rigidità estensoria dell’arto ipsilaterale.
Se ledo quello dell’altro lato
la rigidità ricompare (dovuta a sostanza
reticolare)
Parte pontina (eccitatoria)
Sostanza reticolare
Parte bulbare (inibitoria)
Tronco dell’encefalo: azione facilitatoria su muscolatura estensoria.
Influenze su motoneuroni α:
in continua eccitazione dalle afferenze
eccitatori - nucleo vestibolare laterale
- formazione reticolare pontina
inibitori
formazione reticolare bulbare
ipsilateralmente su α
su α da entrambi i lati
su α
49
Nel preparato decerebrato emerge di più la componente facilitatoria.
Nucleo rosso: - eccitatorio sui motoneuroni flessori
- inibitorio sui motoneuroni estensori
Rigidità α e γ non sono dovute al cervelletto!!!!!! Il cervelletto è inibitorio sui
nuclei vestibolari.
La postura è controllata da diversi livelli dell’encefalo
Effetti della LESIONE CEREBELLARE: i deficit maggiori sono di tipo motorio
(equilibrio, andatura, postura eretta, inizio e corretta temporarizzazione del
movimento, capacità di articolare il linguaggio)
effetti collegabili alle varie
sezioni del cervelletto.
CORTECCIA CEREBRALE:
Appartiene al telencefalo, insieme ai gangli della base. E’ formata da sostanza
grigia, spessore 2,5- 4 mm
6 strati (isocortex o neocortex 95%)
+
3 strati (allocortex 5%)
prevalenza di cellule piramidali, manca il 4°
Notevoli variazioni nei 6 strati
strato
corteccia motoria agranulare
presenza granuli, 4° strato
granulare o koniocortex
corteccia
Corteccia eterotipica
Agranulare
(priva del 4°strato,
area motoria,
prevalenza di cellule
piramidali soprattutto
nel 5° strato di uscita)
Granulare
(aree sensoriali primarie
forte enfasi del 4°strato
somatosensoriale
visiva
acustica
riceve dai nuclei talamici specifici sensoriali)
La corteccia cerebrale è suddivisibile in aree (citoarchitettonica). La
suddivisione in aree citoarchitettoniche è fatta in base alla forma delle cellule
Mappe citoarchitettoniche
50
Brodmann ha individuato diverse aree citoarchitettoniche (inizio ‘900), con
una procedura di taglio orizzontale (dalla sommità alla base della corteccia)
Le diverse aree hanno corrispondenza con i solchi.
Solco centrale (di Rolando) divide l’area 4 di Brodmann (motoria primaria) da
3,2,1 (somatosensoriale primaria)
Solco di Silvio divide lobo frontale/temporale dal parietale.
C’è corrispondenza tra mappe citoarchitettoniche e funzionali.
Differenza con la corteccia cerebellare, che è uguale dappertutto!! Quella
cerebrale è diversa
facilità di riconoscimento delle varie aree in base a
struttura.
Possibile fare confronti con specie diverse dall’uomo (es. scimmia:
somiglianze e differenze)
Anche qui il solco centrale divide le stesse aree dell’uomo.
Differenze uomo/scimmia:
- area visiva primaria: nella scimmia è primario il lobo occipitale,
nell’uomo ne occupa una piccola parte, ha anche posizione interna.
Causa: movimento di rotazione
- nell’uomo: espansione della zona parieto-temporo-occipitale,
maggiore sviluppo di funzioni che partono da substrato sensorimotorio
simile alla scimmia, ma che hanno subito modifiche a livello percettivo
- nell’uomo: ampliamento del lobo pre-frontale (parte più rostrale del
lobo frontale)
Ci sono aree che nella scimmia sono allo stato primitivo e nell’uomo si sono
espanse, pur conservando traccia della funzione originaria.
Lobotomia: turbe dell’umore, incapacità di pianificazione
Per molto tempo si è pensato che la corteccia cerebrale fosse un sistema
dove l’info sensoriale proveniente dal talamo terminasse su aree sensoriali
(parte posteriore) dove veniva elaborata x essere infine percepita
(elaborazione seriale che aumenta di complessità). Si pensava che il frutto di
quest’elaborazione andasse nel lobo frontale (aree motorie) che comandava
il movimento + presenza di aree associative (né sensoriali né motorie), che
51
associavano diverse risposte sensoriali tra loro e queste ultime a comandi
motori.
Questo concetto seriale è stato radicalmente cambiato negli ultimi 20-25
anni:
molte aree chiamate associative non sono polimodali, ma hanno
caratteristiche più complesse delle sensoriali primarie che ricevono info
direttamente dal talamo (aree sensoriali di ordine superiore)
evoluzione di studi neuroanatomici: tracciare connessioni all’interno del
SNC tra le aree posteriori (ritenute sensoriali) e quelle anteriori
(ritenute motorie)
ci sono connessioni reciproche, costituite da
numerosi circuiti (quindi anche gli aspetti motori influenzano quelli
sensoriali)
L’organizzazione della corteccia è da intendersi come un processo che si
svolge in parallelo (diversi moduli corticali che agiscono in parallelo)
Le aree sensoriali e motorie sono state suddivise notevolmente e le aree
associative ridotte.
Linguaggio: aree collegate tra loro che svolgono diverse funzioni (recepire da
pt di vista visivo/acustico, produrre…)
CORTECCIA SOMATOSENSITIVA PRIMARIA:
Dietro al solco centrale di Rolando, superficie laterale e mediale dell’emisfero.
Può essere limitata posteriormente dal solco post-centrale.
Se si fa una sezione sagittale (parallela al piano che divide in 2 il corpo), si
ottiene una fettina di sottile spessore corticale:
contiene 4 aree citoarchitettoniche e funzionali:
- 3a (profondità del solco centrale)
- 3b (all’interno del solco, ma un po’ fuori)
- 1 (su superficie visibile della corteccia)
- 2 (su superficie visibile, in parte entra nel solco post-centrale)
52
L’info che arriva è molto selettiva e rispetta le proprietà dei recettori
processo che si avvale di diversi meccanismi a livello delle tappe del sistema
somatosensoriale.
Il segnale deve essere ben identificato.
Es. afferenze da colonne dorsali al nucleo cuneato
l’ info eccitatoria deve
essere conservata
intervengono meccanismi inibitori che contrastano il
segnale ( si eccitano anche interneuroni inibitori laterali che terminano su
neuroni laterali al neurone eccitato, che vengono inibiti)
Il segnale si “sente” meglio se perifericamente c’è inibizione (sistema a
feedforward, creato man mano che il segnale sale. In più c’è anche un
segnale a feedback originato dal neurone stesso. C’è anche un controllo
diretto da parte della corteccia.
Si crea una linea attivata che mantiene il
segnale.
Sulla corteccia c’è una rappresentazione somatotopica (omuncolo) che
riflette la capacità percettiva e motoria dell’individuo. E’ una rappresentazione
deformata delle proporzioni corporee, in base alla densità di innervazione. Ci
sono parti corporee più innervate di altre. La faccia è rappresentata più
lateralmente.
Utilizzando tecniche di registrazione microelettrica, si è visto che nella
corteccia somatosensitiva primaria esistono 4 omuncoli:
- area 3b e 1 (info tattili): due rappresentazioni appaiate, doppia
rappresentazione della stessa parte corporea
- area 3a e 2 (info dei fusi neuromuscolari e delle articolazioni)
Le sottorappresentazioni possono essere molto raffinate
Es. ratti: vibrisse usate come organi di esplorazione. Possono essere mosse
singolarmente
corteccia cerebrale divisa in cilindretti verticali, ognuno dei
quali corrisponde a un baffo.
Area 3a risponde a stimolazione dei fusi neuromuscolari
Area 3b risponde a stimolazione tattile superficiale (i campi recettivi
dei suoi neuroni sono piccoli)
Area 1 risponde a stimolazione tattile (campi recettivi più grandi) e a
stimoli in movimento su area cutanea
Area 2 risponde a stimolazione delle articolazioni
Modalità tattopressoria e propriocettiva. Hanno connessioni diverse
rispetto al talamo.
53
L’organizzazione intrinseca della corteccia cerebrale è molto precisa.
Osservando in profondità: i neuroni lungo una verticale hanno tutti la
stessa proprietà. Sono collegati tra loro da connessioni sinaptiche. Es.
neuroni che risp a uno stimolo con una scarica che perdura nel tempo o
neuroni a rapido adattamento.
Proprietà colonnare della corteccia (
in tutte le cortecce sensoriali e in molte aree della corteccia cerebrale. E’
un’organizzazione tipica.
E’ difficile trovare una rappresentazione precisa del dolore nella corteccia.
Afferenze talamiche arrivano soprattutto alle aree 3a e 3b, le quali mandano
a1e2
Maggiori capacità
Campi recettivi +grandi
discriminative
(convergenza anatomica)
maggiore complessità e astrazione
della risposta allo stimolo
maggiore livello percettivo
Percezione: si pensava fosse l’eleborazione finale di una serie di sensazioni x
coordinare poi attività motoria.
In realtà attività sensoriale e motoria non possono essere separate.
Deficit di natura sensoriale
In attivazione di area somatosensitiva primaria
(anestesia)
Deficit
a
livello
motorio
Info sensoriale è comunque necessaria x compiere un’attività motoria.
Le info sensoriali ispirano cambiamenti nel movimento.
Area somatosensoriale secondaria:lateralmente a quella primaria, un po’
nascosta nel solco laterale (di Silvio). Contiene neuroni che risp solo a
stimolazione tattile e non propriocettiva, che hanno campi recettivi più grandi
e hanno una rappresentazione bilaterale (ricevono info dall’arto ipsi- e
controlaterale). E’ connessa con aree della corteccia motoria. E’ coinvolta in
azioni esplorative.
Lesione area 43
astereognosia: incapacità di riconoscere aspetto
tridimensionale degli oggetti.
54
Cortecce parietali posteriori (dietro la S-I, formano lobo parietale posteriore,
nell’uomo sono le aree 5,7,39,40) ricevono info somatosensoriale, visiva e
sono collegate all’area motoria.
Sindrome da eminegligenza spaziale: il paziente si comporta come se
ignorasse lo spazio controlaterale alla lesione.
OTTICA:
Il sistema visivo è quello studiato di più.
La retina ha 6 strati. Tre contengono i corpi cellulari dei neuroni. Gli strati
plessiformi contengono le connessioni dei neuroni.
I recettori si trovano nella parte più posteriore di questi strati.
Coni: visione diurna, rispondono bene alla luce assiale,alta acuità
visiva, bassa convergenza, sensibili a 3 fasce di lunghezza d’onda
Bastoncelli: visione crepuscolare, rispondono bene alla luce diffusa, b
bassa acuità visiva, la loro info converge su cellule, acromatici (un solo
pigmento: rodopsina)
Nella fovea ci sono solo coni, non bastoncelli.
Fotopigmento: molecola che capta i fotoni della luce
La risposta dei fotorecettori alla luce è un’iperpolarizzazione.
I bastoncelli al buio sono depolarizzati.
Fotorecettori
d’azione.
Fotorecettori
cellule bipolari
cellula gangliare
È l’unica che può produrre potenziale
Manda l’info verso i centri superiori.
eccitatoria
sinapsi con cellule bipolari
inibitoria
Fotorecettore iperpolarizzato: inibito il rilascio di glutammato inibitorio
è
una facilitazione
le cellule bipolari sono disinibite
le cellule gangliari
vengono facilitate.
Centro-on: una cellula gangliare ha campo recettivo che si eccita quando la
luce colpisce il centro.
55
Centro-off: il campo recettivo si inibisce quando la luce colpisce il centro.
Nervo ottico: nervo di uscita dell’occhio.
La retina accoglie tutta l’info visiva dell’ambiente esterno. Il fascio luminoso
attraversa la pupilla e termina sul fondo della retina. Il campo visivo è circa
180°: zona binoculare + 2 zone estreme monoculari
Finisce sui 2 occhi,
su retine diverse.
Se lo stimolo cade qui viene visto solo dalla
retina nasale dello stesso lato.
Le fibre provenienti da qui si incrociano
(chiasma ottico)
Ai centri nervosi arriva una fibra della retina temporale e una fibra della retina
nasale dell’altro occhio.
TRATTI OTTICI: fibre dopo il chiasma
NERVO OTTICO: fibre prima del chiasma
Due stazioni successive:
1) collicolo superiore
2) corpo genicolato laterale
1) si trova nel mesencefalo. Contiene una mappa completa dell’emicampo
visivo controlaterale. Riceve info grossolana riguardante forma generica e
movimento degli oggetti che serve a indirizzare occhi e testa.
2) Via retino-genicolo-striatale: collega la retina alla corteccia tramite il corpo
genicolato laterale. Esso ha una struttura a lamine e non a nucleo come gli
altri nuclei talamici. Ha 6 strati in senso ventro-dorsale, separati da lamine di
sostanza bianca (suddivisione funzionale): i primi 2 strati (magnocellulari)
contengono cellule più grandi degli altri 4 (parvicellulari). Questi strati
ricevono le info nasali controlaterali e temporali ipsilaterali in modo alternato,
le afferenze non si mescolano. Alla corteccia arrivano info sia magnocellulari
sia parvocellulari dall’occhio dx e sx.
L’info visiva prende due vie
Collicolo superiore (più antica)
Retino-genicolo-striata (più recente)
56
Via retino-genicolo-striata: due vie in parallelo
Via parvocellulare:
molto sensibile ai
dettagli, poco sensibile
al movimento.
Visione a colori.
Via magnocellulare:
sensibile al movimento,
forme grossolane.
Ad ogni livello ogni cellula ha una capacità di elaborazione maggiore di quelle
dei livelli inferiori.
Le cellule gangliari e del corpo genicolato laterale analizzano soprattutto
i contrasti di luminosità
Le cellule semplici (sensibili alle barrette orientate) e complesse
rimodellano queste info con campi recettivi in grado di raffigurare
segmenti netti e margini precisi.
Nella retina e nel CGL ciò che conta è la posizione dello stimolo.
Aumentando la gerarchia delle aree (V1
V5 -responsabile della precisione
del movimento delgi oggetti-) aumenta la grandezza dei campi recettivi. Un
campo recettivo più grande può far vedere un oggetto tutto intero.
AREE VISIVE:
– In una regione corticale citoarchitettonica uniforme possono essere
presenti diverse aree
– La differenza di un’area corticale visiva dipende da:
- le caratteristiche funzionali che la differenziano dalle altre
- gruppo specifico di entrata-uscita
- unica citoarchitettura
– Le aree visive variano come dimensione
– Ogni area visiva da V3 a V5 da V1 e V2 (che contengono tutte le
sottomodalità insieme)
– L’area V1 manda in uscita a tutte le altre aree.
La sensazione di movimento si ha xchè l’immagine si muove e lìocchio resta
fermo o xchè l’occhio si muove. L’immagine sulla retina si trova
temporalmente in punti diversi. L’analisi del movimento è di tipo temporale,
dato che lo stimolo si muove nel tempo.
57
ELABORAZIONE DEI COLORI:
Tre tipi di coni, ognuno analizza uno spettro diverso.
(opponenza cromatica, contrasto cromatico, contrasto di percezione dei
colori)
Cellule a opposizione spettrale doppia: info univoca
può risp a 2 colori
diversi
Cellule a opposizione spettrale semplice: info ambigua
Cellule a banda larga: contrasto di luminosità
Risposta massima quando i due colori cadono uno alla periferia, l’altro al
centro
contrasto che si mantiene anche se cambia la luminosità
In V1 le cellule a opposizione spettrale doppia si trovano nei blob (cilindri
che attraversano la corteccia verticalmente soprattutto in strati superficiali e
profondi, incastonati nelle colonne di orientamento. Processano i colori )
Zone interblob: processano la forma.
Le info che riguardano i colori terminano in V4(prima area visiva dove c’è
risposta al colore), che a sua volta proietta alla corteccia inferotemporale (che
contiene neuroni x il riconoscimento dell’oggetto)
La via magnocellulare porta in V5 dove si ha l’analisi del movimento degli
stimoli.
TEORIA DI LAND (TEORIA RETINEX):
E’ importante la luminosità (diversa da quella dello sfondo) di un oggetto (di
base, è riflessa). Essa dipende dalle intensità luminose che portano con sé le
varie lunghezze d’onda, le quali hanno intensità diverse. I contributi di
lunghezze d’onda diverse danno infine colore all’oggetto.
Il colore dell’oggetto emerge da un procedimento matematico. All’oggetto
sono assegnati 3 numeri (luminosità dell’oggetto in una certa lunghezza
d’onda). Ogni numero è confrontato con la luminosità dello sfondo (rapporto)
Sommatoria dei tre rapporti = colore dell’oggetto.
Il colore dell’oggetto è un contrasto di luminosità con lo sfondo. Colore
diverso a seconda dello sfondo su cui è collocato l’oggetto.
Quest’analisi avviene solo nell’area V4.
58
I pazienti che soffrono di acromatopsia (non riconoscono i colori) hanno
lesione nell’area V4.
Lesione area V5: deficit in percezione del movimento.
AGNOSIA: deficit di carattere percettivo (non sensoriale!!!!!!), per i vari tipi di
sottomodalità visive (trattate separatamente da vie poste in parallelo)
Daltonismo: problema genetico che riguarda i recettori
DUE TEORIE:
I) aree visive superiori
via del lobo inferotemporale (1)
via del lobo parietale posteriore (2)
(1) per percezione degli oggetti (riconoscerli e discriminarli)
(2) per percezione spaziale
Teoria proposta sulla base di pazienti che presentano dissociazioni percettive.
E’ la teoria del che cosa e del dove
(1)
(2)
via ventrale
qualità intrinseche
degli oggetti
via dorsale
localizzazione
spaziale degli oggetti
ci sono connessioni anatomiche reciproche
Noi abbiamo percezione unitaria.
Dal punto di vista funzionale (elettrofisiologia + studi su pazienti) questa
teoria può essere messa in discussione.
II) Altra teoria (Milner e Goodale 1993)
La via dorsale usa le proprietà degli oggetti x guidare i movimenti. E’ una via
di tipo prassico centrata sul soggetto. Estrae le proprietà spaziali degli oggetti
in relazione all’osservatore
trasformazione delle coordinate spaziali degli
oggetti in coordinate egocentriche riferite al corpo
programmazione del
tipo di prensione da utilizzare.
59
La via ventrale, come nella th precedente, fornisce una codifica centrata
sull’oggetto(rappresentazioni percettive e cognitive che riguardano le
caratteristiche degli oggetti e il loro significato)
AREE MOTORIE:
Corteccia agranulare (no 4°strato)
Va
Vb (cellule piramidali più grandi)
Omologia tra aree 4 e 6 della scimmia e dell’uomo. Si trovano davanti al solco
centrale (dietro c’è la corteccia somatosensoriale primaria. L’area 3a tocca
area 4 nel fondo del solco centrale)
Area 4: più posteriore, è l’area motoria primaria. Rappresentazione
dettagliata di alcuni tipi di movimenti.
Su parte visibile di emisfero:
Area 6: più anteriore, aree motorie superiori
area premotoria
(sinergiche)
Su parte invisibile:
area supplementare
Le aree motorie proiettano a diverse strutture.
FASCIO CORTICOSPINALE (o fascio piramidale xchè passa x parte
ventrale del bulbo dove ci sono le piramidi bulbari):
dalla corteccia al midollo spinale x influenzare i movimenti
Proviene anche da alcune aree dietro il solco centrale (area somatosensitiva
primaria, aree 5 e 7 della corteccia parietale posteriore).
Passa nelle colonne laterali. Si assottiglia scendendo lungo il midollo.
E’ fatto:
– per 2/3 da fibre provenienti da aree 4 e 6 (proiettano sia al midollo
spinale che ai nuclei motori del tronco dell’encefalo, x il movimento
della testa)
– per 1/3 da fibre provenienti dalla corteccia parietale.
Due componenti:
1) Tratto corticospinale laterale: 90% delle fibre. Nasce dalla
corteccia cerebrale, a livello delle piramidi bulbari si incrocia e
termina nel midollo spinale controlaterale. Si incanala lungo le
60
colonne laterali e invia i propri assoni alla sostanza grigia
corrispondente alle zone corporee da muovere. I motoneuroni
rispondono al comando della corteccia. Ci sono anche fibre
che terminano nel nucleo rosso (controllo muscolatura
flessoria, soprattutto distale)
2) Tratto corticospinale mediale: 10%. Rimane ipsilaterale.
Va nelle colonne ventrali del midollo. Termina su motoneuroni
che controllano muscolatura assiale. Ci sono anche collaterali
che terminano su sostanza reticolare.
Procedendo nelle scala evolutiva, si nota che il fascio corticospinale termina
in zone sempre più ventrali della sostanza grigia. In animali più primitivi,
influenza i motoneuroni in modo indiretto. Nei primati: influenza diretta sui
motoneuroni x controllo dei movimenti fini, ma soprattutto sugli interneuroni.
La corteccia attua controllo dei movimenti in modo indiretto attraverso il
tronco.
Proiezioni delle cortecce non motorie terminano nelle corna dorsali del
midollo spinale, nei nuclei gracile e cuneato e nei nuclei talamici sensoriali.
Non dirigono il movimento, ma controllano l’afferenza ( l’info sensoriale in
arrivo è controllata dalla corteccia). Facilitano/inibiscono l’entrata
sensoriale.
La corteccia motoria non ha il 4° strato. Le proiezioni talamiche arrivano nel
3° strato (parte + profonda)
Stimolazioni cutanee e propriocettive (muscolari e articolari) attivano anche la
corteccia motoria.
Meccanismi anticipatori (a feedforward)
Movimento volontario
Informazioni a feedback (su perturbazioni avvenute)
La corteccia motoria determina i movimenti. Influenza i motoneuroni spinali
relativi ai muscoli x il movimento che voglio compiere:
es. valutare estensione delle dita (ampiezza del movimento)
“
forza da imprimere
“
direzione del movimento
La corteccia motoria primaria codifica la forza.
Nucleo rosso e corteccia M1: neuroni capaci di registrare variazioni di forza
nel tempo.
61
Ampiezza: non è sicuro da cosa venga codificata.
Direzione: non esistono singoli neuroni che la codificano, ma popolazioni
(codifica di tipo grossolano)
I neuroni della corteccia somatosensoriale sono più fini.
Stimolazione superficiale
mosaico”
2 omuncoli. I movimenti sono rappresentati “a
Differenze tra cortecce premotorie ( PM) e corteccia motoria
primaria (M1):
- La microstimolazione evoca movimenti a bassa soglia nella M1, a soglia
mediamente più alta nelle PM.
- La microstimolazione di M1 evoca sempre movimenti, quella di PM non
sempre.
- La microstimolazione di M1 evoca movimenti semplici, discreti (es.
movimento di un singolo dito), quella di PM evoca movimenti +
complessi.
- Gli effetti della stimolazione elettrica si spiegano bene con le
connessioni corticodiscendenti di M1 e PM.
- Solo M1 ha proiezioni dirette monospinaptiche sui motoneuroni spinali.
PM ha molte connessioni indirette col midollo spinale (es. corticoreticolo-spinale)
- M1 non riceve info visive/acustiche, PM invece sì. PM sono collegate alle
cortecce parietali (circuiti parieto-frontali) in modo reciproco. Gli stimoli
sensoriali arrivati alle cortecce parietali possono arrivare a PM.
La corteccia prefrontale manda all’area 6 (le PM sfruttano info sensoriale x
tradurla in movimento)
-
riceve info di natura sensoriale
connessa con area 4 ( 6
4)
ha proiezioni dirette sul midollo e su circuiti di
formazione
reticolare(potrebbe
agire
indipendentemente da M1)
Nella corteccia prefrontale vengono pianificate le azioni. I programmi motori
più ristretti vengono fatti da PM: se il contesto è adatto, fanno sì che il
programma motorio venga eseguito. Importanza della motivazione,
dell’emozione x pianificare il movimento.
62
Concetto classico: PM codifica il movimento
di x sé non è finalizzato.
Si riferisce allo spostamento
Di una parte corporea
Di fatto PM sono aree legate al concetto di azione
movimenti finalizzati,
con un preciso scopo.
Può essere eseguita
con diversi effettori.
Una volta definito lo scopo dell’azione, bisogna definire il programma motorio
necessario x eseguirla.
Per raggiungere/afferrare un oggetto, bisogna tener conto di: distanza
dell’oggetto dal corpo, sua posizione spaziale, sue caratteristiche fisiche
In base a queste caratteristiche devono essere impostati i parametri:
forza, direzione, apertura/chiusura della mano, sinergia tra braccio e mano.
C’è una codifica neuronale dell’azione!
I movimenti possono essere uguali ma usati x scopi diversi.
Diverse situazioni sperimentali:
- stimolo presentato alla luce e la scimmia lo deve prendere
“
“
“
“ “
“ non lo deve prendere
“
“
al buio la scimmia lo deve prendere
In area premotoria c’è risposta da parte del neurone.
Quando vediamo un oggetto, attiviamo un circuito anatomico-funzionale x
andare a prendere l’oggetto e in più ci facciamo una rappresentazione
mentale dell’oggetto in termini motori (modo in cui prenderemmo
quell’oggetto), oltre che in termini visivi.
Fare programmi motori
Nel cervello ci sono vie sensorimotorie x
Dare
rappresentazioni
mentali
delle azioni
Altre differenze tra PM e M1:
- M1 ha il compito esecutivo di programmi motori già fatti, mentre PM
interviene nella programmazione.
63
- L’arrivo di info visive a diverse aree di PM e non a M1 determina il
coinvolgimento della sola PM nelle trasformazioni visuomotorie x azioni
di diverso tipo fatte con diversi effettori.
- La lesione focale di M1 produce ipotonia e paresi flaccida. La lesione di
PM determina deficit più complessi, spesso di tipo sensorimotorio.
- PM codifica l’azione finalizzata, M1 interviene negli aspetti esecutivi dei
singoli movimenti.
RIASSUNTO CORTECCIA CEREBRALE:
La corteccia cerebrale contiene vari tipi di aree:
- Aree sensoriali primarie
- Aree che elaborano l’info sensoriale
- Area motoria primaria (funzioni esecutive, produce movimenti discreti)
- Aree premotorie (programmano le azioni)
- Circuiti parieto-frontali (trasformano le info sensoriali in azioni
appropriate e operano trasformazione di coordinate)
- Corteccia prefrontale (pianifica le azioni, interviene nelle scelte, è
forse sede di working memory)
- Circuito libico (unisce la corteccia cingolata con centri sottocorticali –
ipotalamo, ippocampo, amigdala, talamo anteriore. Interviene nel
comportamento emozionale, in quello motivazionale, ed ha un ruolo
nella formazione della memoria)
Percezione: è il risultato non solo di un’elaborazione sensoriale, ma anche di
un’interazione di quest’ultima con le rappresentazioni motorie. Non serve solo
x il riconoscimento e la discriminazione, ma anche x l’azione.
64