Tegumento
e i suoi derivati costituiscono un sistema organico
estremamente variabile e adattabile e in virtù di questa grande
plasticità e adattabilità si può dire che nessun altro organo svolge
altrettante funzioni quanto quelle attribuibili al tegumento:
Delimitazione e protezione (scaglie, pigmento…)
Mimetismo e attrazione sessuale (pigmenti, ghiandole..)
Difesa/offesa (unghia, corna, artigli, ghiandole..)
Ricezione di stimoli (recettori e strutture sensitive)
Termoregolazione (penne, peli, vasi, ghiandole)
Regolazione idrosalina
Scambi gassosi
Nutrimento della prole (ghiandole mammarie)
Accumulo di sostanze di riserva (grasso nel derma)
Locomozione (squame, cuscinetti rugosi, ali...)
Pelle di rana: GH: ghiandole, MF: melanofori
Epidermide e derma costituiscono la pelle
Il derma è riccamente vascolarizzato e contiene le ghiandole che hanno
però origine dall’ectoderma epidermico
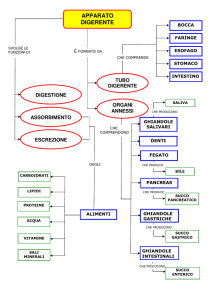
Tegumento e derivati
Epidermide derivati cornei
ghiandole del tegumento
Derma
scaglie dermiche
Dall’epidermide
Dal derma
L’epidermide
poggia sul derma ed è formata nei Vertebrati da un
epitelio pavimentoso pluristratificato più o meno corneificato in
superficie le cui tipiche cellule si chiamano cheratinociti.
La giunzione tra E/D – giunzione dermo-epidermica- è caratterizzata
da invaginazioni dell’E, i solchi epidermici, che si interdigitano con
evaginazioni del derma, le papille dermiche.
.
I cheratinociti si dispongono in strati, distinguibili perché per
un processo di citomorfosi gli strati più superficiali mostrano una
sempre più intensa cheratinizzazione.
Partendo dal più esterno si distinguono:
¾ Strato corneo
¾Strato granuloso
¾Strato spinoso
¾Strato basale sede del ricambio cellulare
Le cellule dello strato granuloso hanno granuli
di cheratoialina contenenti proteine, polisaccaridi e lipidi
Le cellule dello strato spinoso (o malpighiano)
Mostrano i desmosomi che le uniscono e conferiscono
all’epidermide una elevata resistenza meccanica.
Le cellule basali provvedono al ricambio cellulare
Gli spazi fra cheratinociti consentono scambi metabolici
La cheratinizzazione delle cellule è dovuta alla sintesi
delle cheratine che formano dei filamenti
Aumenta verso gli strati più superficiali dove le cellule appaiono
appiattite.
Gli spazi intercellulari diminuiscono e aumenta la presenza di lipidi ad
azione impermeabilizzante
Cheratinizzazione, corneificazione
E’ un processo proprio dei tetrapodi che assume una grande importanza perchè
consente il definitivo distacco dei dall’ambiente acquatico; infatti la
cheratinizzazione dello strato corneo fornisce protezione contro la disidratazione e
l’essiccamento.
L’altezza dello strato corneo varia da zona a zona in uno stesso individuo.
Lo strato corneo può produrre, per un diverso grado di corneificazione, delle
specializzazioni diverse nelle varie regioni del corpo dei Vertebrati contro insulti
meccanici, per l’offesa e la difesa
La cheratinizzazione implica la sintesi di cheratine che si accumulano nelle cellul
più superficiali determinandone la morte. Il processo è particolarmente evidente a
partire dai Rettili.
Sono note α e β cheratine, proteine fibrose fibrillari entrambe insolubili, ma
le prime sono più elastiche e presenti in tutti i vertebrati e protegge dalla disidratazione,
mentre le seconde sono più rigide, offrono protezione meccanica e si trovano in
rettili e uccelli.
L’epidermide dei Vertebrati può essere più o meno corneificata
e lo strato corneo può produrre delle strutture che nell’insieme
si chiamano fanere
Negli Anfibi adulti lo strato corneo è sottile e di solito limitato ad un
solo strato di cellule; l’epidermide perciò resta piuttosto permeabile
e permette gli scambi osmotici; la presenza di ATPasi di membrana,
enzima che regola il trasporto attivo di ioni, testimonia questa funzione
Derivati cornei dell’epidermide sono:
Dentelli stomodeali ( dei Ciclostomi e delle larve di Anfibi)
Squame di Rettili, Uccelli (zampe) e alcuni Mammiferi
Verruche e callosità
Artigli, unghia, zoccoli
Corna, becchi, fanoni
Penne degli Uccelli
Peli dei Mammiferi
Filogeneticamente i primi derivati cornei sono le squame e quindi gli
artigli (e derivati), le corna, le penne e i peli.
Tutti questi annessi epidermici, detti anche fanere sono costituiti
fondamentalmente da cellule morte ricche di cheratina.
La formazione dei derivati cornei quali squame, penne e peli è sempre
associata alla formazione di una papilla dermica, sia come elemento
“induttore” che per la nutrizione delle cellule che costituiscono la struttura
cornea.
Le prime fasi di sviluppo delle squame e delle penne sono molto simili e
poiché gli Uccelli sono derivati dai Rettili si ritiene che le penne siano
omologhe e siano quindi derivate dalle squame dei Rettili.
Il pelo invece sembrerebbe essere un elemento di nuova acquisizione tipica
ed esclusiva dei Mammiferi, che si sarebbe formato primariamente fra le
squame dei Rettili loro progenitori, nei tratti di epidermide sottile interposta
fra le squame (caratteristica peraltro conservata nell’Armadillo).
Squame dei rettili
La cheratinizzazione della pelle dei rettili non è uniforme in quanto
sono evidenti aree di ipercheratinizzazione in corrispodenza della
formazione di squame epidermiche che possono essere embricate
nei Serpenti, a scudo nei Cheloni e Loricati, tubercolate nel
camaleonte
Nell’epidermide dei Rettili si alternano strati di cellule con cheratina
dei due tipi.
L’epidermide dei Rettili va incontro a rinnovi ciclici che comportano
o il distacco di frammenti di pelli o la perdita completa di una sorta
di “astuccio” corneo, la spoglia o exuvie.
Nei serpenti nella fase di rinnovamento si forma una seconda
generazione di strati cheratinizzati e quando questa è completa
si ha il distacco dell’epidermide della precedente generazione
Squame cornee sono presenti sulle zampe degli uccelli e in alcuni
Mammiferi, pangolino e armadillo o nella coda dei roditori
pangolino
Corneificazione della pelle sull’estremità delle ultime falangi
forma l’artiglio o l’unghia o lo zoccolo
L’organizzazione di base è dovuta alla presenza di una
lamina ungueale dura e la solea più molle, ma cambia l’aspetto finale
Corna sono tutte le protuberanze situate sulla testa essenzialmente
dei Mammiferi pari o impari, distinguibili in diversi tipi per la loro
struttura e organizzazione.
Non tutte le corna sono ad esclusiva composizione cornea.
Sono esclusivamente epidermiche le corna del rinoceronte, formato da
una sorta di concrezione di peli.
Al contrario le corna delle giraffe sono protuberanze ossee dell’osso
frontale rivestite da uno strato di epidermide con peli
Altri tipi di corna sono formate da
un nucleo osseo su cui prolifera l’
epidermide che forma una sorta di
astuccio permanente come nei
bovidi o caduco come nell’
Antilocapra.
Le corna “a palco” dei cervidi sono
di natura ossea con rivestimento
epidermico. Sono soggette a cicli
annuali, possono essere presenti
solo nei maschi o in entrambi i sessi
es renna
Altri derivati cornei sono i becchi in rettili, uccelli e mammiferi e i
i fanoni delle balene
Le penne, piume e filopiume sono derivati cornei esclusivi della
classe degli Uccelli che nel complesso costituidscono un rivestimento
protettivo e Isolante legato all’omeotermia, inoltre le penne permettono
il volo.
La formazione delle penne richiede il concorso di derma ed
epidermide, infatti una papilla dermica vascolarizzata si addenserà
sotto l’epidermide lì dove si formerà la penna, che infossandosi
forma la regione del colletto del follicolo della penna ad intensa
proliferazione cellulare.
L’abbozzo è rivestito da una guaina epidermica che si lacera
a completa formazione
Le penne si formano in corrispondenza di linee
ben precise sul corpo gli pterili
Il colletto è un ispessimento
dell’epidermide e rappresenta la zona
proliferativa della penna in
accrescimento.
Nella formazione della piuma, dal
colletto si formano delle colonne
parallele di cellule che si accrescono
e alla fine si distaccano a formare un
ciuffo di barbe, una per ogni colonna
di cellule.
Nella formazione delle penne invece
in una zona del colletto, che
corrisponde alla parte dorsale della
futura penna, si ha una maggior
proliferazione in forma di asta che
darà il rachide cui si saldano le
Barbe
Componenti della penna:
Calamo: infisso nel derma, cavo
Rachide, parte sporgente che porta
il vessillo
Vessillo formato da due serie di
barbe inserite sul rachide
Barbe che portano 2 file di barbule
di cui una provvista di amuli,
specie di ganci che si tengono ancorati
alle barbule delle serie adiacenti
per tenere il vessillo coeso.
Il “piumino” è formato da piume e
filopiume in cui è assente il
vessillo e le barbe sono direttamente
inserite sul calamo o sul rachide
Le penne principalmente
coinvolte nel volo sono
quelle delle ali, le
remiganti e quelle della
coda, le timoniere
Le penne sono soggette
a cambiamenti stagionali nei
colori e ad un ricambio di
solito collegati ai cicli
riproduttivi, grazie alla
permanenza di una papilla
ricoperta da epidermide che
riprende il ciclo di formazione
I peli sono i derivati cornei ad esclusiva presenza nei Mammiferi.
Sono associati ad una ghiandola sebacea e al muscolo erettore del pelo
il pelo si forma dal follicolo pilifero in cui il bulbo, epidermico,
circonda la papilla dermica;
la radice del pelo è la zona di corneificazione a contatto
con il follicolo
Il fusto è la parte sporgente del pelo formato da midolla, corteccia e
rivestito da una cuticola, si allunga per proliferazione delle cellule
del bulbo; la corteccia è la parte del fusto che può avere spessore
diverso in specie diverse, fino a formare degli aculei (riccio)
I peli possono variare nello spessore, distribuzione e possono
avere funzioni speciali come recettori tattili, le vibrisse.
Il colore dei peli, così come anche le penne è dato essenzialmente
dalla melanina, maggiore o minore presenza o melanine diverse
Inoltre nella loro formazione queste due strutture intrappolano
bolle d’aria, che influenzano il colore per effetti fisici.
Il ricambio dei peli è continuo o su base stagionale,
la loro crescita è limitata (>>peli, ciglia) o prolungata
(criniere, capelli..) e poi comunque c’è il ricambio.
Le ghiandole del tegumento
sono presenti in tutte le classi ma variano per tipo di secreto,
varietà e distribuzione.
Sono ghiandole esocrine e riversano il loro contenuto all’esterno
Le ghiandole esocrine - unicellulari o pluricellulari - derivano
dall’epidermide e vi restano sempre collegate mediante un dotto escretore.
In base alla forma dell’adenomero, ossia della parte secernente, si
distinguono in
alveolari se la forma è sferica,
tubulari se la forma e la cavità sono cilindriche
acinose se la forma dell’adenomero è sferica e la cavità cilindrica.
In base alla qualità del secreto possono essere classificate in ghiandole
mucose, sierose o miste.
In base alle modalità di secrezione le ghiandole esocrine vengono distinte in:
merocrine o eccrine se secernono i prodotti mantenendo la loro integrità;
apocrine, se con il secreto viene distrutta anche la parte apicale delle cellule
(es gh. mammaria);
olocrine, quando con la secrezione si ha il disfacimento anche delle cellule
(es. gh. sebacea).
Lo svuotamento dell’adenomero è sovente facilitato dalla contrazione di uno
strato di cellule mioepiteliali tra lo strato ghiandolare e la membrana basale.
Nella pelle dei pesci sono presenti numerose ghiandole unicellulari a
secreto mucoso o sieroso e alcune ghiandole pluricellulari.
La maggior parte produce muco le cui funzioni sono :
battericida, ridurre l’attrito con l’acqua
evitare variazioni osmotiche fra l’organismo e l’acqua.
Altri secreti possono avere funzione offensiva, velenosa o irritante
In pesci abissali, la pelle presenta dei fotofori che sono ghiandole
trasformate capaci di emettere un segnale luminoso;
possono presentare una specializzazione a mò di lente
e del pigmento intorno che isola il fotoforo dai tessuti circostantiLa
reazione che avviene nelle cellule dei fotofori spesso è ad opera
di batteri simbionti e genera luce per bioluminescenza
Nella pelle di Anfibi sono presenti ghiandole pluricellulari che affondano
nel derma e producono muco o secreto sieroso (peptidi o amine biogene)
ad azione tossica o antibatterica
I rettili hanno una pelle corneificata e presentano scarsissime
ghiandole spesso a secreto odoroso o localizzate in ristrette aree
come i pori femorali delle lucertole o in regioni pericloacali es nei
cheloni
Anche gli Uccelli presentano una limitata popolazione ghiandolare
a secrezione lipidica come la ghiandola dell’uropigio o ghiandole
ceruminose nel meato uditivo esterno
Nei Mammiferi ricompare una varietà di ghiandole con diversi tipi di
secreto e diverse funzioni:
ghiandole mammarie
ghiandole sebacee
ghiandole sudoripare
ghiandole odorifere
Le ghiandole mammarie si sviluppano lungo le linee del latte
che vanno dal cavo ascellare fino all’inguine;
il numero delle coppie di capezzoli che si formano è
proporzionale alla prole
I dotti delle gh. mammarie possono sboccare direttamente
all’esternoin corrispondenza di un capezzolo o in una cisterna del
latte, che è una piega cutanea
Tipologia delle ghiandole mammarie
in diversi mammiferi.
Nei monotremi le ghiandole del latte
sboccano in corrispondenza di un
ciuffetto di peli
Nei ruminanti è presente una cisterna
del latte dove sboccano i dotti galattori
Il differenziamento e proliferazione delle ghiandole mammarie
e produzione del latte sono sotto l’influenza di stimoli ormonali
Nei mammiferi sono presenti
ghiandole sebacee, sudoripare e ghiandole a secrezione
odorosa (feromoni).
Le ghiandole sebacee sono strettamente associate ai peli, le
ghiandole sudoripare sono diversamente distribuite sul corpo, con
maggior concentrazione dove scarseggiano i peli
La ghiandola sebacea produce un secreto lipidico che lubrifica e
rende la pelliccia idrorepellente
Le ghiandole sudoripare producono un secreto acquoso
e ricco di elettroliti con funzione soprattutto di termoregoalzione
I mammiferi posseggono numerose ghiandole odorifere, il
cui secreto può avere funzione di richiamo, o sevono per
segnare il territorio.
Sono per esempio localizzate in prossimità dei genitali o sulla testa
o in posizione ascellare.
I loro principi attivi venivano usati nella produzione dei profumi
Mineralizzazione del derma
Il derma è un tessuto connettivo che possiede antiche e persitenti potenzialità di
formare tessuto osseo: un esoscheletro superficiale più o meno sviluppato ad opera
di osteoblasti od odontoblasti.
La mineralizzazione implica la formazione di una trama collagenica e deposizione di
sali minerali (cristalli di idrossiapatite) che porta alla formazione di osso o dentina
La dentina è una variante di osso di consistenza più dura.
Altro tessuto fortemente mineralizzato è lo smalto, prodotto ad opera degli
adamantoblasti di origine ectodermica. E’ comunque sempre associato a dentina.
Osso, dentina e smalto comparivano tutti nelle antiche scaglie dermiche dei Pesci
corazzati; nel corso dell’evoluzione però il dermascheletro si alleggerisce e lo si trova
attualmente nei Pesci moderni per lo più in forma di piccole scaglie flessibili.
L’armatura dermica svolgeva un’azione di difesa e rappresentava una riserva di calcio
e fosfati.
Componenti dermiche mineralizzate nei tetrapodi
Scaglie dermiche in alcuni anfibi (es apodi)
Consistenti piastre dermiche in cheloni e coccodrilliani
Piastre dermiche degli armadilli
Le scaglie dermiche
dei Pesci:
lungo due linee
evolutive diverse si
sono formate a partire
dalla scaglia cosmoide
S. ganoidi
S. elasmoidi
S. placoidi
Scaglia cosmoide antica con diversi strati
ossei e rivestimento di smalto (scaglia più antica) la cui evoluzione
ha portato a forme più moderne di scaglie
Scaglie elasmoidi sono le più diffuse tra i pesci ossei attuali
Scaglie placoidi sono presenti nel tegumento dei Condroitti
La pigmentazione della pelle è principalmente dovuta alla presenza
di cellule specializzate: i cromatofori presenti nel derma e derivati
dalle cellule delle creste neurali, oltre che dalla vascolarizzazione.
I cromatofori hanno forma stellata e possono contenere diversi tipi
di pigmento. Nei mammiferi si trovano i melanociti
Funzioni della pigmentazione
Mimetismo/corteggiamento
Termoregolazione
Protezione
I cromatofori si possono distinguere in base ai tipi di pigmenti in:
1- melanofori/ melanociti, con granuli da rosso scuro/bruno
2- xantofori, con granuli di pigmento giallo
3- eritrofori, con granuli di pigmento rossi
4- iridofori o guanofori con microcristalli incolori di guanina
1, 2 e 3 danno un tipo di colorazione su base chimica e fisica;
il tipo 4 produce invece una colorazione su base fisica (strutturale)
Cambiamenti di colore
I cambiamenti di colore tipici dei Vertebrati eterotermi sono dovuti alla
morfologia e fisiologia dei melanofori ed anche alla combinazione di una
colorazione strutturale con una colorazione pigmentaria.
Il cambiamento di colore è determinato da
- controllo nervoso per azione dell’acetilcolina e dell’adrenalina
- e/o meccanismo ormonale per azione di ormoni dell’ipofisi dell’epifisi;
- vi può essere anche una combinazione dei due meccanismi.
Nei melanofori i granuli di melanina possono muoversi diffondendo
nei prolungamenti citoplasmatici o concentrandosi in un’area
ristretta attorno al nucleo.
Sono provocati da informazioni visive che raggiungono l’encefalo tramite
il nervo ottico e infatti, l’accecamento è seguito dalla perdita delle capacità
mimetiche dell’animale.
negli Uccelli e nei Mammiferi la melanina è sintetizzata nei melanociti
ma durante lo sviluppo di penne e peli i granuli di pigmento
vengono ceduti alle cellule all’epidermide e agli annessi cutanei (penne o
peli) in via di formazione
Ormoni specifici – melatonina (epifisi) intermedina (ipofisi) - e
l’adrenalina controllano sia la sintesi sia il grado di
distribuzione dei granuli nei cromatofori
I due gruppi di molecole agiscono con meccanismo antagonista
A- per concentrare i granuli nel corpo cellulare
B- per disperderli nei prolungamenti cellulari