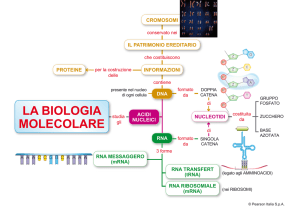
Le differenze morfologiche e funzionali delle cellule che compongono un individuo
riflettono l’esecuzione di definiti programmi genetici che, essenzialmente, fanno sì che
certi geni siano espressi in un tipo cellulare e non in un’altro.
L’espressione di un gene avviene attraverso due processi:
- la trascrizione del DNA in RNA
- la traduzione di questo in proteine.
Su questi processi intervengono numerosi altri meccanismi che in modo
integrato garantiscono la:
REGOLAZIONE DELL’ESPRESSIONE GENICA
DNA
TRANSCRIPTION
RNA
TRANSLATION
PROTEIN
So, what makes different cells different
from one another?
Different gene expression profiles
(sets of genes) when regulated in
a coordinated manner lead to dramatic
differentiation of cells.
Although all cells always have all genes
not all genes are activated in the same
way in all cells.
L’esempio dei geni
globinici che
vengono espressi
in tessuti e tempi
specifici durante lo
sviluppo
dell’individuo
Geni globinici
cromosoma 16
famiglia
genica
cromosoma 11
famiglia
genica
Genoma
Insieme delle informazioni genetiche che caratterizzano un organismo.
Trascrittoma
Insieme degli RNA messaggeri prodotti da una
determinata popolazione cellulare.
Per ogni tipo cellulare diverso sono espressi
all’incirca 10000 geni diversi.
Proteoma
Insieme delle proteine prodotte da una determinata
popolazione cellulare.
Scienze «OMICHE»:
“…in biologia cellulare e molecolare…. il suffisso -omiche è
utilizzato per formare nomi con significato di… indicare tutti i
costituenti analizzati collettivamente”
• Genomica
• Trascrittomica
• Proteomica
•
•
•
•
Farmacogenomica
nutrigenomica
Metabolomica
Etc etc
La regolazione dell’espressione genica negli eucarioti
avviene a più livelli
Il primo evento che accade durante l’espressione di un gene è la
TRASCRIZIONE
TIPI DI RNA TRASCRITTI
mRNA – RNA messaggero
rRNA – RNA ribosomale
tRNA – transfer RNA
snRNA – small nuclear RNA
snoRNA – small nucleolar RNA
miRNA Micro RNA
….RNA…
RNA POLIMERASI EUCARIOTICHE
RNA Polimerasi I - rRNA
RNA Polimerasi II - mRNA
RNA Polimerasi III - tRNA & other small RNA
RNAs in the Genome
Strachan and Read (2004) p.247 F9.4
~200 snoRNA
small nucleolar, over 100 types –
RNA modification and processing
~100 snRNA
small nuclear - involved in splicing
~200 miRNA
very small ~22bp , regulation
~175 28S,5.8S,5S
large cytosolic subunit
~175 18S
small mitochondrial subunit
~250 5S
large mitochondrial subunit
>500
tRNA
transfer RNA
>1500 Antisense RNA
20000-10000
> 1500 types
long non coding RNA
RNA POLIMERASI
RNA polimerasi eucariote
• RNA pol I sintetizza i rRNA 5.8s 28s e 18s (strutturali)
• RNA pol II sintetizza i mRNA (codificanti per proteine)
• RNA pol III sintetizza i tRNA ed altri piccoli RNA strutturali
• Tutte le RNA pol iniziano la trascrizione da un sito discreto (sito di inizio)
• Il sito di inizio della trascrizione è definito geneticamente da discrete sequenze
nucleotidiche che costituiscono il promotore
• La frequenza con cui la RNA pol inizia la trascrizione può essere regolata da
sequenze nucleotidiche anche molto distanti dal sito di inizio (enhancers e
silencers)
• Tutte le RNA pol terminano la trascrizione in siti discreti (terminatore)
TRASCRIZIONE NEGLI EUCARIOTI
RNA polimerasi
eucariote
PROMOTORE
DNA
gene
TERMINATORE
5’….CCATGTCCCCCACGGGGTAAATGGCTC……3’
3’.. GGTACAGGGGGTGCCCCATTTACCGAG……5’
DNA
5’….CCATGTCCCCCACGGGGTAAATGGCTC……3’
RNA
5’….CCAUGUCCCCCACGGGGUAAAUGGCUC…3’
PROMOTORE
DNA
gene
TERMINATORE
5’….CCATGTCCCCCACGGGGTAAATGGCTC……3’
3’.. GGTACAGGGGGTGCCCCATTTACCGAG……5’
5’….CCATGTCCCCCACGGGGTAAATGGCTC……3’
RNA
5’….CCAUGUCCCCCACGGGGUAAAUGGCUC…3’
Template3’.. GGTACAGGGGGTGCCCCATTTACCGAG……5’
senso 5’
anti-senso
senso
3’
anti-senso
3’
5’
La direzione di trascrizione è determinata dal promotore
La regolazione dell’espressione genica negli
eucarioti avviene a più livelli
Il controllo della trascrizione avviene a numerosi
livelli:
Pre-attivazione
(Induzione di uno stato
trascrizionalmente
competente)
Inizio/re-inizio della trascrizione
Elongazione del trascritto
Maturazione del trascritto
Terminazione del trascritto
Fattori di trascrizione legati a specifici
promotori/enhancers
Co-attivatori (acetilasi degli istoni,
attivita’ che rimodellano la cromatina)
Fattori “generali” di trascrizione (GTFs)
e RNA polimerasi
Gli elementi regolatori della trascrizione sono siti di
legame di specifiche proteine –
FATTORI DI TRASCRIZIONE -
- martedi 22 novembre BIOLOGIA al posto di statistica INIZIO ore
8.45
_mercoledi 23 NO BIOLOGIA ma FISICA
-
Mercoledi 30 novembre inversione ore chimica – biologia
-
1 dicembre NO biologia
-
venerdi 9 dicembre NO biologia ma statistica
- biologia il 15 dicembre ore 14.30 -17-30
- biologia al 23 dicembre. Ore 11.30 -14.30
La trascrizione dei geni eucarioti è controllata da numerose
proteine: FATTORI DI TRASCRIZIONE
La funzione principale dei fattori di trascrizione e’ di facilitare
l’assemblaggio del macchinario basale di trascrizione sul promotore essenziale
(core).
A livello trascrizionale, la regolazione si esercita sulla
frequenza con cui un gene è trascritto.
Le sequenze che segnalano il sito di inizio della trascrizione di un gene sono dette promotori
Figure 6-17 Molecular Biology of the Cell (© Garland Science 2008)
Il core dei promotori riconosciuti dalla RNA pol II è costituito da una
combinazione di diversi elementi:
- BRE (TFIIB recognition element)
- TATA (TATA box) (25 nt a monte dall’inizio della trascrizione)
- Inr (elemento iniziatore)
- DPE (Downstream promotoer element)
- MTE (motif 10 element)
Altre seq importanti in vivo:
- Altri elementi del core del promotore
- UAS (upstream activator sequences)
- enhancer
- silencers
- boundary elements
-Insulators
Tutte queste seq di DNA legano proteine regolatorie specifiche. Alcune di queste seq possono essere localizzate a
grande diatanza (x10-x100 bp) dal promotore
The specific combination of promoter and enhancers of a gene
determine its transcriptional activity.
The activity or presence of these factors in a given cell determines
whether a particular gene is on or off.
Enhancer
Promoter (general transcription factors)
intron
intron
Exon 1
Exon 2
Enhancer
Exon 3
RNA polymerase just beyond the initiation site.
“Specific” transcription factors are required
in order to activate RNA polymerase.
Transcription
termination
L’inizio della trascrizione da parte dell’RNA Pol II
richiede multipli fattori
Fattori di trascrizione generali (GTF) insieme con l’RNA Pol II
formano l’apparato trascrizionale basale, che lega il promotore
essenziale ed è sufficiente alla trascrizione in vitro
TBP (che con i fattori associati a TBP (TAF) forma -> TFIID)
TFIIA, TFIIB, TFIIF, TFIIE, TFIIH
Fattori di trascrizione specifici, anche richiesti per la trascrizione
attivata: servono per reclutare e assemblare l’apparato trascrizionale
– Si legano a elementi di riconoscimento sul promotore e sull’enhancer
– Multipli fattori di trascrizione sono normalmente coinvolti nell’attivazione di un
gene
Il controllo della trascrizione è basato sul riconoscimento di corte
sequenze di DNA da parte di diverse classi di proteine
Il core dei promotori riconosciuti dalla RNA pol II è costituito da una
combinazione di diversi elementi:
- BRE (TFIIB recognition element)
- TATA (TATA box) (25 nt a monte dall’inizio della trascrizione)
- Inr (elemento iniziatore)
- DPE (Downstream promotoer element)
- MTE (motif 10 element)
Altre seq importanti in vivo:
- Altri elementi del core del promotore
- UAS (upstream activator sequences)
- enhancer
- silencers
- boundary elements
-Insulators
Tutte queste seq di DNA legano proteine regolatorie specifiche. Alcune di queste seq possono essere localizzate a
grande diatanza (x10-x100 bp) dal promotore
L’elemento TATA-box è molto spesso
presente nei promotori eucarioti
Figure 6-16 Molecular Biology of the Cell (© Garland Science 2008)
Assemblaggio
del complesso
d’inizio della
trascrizione
negli eucarioti
TBP
TFIIB
Fattori di trascrizione eucariotici
TATA binding protein (TBP) component of TFIID binds to TATA
box
Distorts (kinks and unwinds) DNA upon binding, provides platform
for other TFs to bind
Struttura del dominio C-terminale
di TBP legato alla TATA-box
TBP
TFIIB
Struttura del complesso tra
TBP, DNA e TFIIB
TFIIE
TFIIH
ELICASI
TFIIE: DNA melting al
promotore
TFIIH: 9 subunità
DNA elicasi per
esporre il frammento
stampo
Fosforilazione della coda
(CTD) della RNA Pol II:
necessaria per il distacco
della RNA Pol II dal
complesso di inziio della
trascrizione
La trascrizione della maggior parte dei geni eucarioti è regolata da
multipli elementi
L’attività della RNA pol II è spesso influenzata da elementi anche molto
distanti dal sito di inizio (enhancers o silencers)
Gene Umano Tipo
inizio e direzione
di trascrizione
elementi distali (enhancer o silencer)
elementi prossimali
TATA box
esone
introne
Altre proteine sono necessarie perché la trascrizione avvenga in vivo
Modello di regolazione trascrizionale
L’inzio della trascrizione del gene Pax6 è diversa a seconda
dei diversi tessuti in cui è espresso
Il gene di Pax6 possiede 3 diversi siti di inizio
della trascrizione, che vengono attivati in
diversi tessuti in circostanze specifiche.
Interazione tra fattori trascrizionali
e DNA
Gal4
(zinc-finger)
Gcn4
(leucine-zipper)
Max
(helix-loop-helix)
Transcription factors
Prokaryotes
Eukaryotes
RNA eucariote vs procariote: 5’ capping, RNA splicing, 3’ polyadenilation
Poly-cistronic mRNA
Mono-cistronic mRNA
3’-poly(A) tail
5’-cap
Figure 6-22a Molecular Biology of the Cell (© Garland Science 2008)
07_20_Pro_v_Eucar.jpg
Maturazione dei RNA (RNA processing)
•
capping
•
poliadenilazione
•
splicing
•
editing
Regolazione
post-trascrizionale
La particolare organizzazione delle
sequenze codificanti dei geni umani
favorisce meccanismi di regolazione
post-trascrizionale dell’espressione
genica.
Maturazione del mRNA
AAUAAA
AAAAAA 200-300
AAAAAA 200-300
mRNA processing: il cappuccio
- Il CTD di PolII si trova adiacente
al solco di uscita del pre-mRNA
- Il capping comincia quando ~ 20-30
nucl. sono stati sintetizzati ad opera di
3 enzimi che agiscono in modo coordinato.
5’-cap structure
Il 5-cap è aggiunto al nascente mRNA
FOSFATASI
Guaniltransferasi
metiltransferasi
- Il cappuccio stabilizza l’mRNA (no degradaz. da esonucleasi).
- Legato a elF-4E e altri fattori di iniziazione promuove l’impegno delle
subunita’ ribosomali con l’mRNA (interazione con PABP1-circolarizzazione)
Terminazione della Trascrizione
•I meccanismi di terminazione della trascrizione sono diversi e dipendono
dal tipo di RNA polimersi
•L’attività di RNA pol I è terminata da fattori pol I specifici che legano
regioni 3’ dell’unità trascrizionale
•L’attività di RNA pol II termina in una regione di 0.5-2 kb oltre il sito di
poliadenilazione, e la terminazione è accoppiata al processo che taglia
e poliadenila l’estremità 3’ del trascritto
•L’attività di RNA pol III termina dopo la sintesi di una serie di U
Poliadenilazione e terminazione del mRNA nascente
mRNA processing: formazione dell’estremita’ 3’
Il taglio del pre-mRNA avviene dopo la
sequenza AAUAAA
07_20_Pro_v_Eucar.jpg
Maturazione del mRNA
AAUAAA
AAAAAA 200-300
AAAAAA 200-300
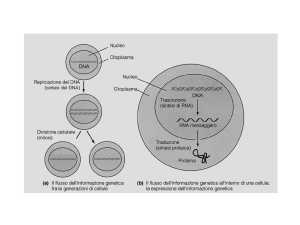
Genetic information is transferred from genes to the proteins they
encode via a “messenger” RNA intermediate
DNA
GENE
transcription
messenger RNA
(mRNA)
translation
protein
Splicing
Alcuni geni hanno la loro informazione codificante per proteina
interrotta da sequenze non codificanti.
Le seq. codificanti si chiamano ESONI
le seq non codificanti INTRONI
DNA
esone 1
GE
introne
esone 2
NE
transcription
precursor-mRNA
(pre-mRNA)
introne
Gli introni devono essere rimossi mediante un processo
chiamato “RNA splicing”
Thus expression of a gene with an intron requires an extra
step to remove the intron
exon 1
DNA
GE
intron
exon 2
NE
transcription
pre-mRNA
intron
RNA splicing
mRNA
translation
protein
Prokaryotic gene
Eukaryotic gene
Regioni Codificanti
(ESONI)
Regioni Non-codificanti
(INTRONI)
Pre-messenger RNA Processing
pre-mRNA
M7G
exon
exon
intron
cap
mRNA
AAAAAAA200
RNA splicing
M7G
nucleus
AAAAAAA200
transport
cytoplasm
M7G
AAAAAAA200
ribosomes
protein
poly(A) tail
In humans, many genes contain multiple introns
intron 1
1
intron 3
intron 2
2
3
1
2
3
intron 4
4
4
5
Usually all introns must be removed before the mRNA can
be translated to produce protein
5
SPLICING
Figure 6-25 Molecular Biology of the Cell (© Garland Science 2008)
A typical mammalian gene and an especially large one.
SUR2 gene:
exon
9.6 kb intron
intron
40 kb
Human Dystrophin gene
260 kb intron
2.4 Mb
Questo gene è lungo 2.4 MILIONI di basi
Numero di introni/gene nelle diverse specie eucariotiche
Genes and Gene Structures I
•Presently estimated Gene Number: 24.000 (reference: )
•Average Gene Size: 27 kb
•The largest gene: Dystrophin 2.4 Mb - 0.6% coding – 16 hours to transcribe.
•The shortest gene: tRNATYR 100% coding
•Largest exon: ApoB exon 26 is 7.6 kb
Smallest: <10bp
•Average exon number: 9
•Largest exon number: Titin 363
Smallest: 1
•Largest intron: WWOX intron 8 is 800 kb
Smallest: 10s of bp
•Largest polypeptide: Titin 38.138 smallest: tens – small hormones.
•Intronless Genes: mitochondrial genes, many RNA genes, Interferons, Histones,..
Gli esoni codificano domini proteici
Pre-messenger RNA Processing
pre-mRNA
M7G
exon
exon
intron
cap
mRNA
AAAAAAA200
RNA splicing
M7G
nucleus
AAAAAAA200
transport
cytoplasm
M7G
AAAAAAA200
ribosomes
protein
poly(A) tail
In humans, many genes contain multiple introns
intron 1
1
intron 3
intron 2
2
3
1
2
3
intron 4
4
4
5
Usually all introns must be removed before the
mRNA can be translated to produce protein
5
However, multiple introns may be spliced differently
in different circumstances, for example in different
tissues.
Heart muscle
1
1
2
2
Uterine muscle
5
3
3
1
3
4
4
5
5
Thus one gene can encode more than one protein. The proteins are similar but
not identical and may have distinct properties. This is important in complex
organisms
Gli esoni codificano domini proteici
Figure 6-27 Molecular Biology of the Cell (© Garland Science 2008)
Splicing occurs in a “spliceosome”
an RNA-protein complex
spliceosome
(~200 proteins + 5 small RNAs)
pre-mRNA
spliced mRNA
Lo spliceosoma è un complesso
ribonucleoproteico
SPLICING
SPLICEOSOME
Lo Splicing avviene tra sequenze conservate
Sequenze Consenso di splicing al 5 e 3 degli introni
Sequences within the intron itself provide important information for proper splicing
to take place. These consensus sequences is all that is necessary for an intron
to be processed.
The internal intron sequences can be anything
Sequenze di consenso attorno ai siti di splicing nei pre-mRNA dei vertebrati
Lo splicing richiede:
-Il riconoscimento di uno splice site al 5’
- Un residuo di Adenina (punto di ramificazione) , dove
avviene una ramificazione del la struttira a cappio del RNA.
A è a 20-50bp dall sito di splicing al 3’,
- Regione ricca di pirimidine (15bp), sito di legame per le
hnRNP di tipo A1, C e D (domini RRM)
Lo Splicing comporta due reazioni
sequenziali di trans-esterficazione
How does splicing work?
RNA Splicing is the elimination of
intron and/or exons from the primary
transcript in order to produce an
RNA molecule that goes on to code
for a protein.
How does splicing work?
RNA Splicing is a precise
biochemical process that is
guided by specific sequences
within the primary transcript
as well as other RNA molecules
within the “splicesome”
ribonucleoprotein (RNP)
enzyme.
Splicing is strictly regulated
(this is not a random process).
How does splicing work?
1.A specific adenine in the intron
attacks the 5’-splice site, using its
2’-OH as a “knife”, and cuts the RNA
backbone.
2.This results the 5’-end of the transcript physically detached from the
rest of the RNA molecule.
The 3’-end has a loop where the
2’-OH of the attacking adenine is
now covalently linked to the 5’-end
of the intron. This is the branch point.
How does splicing work?
3.The free 3’-OH of the exon
then attacks at the precise start
of the next exon attaching itself
to the RNA backbone.
This displaces the intron and results
in a contiguous exon/exon joint. The
intron is conatined in lariat structure
that then is discarded.
Le reazioni di splicing sono assistite da
piccoli RNA nucleari ricchi di uridine (snRNAs)
The splicesome catalyzes the RNA
splicing reaction.
The splicesome is a multi-subunit
complex of ribo- nucleo proteins (RNPs)
that are usually small and nuclear (sn)
and thus called snRNPs.
The splicesome contains both proteins
and RNA molecules. This is why it is
called a RNP complex (like the ribosome).
Different parts of the splicesome are
called U1, U2, etc. and each contain
protein and RNA.
1.U2Af and the branch binding
protein (BBP) first bind the
adenine in the intron.
2.Then, the rest of U2 assembles
displacing U2AF and BBP
and U1 assembles on the 5’-end
of the intron.
3.Other snRNPs bind and bring
the two ends of the intron
together and catalyze cleavage of
the 5’-end of the intron and
lariat formation.
4.U1 and U4 come off and U6
catalyzes the 3’-OH of the exon
attack on the 5’-end of the
following exon.
5. this results in joining of the two
exons and excission of the lariat
structure that was the intron.
The RNA components of the snRNPs are critical for binding specificity, and for guiding
the snRNPs to the correct positions on the transcript by virtue of the H-bonds formed by
basepairing snRNP RNA and transcript RNA.
RNA/RNA basepairing between
different snRNP subunits also
occurs in order to form
“docking sites” with particular
structures that help bring distant
intron sequences next to one another.
After the initial cleavage,
U5 holds on to exon #1,
and helps catalyze the next
step of 3’-OH of the exon
joining to the 5’-end of
exon #2.
all this requires energy from
ATP.
Formation of RNA/RNA basepairing
and breaking of these basepairs is
critical for the formation of specific
structures within the splicesome.
all this requires energy from
ATP, and RNA helicases are one
set of enzymes that uses ATP to
“melt” RNA/RNA H-bonds
during all these structural changes.
RNA/RNA basepairing between
different snRNP subunits and
between snRNP and pre-mRNA
form the structures that are largely
responsible for most of the catalytic
activity of the splicesome.
Il ciclo dello
spliceosoma
As mRNA is synthesized SR proteins (rich in serines & arginines) coat the exons as
they come off the RNA polymerase. U1 & U2 also mark the boundaries of where the
introns start and end.
This increases the accuracy of splicing.
Because some introns can be very long they can be packaged into
hnRNP complexes (like DNA is packaged into chromatin). Exons
can also be packaged into hnRNPs.
The splicing chemistry does not begin until after the assembly of all the initial
components is completed. This delay can allow splicing out of introns & exons in any
order, not just the order they happen to be synthesized.
La maturazione dei RNA avviene in un
limitato numero di localizzazioni
nucleari
DNA: blu
RNA Poliadenilati : rosso
Proteine spliceosome: green
RNA Poliadenilati : rosso
Splicing alternativo
Splicing alternativo
Splicing alternativo
Mutazioni che interessano il meccanismo di Splicing
DIFETTI DI SPLICING E MALATTIE EREDITARIE
Cooper et al.
Cell 2009
TIPI DI RNA TRASCRITTI
mRNA – RNA messaggero
rRNA – RNA ribosomale
tRNA – transfer RNA
snRNA – small nuclear RNA
snoRNA – small nucleolar RNA
miRNA Micro RNA
RNA POLIMERASI EUCARIOTICHE
RNA Polimerasi I - rRNA
RNA Polimerasi II - mRNA
RNA Polimerasi III - tRNA & other small RNA
la RNA pol I
Nel genoma i geni per i rRNA sono centinaia, organizzati in copie multiple
che vengono trascritte simultaneamente in una particolare zona del nucleo
chiamata nucleolo
Anche i RNA strutturali vanno incontro a maturazione
Organizzazione dei geni ribosomali
Maturazione dei rRNA
I geni per rRNAs funzionano da
organizzatori nucleolari
Piccoli RNAs nucleolari (snoRNAs) assistono la maturazione
dei rRNAs e l’assemblaggio delle subunità ribosomali
TIPI DI RNA TRASCRITTI
mRNA – RNA messaggero
rRNA – RNA ribosomale
tRNA – transfer RNA
snRNA – small nuclear RNA
snoRNA – small nucleolar RNA
miRNA Micro RNA
RNA POLIMERASI EUCARIOTICHE
RNA Polimerasi I - rRNA
RNA Polimerasi II - mRNA
RNA Polimerasi III - tRNA & other small RNA
Tutti i pre-tRNAs subiscono maturazione: taglio,
splicing e modificazione di basi
Lo Splicing dei pre-tRNAs differisce dagli altri
meccanismi di splicing
La diversa stabilità dei mRNA contribuisce alla
regolazione dell’espressione genica