Meristemi e differenziamento
Diversamente dagli animali, le piante hanno regioni di
crescita continua, con cellule in continua divisione
cellulare: MERISTEMI.
Le nuove cellule prodotte dalle cellule meristematiche sono
piccole, prive di vacuoli o con vacuoli molto piccoli.
Con la maturazione, le cellule assumono forme e dimensioni
in relazione alla specifica funzione cellulare e i vacuoli
aumentano di dimensioni.
MERISTEMI APICALI – Si trovano agli apici di radici e
germogli. Determinano la CRESCITA IN LUNGHEZZA della
pianta (CRESCITA PRIMARIA).
Meristema
apicale
caulinare
Meristema apicale radicale
La pianta attua due tipi di crescita:
1) Accrescimento in altezza (crescita primaria di
fusto, radici e foglie) (eredità del pattern
longitudinale definito nell’embrione)
2) Accrescimento primario in spessore (eredità del
pattern radiale definito nell’embrione), seguito
nelle specie arboree di gimnosperme ed
angiosperme
dicotiledoni
da
accrescimento
secondario
Entrambi i tipi di crescita primaria sono a carico
di meristemi e prevedono divisione, crescita e
differenziamento cellulare.
La definizione dei meristemi caulinare e
radicale avviene durante l’embriogenesi
zigotica (v.lezione sull’embrione zigotico)
I MERISTEMI
sono regioni costituite da cellule
perennemente giovani che conservano la capacità di
dividersi per moltissimo tempo, in molti casi per tutta
la vita della pianta.
Sono costituiti da un gruppo eterogeneo di cellule,
oltre ai primari di diretta derivazione embrionale,
altri meristemi si formano in momenti e posizioni
diverse durante lo sviluppo post-embrionale della
pianta ed hanno anche potenzialità differenziative
diverse.
Per meristema vero e proprio si intende una
popolazione di cellule organizzate in cui le divisioni
avvengono, ed in modo ordinato.
Meristemi postembrionali: gemme
ascellari apici di
radici laterali
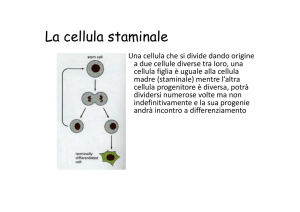
Una cellula meristematica è una cellula in grado di
proliferare dando una progenie
Caratteristiche citologiche delle cellule dei meristemi.
hanno nuclei grandi
hanno parete cellulare primaria e molto sottile
mancano di spazi intercellulari
la forma e le dimensioni di queste cellule sono variabili,
tuttavia si possono considerare più o meno isodiametriche
presentano un considerevole numero di plasmodesmi
sono assenti i grandi vacuoli, ma in alcuni meristemi
possono essere presenti piccoli vacuoli dispersi nel
citoplasma. Nelle gimnosperme sono frequenti meristemi
con cellule vacuolate.
Le cellule del cambio subero-fellodermico
(meristema
secondario)
presentano
spesso
cloroplasti, le iniziali del cambio cribrovascolare
(meristema
secondario)
possono
presentare amido e tannini, infine nei meristemi
di alcuni embrioni maturi possono essere
presenti sostanze di riserva.
Quali sono le cellule che compongono i meristemi?
Le cellule meristematiche si distinguono in
cellule fondatrici, cellule iniziali e cellule
derivate.
Questa suddivisione si basa sulla frequenza
di divisione e sulla posizione che occupano
all’interno di un meristema.
Le cellule fondatrici sono situate nella regione
centrale di un meristema apicale, ad es. le cellule
del centro quiescente (CQ) della radice.
Si dividono lentamente e controllano il destino
delle cellule circostanti (cellule iniziali).
Le cellule iniziali sono cellule meristematiche che
danno origine alle cellule derivate. Queste ultime
si dividono rapidamente e formano i tessuti
meristematici da cui differenziano i tessuti adulti.
Le cellule fondatrici e le cellule iniziali formano la
nicchia staminale
Una cellula iniziale è capace di dividersi in
modo da formare una cellula meristematica
iniziale e una meristematica derivata, è cioè
capace di divisione asimmetrica.
Le cellule derivate continuano a dividersi distanziandosi
della popolazione delle cellule iniziali e dando vita ai
meristemi primari:
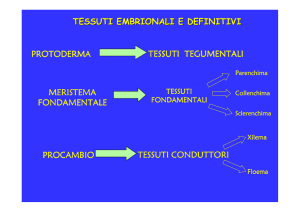
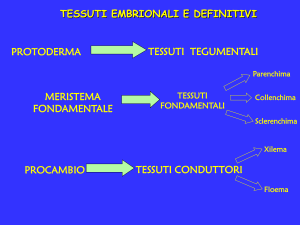
Protoderma
Meristema fondamentale
Procambio
Il termine staminale significa “filo conduttore” o
principio germinale costitutivo di organismi viventi.
Coerentemente con la loro etimologia le cellule
capostipiti di una
discendenza cellulare, sono staminali
le cellule fondatrici e le iniziali dei
meristemi.
staminali
sono
le
Le cellule staminali animali, un
esempio
• Le cellule staminali mesenchimali (MSCs)
sono cellule immature con la capacità di
autorinnovarsi e differenziarsi
continuamente in cellule specializzate
tessuto specifiche. Sono cellule di origine
midollare ma si trovano anche nel tessuto
adiposo, nel sangue periferico, nel cordone
ombelicale, nel derma, nel pancreas, nel
fegato, nel polmone ed in altri tessuti fetali.
Sia il meristema caulinare che quello radicale e le cellule
cambiali contengono cellule staminali che si dividono in modo
simmetrico o asimmetrico generando una cellula figlia che
andrà incontro a differenziamento ed una cellula che rimane
staminale, l’instaurarsi di un equilibrio dinamico tra divisione
cellulare e differenziamento è alla base del mantenimento
dell’attività di un meristema.
Divisione
cellulare
Differenziamento
Centri di organizzazione staminale (cellule
fondatrici) nei meristemi apicali di fusto e radice
Dal Libro di testo:
Elementi di
Biologia dello
Sviluppo delle
Piante, ed. EdiSES
Negli animali queste cellule vengono prodotte
durante
l’embriogenesi
e
rimangono,
anche
nell’individuo adulto, in vari organi.
Le cellule staminali, sia negli animali che nelle piante
sono cellule indifferenziate, in grado di dare origine
ad un’altra cellula staminale e a cellule che
andranno incontro al differenziamento.
Le cellule staminali animali sono multipotenti,
le cellule staminali vegetali sono multipotenti, ma
quelle dell’embrione fino a che non si separano i poli
caulinare e radicale sono totipotenti.
Per totipotenza si intende la capacità di una cellula di
dare origine a tutti i tipi cellulari possibili.
Es. lo zigote è totipotente perchè è in grado
differenziare tutte le cellule di un individuo adulto
(sia animale che vegetale).
Negli animali questa caratteristica si riduce nel
tempo. Infatti,le cellule staminali embrionali, durante
lo sviluppo, perdono la loro totipotenza e in un
animale adulto queste cellule sono in grado di
differenziare uno o pochissimi tipi cellulari.
Nei vegetali, non solo le cellule staminali sono in
grado di modificare il loro destino, ma anche le
cellule già differenziate,tuttavia per le prime è più
facile.
E’ importante notare che a differenza delle cellule
nelle piante una cellula
differenziata può dare origine anche ad
una cellula staminale.
staminali animali,
Nicchia delle cellule staminali
Nell’apice radicale di
Arabidopsis sono ben
distinguibili
nel
meristema le cellule
iniziali, le fondatrici
e le derivate.
Dal Libro di
testo: Elementi di
Biologia dello
Sviluppo delle
Piante, ed.
EdiSES
Cellule fondatrici
Le cellule staminali, come detto, sono considerate
precursori di cloni di cellule che possono rimanere
staminali o differenziare.
E’ stato dimostrato, per le cellule staminali, sia
animali che vegetali, che il loro stato è mantenuto
da
segnali
che
provengono
dall’ambiente
circostante.
Il microambiente dove sono presenti queste cellule
è detto nicchia delle cellule staminali.
Quest’ambiente
anche le cellule
che inducono il
cellula staminale
comprende oltre alle staminali
circostanti che producono i segnali
mantenimento della condizione di
(informazioni posizionali).
Le 4 cellule del centro quiescente dell’apice
radicale di Arabidopsis thaliana sono le cellule
fondatrici, le cellule circostanti sono le iniziali,
caratterizzate da una regolare disposizione
spaziale e le loro divisioni danno origine a tutte
le cellule derivate dei
tessuti primari della
radice.
La zonazione del doma
meristematico del germoglio
Nel doma apicale si individua una
zona centrale (ZC) in cui le cellule
sono relativamente grandi, vacuolate
e si dividono lentamente ed una zona
periferica (ZP) che circonda la ZC ai
lati. Nella ZP le cellule sono più
piccole ed hanno un’intensa attività
mitotica.
Nella ZP le cellule (derivate) iniziano a
differenziare in quanto vengono
canalizzate verso la formazione dei
primordi fogliari, delle gemme
ascellari o verso il differenziamento
dei tessuti più esterni del fusto.
Cellule
staminali
V. libro di
testo
Elementi
di Biologia
dello
sviluppo
delle
piante.
EdiSES
L’uscita dalla ZC segna l’inizio del differenziamento,
infatti le cellule della zona periferica sono incanalate
verso il differenziamento delle bozze fogliari o dei
primordi dei rami.
I confini della nicchia staminale sono determinati
dall’espressione di particolari geni. Ad es. nel caso
dell’apice di A.thaliana, nella nicchia staminale è
espresso il gene WUSCHEL (WUS). Nelle cellule
immediatamente fuori si esprimono geni appartenenti
alla famiglia genica CLAVATA.
Quindi
una
serie
di
informazioni
impartite
dall’attivazione di particolari geni determina l’esatta
localizzazione e dimensione di queste regioni cellulari.
Pattern di espressione di alcuni geni nel meristema apicale
di Arabidopsis.
STM mantiene la proliferazione cellulare nella zona
periferica svolgendo un ruolo complementare a quello di
WUSCHEL.
CLAVATA 3 regola negativamente l’espressione di
WUSCHEL
WUSCHEL regola positivamente l’espressione di
CLAVATA 3
SHOOTMERISTEMLESS (STM):
STM è espresso specificamente nelle cellule che
rimangono meristematiche.
STM inibisce il processo di differenziamento assicurando
che le cellule meristematiche rimangano indifferenziate.
STM è necessario anche per il mantenimento dell’identità
meristematica delle cellule nella pianta adulta.
Come è stato dimostrato?
Mutanti stm omozigoti, loss of function, non formano
meristema apicale e le cellule si differenziano
precocemente.
Mediante esperimenti di microchirugia è stato dimostrato che
le cellule della ZC (staminali) conferiscono al germoglio
un’elevata capacità rigenerativa.
Se
l’apice
viene
sezionato in due parti in
senso longitudinale, le
due
metà
formano
ognuna
un
meristema
intero.
Un
apice
sezionato in più parti e
in grado di formare
altrettanti
nuovi
meristemi.
In altri casi, se l’apice viene forato con aghi in modo da distruggere
alcune cellule del doma in posizione centrale, questo formerà uno o più
apici a partire dalle zone periferiche non danneggiate. Ciò sta a
dimostrare il ruolo delle cellule della ZC: la loro distruzione induce un
processo che porta alla loro sostituzione da parte di cellule derivate che
acquisiscono funzione staminale.
Dal Libro di testo: Elementi di Biologia dello
Sviluppo delle Piante, ed. EdiSES
Durante il normale sviluppo della pianta
Le cellule derivate si dividono vicino all’apice
vegetativo o radicale prima di andare incontro a
differenziamento. Tuttavia le divisioni cellulari
non sono ristrette esclusivamente alle cellule
derivate, bensì, continuano a dividersi anche le
cellule dei tessuti
che da loro prendono
origine.
Tali tessuti sono i tessuti meristematici primari,
sono parzialmente differenziati e rimangono in
questo stato per un certo periodo di
tempo
prima di dare origine alle cellule definitive dei
tessuti primari.
Il vantaggio, per una pianta, di avere i
meristemi localizzati in aree ben precise, separati
spazialmente dalle cellule mature, in cui i processi
fisiologici e metabolici sono attivi, è che può
crescere continuamente e contemporaneamente
essere in piena attività metabolica.
La crescita delle piante è per questo definita
indeterminata.
La crescita secondaria di una pianta è dovuta
all’attività di altri meristemi: cambio vascolare e
cambio subero-fellodermico. Questi meristemi sono
detti meristemi laterali secondari e sono localizzati
alla periferia del fusto e della radice.
Meristemi secondari/laterali
Cambio vascolare
Periciclo
Cambio del sughero
Floema secondario
Xilema secondario
Sughero
Felloderma
Meristemi secondari: si formano durante lo sviluppo
postembrionale
Meristemi ascellari: si formano alle ascelle delle foglie;
derivano dal meristema apicale
Meristemi delle radici laterali: si formano dalle cellule del
periciclo della radice primaria
Meristemi delle radici avventizie: si formano da cellule di
tessuti diversi dal periciclo della radice primaria (v. anche
meristemi avventizi)
Cambio vascolare: iniziali fusiformi e iniziali dei raggi;
xilema e floema secondario; raggi di tessuto parenchimatico
Cambio subero fellodermico: si sviluppa nelle cellule mature
del cortex e del floema secondario; si differenziano in cellule
del sughero che formano il periderma.
Meristemi intercalari: si trovano alla base degli
organi; graminacee
Un’ulteriore tipologia di meristemi è rappresentata dai
meristemi infiorescenziali e fiorali (v. Lezione transizione
a fiore e Lezione sul fiore)
Dove studiarli?
In piante modello
• tabacco
• arabidopsis
Un’altra tipologia di meristemi è rappresentata dai
meristemi avventizi.
Questi si formano per sdifferenziamento di tessuti
precedentemente differenziati in modo diverso.
Sono presenti nella pianta, es. i meristemoidi che
danno origine alle cellule di guardia degli stomi e ai
tricomi.
Ma i meristemoidi si organizzano anche in coltura in
vitro e la loro formazione precede sempre quella degli
organi avventizi (radici, gemme vegetative o fiori,
oppure di noduli vascolari, detta xilogenesi) (v.
Lezione
Caulogenesi
Radicazione avventizia)
Sperimentale
e
Lezione
Le cellule di guardia degli stomi si
originano per transdifferenziamento
Il meristemoide è una cellula epidermica che prima si divide per divisione diseguale
(cioè con separazione del destino fra le due cellule figlie) e poi per divisione uguale
(cioè con formazione delle due cellule di guardia, a stesso destino)
(trasdifferenziamento diretto) (dal Libro di testo)
Meristemi Avventizi
I meristemi avventizi si formano per
sdifferenziamento di tessuti parenchimatici, in
particolari condizioni. In seguito a ferite o in
colture in vitro.
Torniamo all’apice radicale ed a
quello caulinare
La stabilità di tutto il meristema si raggiunge
grazie ad un complesso dialogo molecolare fra le
cellule.
Nel meristema caulinare esiste un sistema di
segnalazione a breve distanza che controlla le
dimensioni ed i confini della nicchia delle cellule
staminali.
Nell’apice radicale è stata dimostrata sia l’esistenza
di una rete di segnali a breve distanza sia un sistema
di segnalazione a lunga distanza entrambi cooperano
per mantenere costante la forma e l’attività del
meristema stesso.
Nella
radice
il
segnale
operato
dal
dell’auxina che
radicale.
a
lunga
trasporto
è
polare
distanza
dal germoglio arriva all’apice
Quello a breve distanza va dalle cellule del centro
quiescente a quelle circostanti e viceversa.
Domande:
•
•
•
•
•
•
•
•
•
•
•
Quali sono i tipi di crescita della pianta, e come si chiamano i tessuti che li
garantiscono?
In cosa la crescita primaria è diversa dalla secondaria?
Quali sono le caratteristiche citologiche delle cellule meristematiche?
Le cellule di un meristema sono tutte con uguale funzione?
Quali sono i tipi cellulari che formano un meristema ed in che rapporto
reciproco sono fra loro?
Quali sono le cellule staminali di un meristema? E perché si chiamano così?
Come è fatta la nicchia staminale dell’apice del germoglio?
E quella dell’apice radicale?
Che tipo di segnali governano la nicchia staminale?
Che cos’è un meristema ascellare? Ed un meristema laterale?
Cosa si intende per meristema avventizio?
Dopo la germinazione il SAM dà luogo agli organi laterali.
SAM mantiene una struttura organizzata pur
rispondendo a segnali interni ed esterni di sviluppo.
A questo scopo al centro del SAM viene mantenuta una
popolazione di cellule indifferenziate che si dividono
lentamente.
Cellule che lasciano questa zona entrano in quelle
periferiche e si differenziano a formare gli organi laterali;
oppure nella zona sottostante (rib zone) e si differenziano
in cellule del fusto.
La velocità di proliferazione delle cellule iniziali nel SAM
deve essere coordinata con la velocità di differenziamento
delle cellule figlie.
• L’attività dell’apice caulinare
Utilizzando traccianti fluorescenti è stato visto che le cellule
della ZC sono collegate fra di loro da una fitta rete di
plasmodesmi, così come le cellule delle ZP, al contrario
sembra siano scarsi i collegamenti fra le diverse zone.
Questo fa ipotizzare che comunicazioni cellulari selettive, cioè
trasferimento da cellula a cellula di molecole segnale
nell’ambito delle diverse zone siano alla base del
mantenimento dei diversi domini cellulari nel doma e del
mantenimento della rispettiva organizzazione.
In mais l’espressione del gene KNOTTED1 (FT ad omeodominio del
tipo elica-ansa-elica) si correla positivamente alla formazione ed
attività del meristema caulinare. La proteina, come il suo mRNA, si
spostano da cellula a cellula attraverso i plasmodesmi.
Il gene di Arabidopsis omologo a KNOTTED1 di mais è SHOOT
MERISTEMLESS (STM). L’espressione di questo gene avviene in tutte
le cellule del doma caulinare tranne che nelle cellule interessate
all’organizzazione dei primordi fogliari e nei primordi.
STM è quindi un gene associato alle
cellule iniziali ed è marcatore
dell’attività
meristematica
delle
cellule del doma apicale.
La sua azione sembra quella di
impedire alle cellule della ZC di
partecipare all’organogenesi laterale.
Dal Libro di testo: Elementi di
Biologia dello Sviluppo delle
Piante, ed. EdiSES
MANTENIMENTO DELL’IDENTITA’ DI CELLULE
MERISTEMATICHE
I programmi di sviluppo sono regolati da reti di geni
interagenti.
WUSCHEL (WUS)
Nei mutanti wus loss of function l’attività meristematica
(SAM) cessa alla fine dell’embriogenesi e la crescita si arresta
allo stadio di cotiledoni.
Non viene mantenuta la popolazione delle cellule iniziali.
É stato IDENTIFICATO UN CIRCUITO REGOLATIVO CHE
COINVOLGE WUSCHEL (WUS) , SHOOTMERISTEMLESS
(STM) e CLAVATA (CLVI, CLV3).
STM non è solo, anche il FT ad
omeobox WUSCHEL (WUS) è
necessario per mantenere l’attività del
doma caulinare (come WOX5 nel CQ
della radice)
• L’espressione di WUS impedisce il
differenziamento delle cellule della ZC,
garantendo la persistenza di un gruppo di
cellule pluripotenti (staminali) e quindi la
crescita indefinita dell’apice vegetativo.
RADICE
Il meristema radicale diviene attivo nell’embrione allo
stato cordato. Se c’è un difetto nella formazione
dell’ipofisi il meristema radicale non si forma.
Infatti, in mutanti con aberranti modificazione a carico
dell’ipofisi non si ha la formazione del meristema
radicale.
La regione più centrale del meristema radicale è caratterizzata
dalla presenza di cellule con attività mitotica molto rallentata
Questa regione è detta CENTRO QUIESCENTE (QC).
Le cellule del centro quiescente, in condizioni normali, hanno un ciclo
mitotico molto rallentato ma in caso di danni all’apice meristematico
sono in grado di dividersi attivamente e così ripristinare il meristema
danneggiato.
Sembra quindi che il centro quiescente della radice rappresenti una
riserva di cellule da utilizzare al momento del bisogno.
Questo meccanismo di sostituzione è estremamente importante perché
l’apice della radice viene frequentemente danneggiato da agenti biotici
ed abiotici.
Caratteristiche citologiche del centro quiescente: contenuto di
RNA e sintesi proteica inferiore rispetto alle cellule circostanti.
Bassa attività del Golgi, creste mitocondriali poco sviluppate,
reticolo endoplasmatico poco sviluppato.
Esistono dei marcatori molecolari
d’identità del CQ
Ad
esempio
pAGL42::GFP
(fluorescent marker
of stem cell niche)
Nella radice di Arabidopsis tutte le cellule traggono origine dalle
cellule iniziali che circondano le 4 cellule fondatrici (CQ) e che
con queste formano la nicchia staminale.
Al di sopra del CQ è localizzato un
piccolo gruppo di cellule che sono le
iniziali
del
tessuto
vascolare,
circondate (in sezione trasversale) da
uno strato di iniziali del periciclo a
cui
seguono
le
iniziali
dell’endodermide e della corteccia e
lo strato più esterno è costituito dalle
iniziali di epidermide e cuffia laterale.
Al di sotto del CQ ci sono le iniziali
della columella.
v. Illustrazioni sul Libro di testo: Elementi
di Biologia dello Sviluppo delle Piante, ed.
EdiSES
L’auxina si muove nella pianta dall’apice caulinare verso la radice,
entra nelle cellule mediante i trasportatori AUX1 ed esce mediante
le proteine PIN.
In questo modo è stato osservato che in corrispondnza del
meristema apicale c’è un forte accumlo di auxina, in particolare
nelle cellule iniziali della columella e nel CQ.
• Come è stato detyerminato?
• Con il sistema DR5::GUS (v. Libro di testo
Elementi di Biologia dello Sviluppo delle
Piante, ed. EdiSES per la descrizione)
La determinazione del meristema apicale radicale (RAM) è
generata da alti livelli di IAA nella regione basale del
proembrione.
Elevati livelli di IAA sono causati dal trasporto polare di
IAA mediato dalle proteine PIN.
Il CQ inibisce il differenziamento delle cellule che lo
circondano e che quindi mantengono indefinitamente
caratteristiche di cellule staminali.
Il bilanciamento tra i segnali che impediscono il
differenziamento, provenienti dal CQ e segnali che
invece lo promuovono sono alla base di un corretto
funzionamento del meristema radicale.
Tutti i tessuti della radice prendono origine da
specifiche cellule staminali.
LE CELLULE INIZIALI MERISTEMATICHE
GENERANO LUNGHE FILE DI CELLULE.
IN TEORIA SI POTREBBE
RINTRACCIARE LA CELLULA
MERISTEMATICA DA CUI SI
ORIGINANO LE SINGOLE
CELLULE MATURE, perchè nella
radice le divisioni sono per lo più
anticlinali (trasversali all’asse
della radice) in modo da produrre
una crescita in lunghezza.
Poche divisioni periclinali invece
determinano l’aumento del
diametro della radice.
Le divisioni periclinali hanno luogo
soprattutto nell’apice radicale e
generano nuove file di cellule
Le cellule del centro quiescente producono segnali che
condizionano il comportamento delle iniziali circostanti.
Mediante esperimenti di
ablazione laser, è possibile
uccidere una determinata
cellula di un tessuto senza
distruggere l’intera radice.
È stato osservato che con
il
successivo
sviluppo
cellule circostanti sono in
grado di prendere il posto
delle cellule eliminate.
v.Elementi di
Biologia dello
Sviluppo delle
Piante, ed. EdiSES
Gli effetti dell’ablazione laser sulle cellule del
centro quiescente dimostrano come possa
essere acquisita una nuova identità cellulare
Le cellule procambiali
assumono per
transdifferenziamento
posizione e funzione di
cellule del centro
quiescente
Allora? Nella pianta, l’origine
cellulare non è poi così
importante!!!!!
• La cellula vegetale esprime il destino
appropriato alla sua nuova posizione, e
l’espressione genica relativa, nel caso
precedente, ad es., il gene specifico delle
cellule CQ: WUSCHELHOMEOBOX5
(WOX5)
Esperimenti di questo tipo sono stati utili anche per studiare la
comunicazione cellula-cellula nell’apice radicale e l’effetto
posizione.
È stato dimostrato, inoltre, che uccidendo una cellula del
CQ la cellula iniziale della columella (cellula derivata) più
vicina alla cellula eliminata non si divide più attivamente
(perde le caratteristiche di cellula iniziale) ma incomincia
subito a differenziare (accumulando di amido, che per la
columella rappresenta un marker del differenziamento). Questo
sta ad indicare che le cellule del CQ inviano segnali alle cellule
vicine che reprimono il precoce differenziamento della cuffia.
Ancora, le iniziali di corteccia ed endodermide vicine alla cellula
del CQ ablata smettono di dividersi trasversalmente e si
dividono longitudinalmente, in questo modo viene a mancare la
cellula che rimane iniziale e si formano direttamente la cellula
endodermica e corticale. Anche in questo caso sembra che le
cellule del CQ inviino segnali a breve raggio alle iniziali di E/C.
Il trasporto dell’auxina controlla anche il pattern distale del
differenziamento radicale.
L’auxina raggiunge l’apice radicale dal germoglio, si
accumula nel CQ e nelle iniziali della columella dopo di
che viene di nuovo trasportato verso l’alto in particolare
passando nelle cellule epidermiche. Questo percorso è
necessario per regolare il processo di distensione cellulare.
Le proteine PIN controllano la taglia della zona di divisione
dell’apice e regolano anche l’estensione della zona di
espansione/allungamento, attraverso una ridistribuzione
dell’auxina.
La zona di accrescimento per distenzione è seguita dalla ZONA
DI MATURAZIONE o DI DIFFERENZIAMENTO
CELLULARE.
In questa regione la maggior parte delle cellule dei tessuti primari
differenzia ed acquisisce la forma definitiva.
In corrispondenza dell’inizio di questa regione compaiono i PELI
radicali.
I peli radicali servono per aumentare sensibilmente la superficie
di assorbimento, infatti l’acqua ed i sali vengono assorbiti per la
maggior parte a livello dei peli radicali. Svolgono anche il ruolo di
protezione dell’organo.
Sono estroflessioni di alcune cellule epidermiche dette tricoblasti.
In questa regione, nella zona centrale si ha inoltre la definizione
della stele (differenziamento ed organizzazione delle cellule del
sistema vascolare).
I peli radicali hanno vita breve, pochi giorni, man mano che
muoiono vengono sostituiti da altri neo formati.
La velocità di formazione e
quella di degenerazione sono
simili per cui l’estensione della
zona pilifera resta costante nel
tempo. Non resta costante invece
la sua posizione rispetto al suolo.
Nella zona in cui i peli radicali
sono morti la funzione protettiva
viene svolta dagli strati più
esterni del parenchima corticale,
i quali suberificano (formando
esoderma, tessuto di origine
primaria)
I peli hanno una crescita apicale ed un grande vacuolo
centrale.
In alcune piante tutte le cellule epidermiche hanno la
possibilità di formare i peli, in altre, tra cui Arabidopsis solo
alcune possono formare i peli.
In Arabidopsis il controllo genico che dà alla cellula
epidermica la possibilità di formare un pelo è stato studiato
dettagliatamente anche con utilizzo di mutanti.
Atricoblastii (normali
celluledel rizoderma)
Tricoblasto (pelo radicale)
Con ablazione laser si
dimostra che quando un
atricoblasto viene
distrutto, il tricoblasto
vicino prende il suo
posto, manifestando poi
destino da atricoblasto,
e viceversa
TRASPARENT TESTA GLABRA1 (TTG1) è coinvolto nel destino atricoblastico nella radice (il
mutante fa solo tricoblasti), anche GLABRA2, FT ad omeodominio, è indispensabile per lo sviluppo
dell’atricoblasto ed è il bersaglio di TTG1 e di WEREWOLF (WER, FT di tipo MYB). CAPRICE (FT
di tipo MYB) (CPC) è coinvolto nel destino tricoblastico.
E’ stato dimostrato che CPC è un regolatore negativo dell’identità atricoblastica nella cellula in
cui si sposta (controllo genico per inibizione laterale).
Figure sul Libro di testo
le direzioni del differenziamento dei tessuti primari della
radice determinano i pattern distale e radiale
di sviluppo
I geni SHORT-ROOT (SHR) e SCARECROW (SCR) sono implicati nel
mantenere
la
corretta
organizzazione
del
parenchima
corticale/endodermide, mutanti per questi geni mostrano alterazioni
nella corteccia della radice primaria e delle laterali ed avventizie.
Il mutante scr ha un solo strato di parenchima corticale con
caratteristiche tipiche dell’endodermide, ad es. presenta la banda del
Caspary ma viene riconosciuto da un anticorpo che riconosce il
parenchima corticale e non l’endodermide. Nel mutante shr manca
l’endodermide
SCR e SHR sono fattori di
trascrizione della famiglia GRAS
i mutanti scr e shr hanno radice
corta, ed il CQ in scr è parzialmente
inattivo
Nei mutanti scr la cellula derivata non è in grado di fare
una divisione asimmetrica
periclina e si divide
trasversalmente. Il risultato è un solo strato cellulare con
caratteristiche intermedie fra endodermide e corteccia.
All’interno dell’endoderma della radice c’è il cilindro
centrale o stele.
Il primo tessuto della stele è il PERICICLO.
È mono o pluristratificato, è formato da cellule con
caratteristiche meristematiche.
È il tessuto da cui si originano le radici laterali o
secondarie.
Prende parte alla formazione del cambio cribro vascolare
e del cambio del sughero.
Deriva dal procambio e differenzia in prossimità
dell’apice radicale.
È stato denominato anche pericambio e gli è
attribuito il ruolo di meristema laterale.
stato
Una specifica espressione
genica nel periciclo induce la
formazione delle radici laterali.
Immagini dal Libro di
testo:
Elementi di Biologia dello
Sviluppo delle Piante, ed.
EdiSES
Queste prime divisioni sono sotto controllo auxinico. L’auxina
arriva al primordio laterale dall’apice della radice principale,
ma viene anche prodotta in loco.
L’importanza
dell’auxina
è
stata
dimostrata
sia
somministrando auxina esogena, particolari concentrazioni di
auxina portano alla produzione di un elevato numero di radici
laterali fino all’apice, sia somministrando inibitori del
trasporto auxinico, es. l'acido naftilftalamico (NPA), in
quest’ultimo caso si ha una fortissima riduzione del numero
di radici laterali.
Il ruolo dei PIN e la formazione del maximum auxinico
all’apice della radice laterale ha luogo come nella radice
primaria
Dal periciclo si formano le radici laterali ma anche il
cambio cribro-vascolare, in parte, e il fellogeno cioè i
meristemi secondari.
Le cellule del periciclo formano le radici laterali non
lontano dalla zona di distensione, i meristemi
secondari si formano in posizione molto lontano da
questa . È stato ipotizzato che un effetto posizionale
possa essere modulato nella diversificazione
dell’espressione genica nel periciclo.
Il floema e lo xilema primario della radice si formano dal
procambio. Il floema si differenzia più vicino all’apice, nella
regione di distensione, rispetto allo xilema.
Il sistema vascolare della radice si organizza durante le ultime
fasi dell’embriogenesi (orgnizzazione del procambio).
Le informazioni sul controllo genico del differenziamento
vascolare sono molto scarse.
È stato identificato un fattore di trascrizione di tipo MYB
necessario per il differenziamento del floema, ALTERED
PHLOEM DEVELOPMENT (APL). Nel mutante apl le
divisioni che dovrebbero dare origine a cellule floemaiche
sono pochissime e nella regione dove dovrebbe esserci il
floema si osservano cellule xilematiche.
.
Il gene ATHB-8 sembra essere un marcatore precoce
del differenziamento vascolare.
Si esprime nel procambio dell’embrione, radice, fusto
e foglia e nella regione di allungamento della radice
embrionale si esprime nelle cellule che differenziano
in xilema.
È regolato dall’auxina
Nonostante le informazioni sulla definizione del pattern
vascolare non siano molte, si sta affermando un’ipotesi
che ritiene che l’auxina e la citochinina siano implicate
entrambe nel controllo del differenziamento vascolare.
Secondo questo modello temporale l’auxina sarebbe
indispensabile per la formazione del tessuto vascolare,
ma la citochinina sarebbe necessaria prima, cioè per
attivare la proliferazione del procambio e per identificare
i siti di comparsa dei primi elementi di floema e xilema.
Domande:
•
•
•
•
•
•
•
•
•
Quali sono nell’apice caulinare i geni implicati nella formazione dei primordi
di foglie e gemme ascellari? Ed in che relazione sono con i geni della
“staminalità” del doma?
Nel doma caulinare quali geni sono implicati nel circuito regolativo della
nicchia staminale?
Nel doma radicale, a cosa servono le cellule del centro quiescente?
L’auxina ha un ruolo nel mantenimento della nicchia staminale della radice?
Quali cellule possono sostituirsi alle cellule del centro quiescente distrutte
assumendone l’identità?
Come e dove ha luogo il differenziamento dei peli radicali?
Quali geni sono essenziali per il differenziamento dell’endodermide?
Cos’è il periciclo e quanti e quali ruoli morfogenici assolve nella radice?
L’auxina ha un ruolo nel differenziamento delle radici laterali?