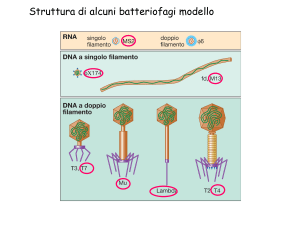
Struttura di alcuni batteriofagi modello
Il fago MU , un fago-trasposone.
Il DNA del fago Mu si integra in qualunque sito del genoma dell’ospite
come i trasposoni. Allo stato lisogeno determima quindi l’inattivazione nel
gene in cui si è inserito
Struttura di un fago temperato il fago MU
39 kb DNA a DS
le estremità contengono DNA dell’ospite ( circa 50-150 bp) a
destra e 1-2 kb a sinistra
MU è un fago/trasposone.
Una volta iniettato il DNA di MU si
integra con un meccanismo di
trasposizione conservativa mentre
durante l’infezione litica si
moltiplica con un meccanismo di
trasposizione replicativa
Integrazione del fago Mu nel cromosoma
Sia per il ciclo litico che
per il ciclo lisogenico il
fago MU si integra nel
cromosoma.
• L’integrazione è mediata dalla
trasposasi sintetizzata dal fago
• avviene un taglio sfalsato nel
Dna dell’ospite
•in seguito all’integrazione del
fago la polimerasi dell’ospite
replica i tratti a DNA SS
generando cosi duplicazioni
direttamente ripetute
all’estremità del fago ( 5 bp).
Il fago MU ha la potenzialità di codificare 2 tipi di fibre caudali
SU oppure S’U’ a seconda che il frammento che contiene questi geni
( frammento G ) si trovi nell’orientamento + o Dal momento che le fibre caudali servono per il riconoscimento di
proteine o macromolecole situate sulla parete cellulare del batterio
ospite se il frammeto G si trova nell’orientamento + il fago Mu potrà
infettare E.coli K12 se si trova nell’orientamento - potrà infettare
altri enterobatteri.
GIN
P
GIN
P
S
U
S’
U’
S’
Orient +
U’
U
S
Orient -
L’inversione del frammento G è mediata dalla proteina
MU-specifica Gin che agendo su delle brevi sequenze
omologhe situate all’estremità del segmento G
permette l’inversione del frammento.
Si tratta di un meccansimo di ricombinazione sito
specifica ( o illegittima) che richiede una proteina
specifica ( Gin) che agisce su sequenze localizzate
nelle adiacenze del gene gin.
Non è richiesto il sistema di ricombinazione
dell’ospite.
Il meccanismo è analogo a quello dell’inversione di
fase dei geni del flagellina in Salmonella ( flip-flop in
Salmonella)
MS2 fago virulento ad RNA SS molto piccolo
Genoma 3.5 kb ( T4 186 kb)
Proteine del pilo come recettore
Codifica solo 4 proteine :
Una sola proteina capsidica
Il capside costituito da 180 copie della proteina capsdica.
Non ha coda o fibre caudali
Codifica solo 4 proteine
La RNA replicasi ( gene P) è un complesso proteico ibrida che
contiene proteine dell’ospite oltre al polipeptide fago specifico
La proteina di maturazione ( gene A) determina il corretto
processamento di alcune proteine.
La proteina di lisi è codifica da un gene L sovrapposto sia al gene per la
proteina capsidica C che a quello della replicasi (geneA)
La proteina di maturazione A rimane associata
all’RNA nel virione ed è necessaria per l’iniezione del
RNA nella cellula batterica .
La regolazione della proteina A avviene a livello della
traduzione in quanto una lunga sequenza al 5’ UTR
del gene A forma una complessa struttura a
trifoglio mascherando ai ribosomi l’accesso alla
regione Shine –Delgarno. La struttura però viene a
formarsi lentamente permettendo cosi per un breve
lasso di tempo la sintesi della proteina.
Genoma a RNA a polarità+
• può essere utilizzato
direttamente per la
traduzione
•Può servire da stampo
per la sintesi di RNA a
polarità – da parte della
RNA replicasi
•Le molecole di RNA –
serviranno poi come
stampo per la sintesi di
numerose molecole a
polarità +
In MS2 il processo di traduzione controlla il n. di copie
delle 4 proteine.
AUG più disponibile per la traduzione è quello per la proteina capsidica
e per la replicasi che sono tradotte precocemente.
Ad alti livelli la proteina capsidica si associa all’RNA virale in
prossimità dell’AUG della replicasi impedendone la sintesi.
AUG della proteina di lisi è contenuto all’interno della sequenza della
proteina capsidica ed è difficilmente riconoscibile da parte dei
ribosomi per la complessa struttura secondaria che assume l’RNA virale.
Quando il ribosoma termina la traduzione della proteina capsidica l’AUG
per la sintesi della proteina litica diventa disponibile e la proteina viene
tradotta.
FX174 un fago a DNA SS a
polarità positiva.
FX174 è un piccolo batteriofago a
testa icosaedrica con un capside
costituito da 60 copie della stessa
proteina.
Ai vertici della struttura ad icosaedro del capside sono
presenti altre proteine delle spicole.
Genoma di 5.380 nucleotidi : fu il 1 genoma ad essere
sequenziato da Sanger ( premio Nobel)
Contiene numerosi geni sovrapposti: identificati per
la 1 volta in questo fago
Organizzazione del genoma del fago FX174
Presenza di numerosi geni sovrapposti
A e A* hanno la stessa fase di lettura ma due AUG
diversi
B è contenuto dentro A
K si sovrappone a A e C
E è all’interno di D
codone di stop di E è l’AUG di J
Replicazione di FX174
Il DNA a SS viene convertito in DS detta , Forma Replicativa (FR) da
•proteine dell’ospite,
•primasi ,
•DNA pol.I e III,
•Girasi e ligasi.
Una volta sintetizzati dalla primasi dei brevi inneschi di RNA, la DNA
polIII sintetizza il DNA fagico mentre la DNA polI sostituirà le
porzioni di RNA e la ligasi salderà i frammenti ( analogamente alla
sintesi del frammento copia).
Dalla FR viene sintetizzato
mRNA
altre copie a polarità + mediante il meccanismo del cerchio rotante


![Lezione 15 Virus [modalità compatibilità]](http://s1.studylibit.com/store/data/000771737_1-84b1cca561c5813066d1b76125338a98-300x300.png)