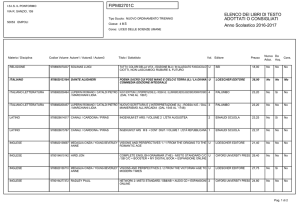
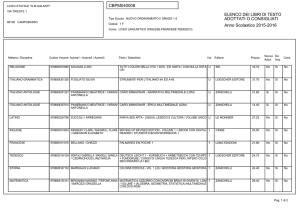
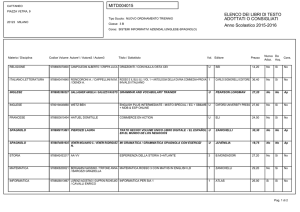
Capitolo 35
La struttura e le funzioni
delle piante
Copyright © 2006 Zanichelli editore
L’anatomia delle angiosperme
35.1 Le angiosperme si classificano nei due grandi
gruppi delle monocotiledoni e delle dicotiledoni
L’anatomia delle monocotiledoni e delle dicotiledoni
differisce nel numero di foglie embrionali, i cotiledoni, e
nella struttura delle radici, dei fusti, delle foglie e dei fiori.
Foglie embrionali
MONOCOTILEDONI
Un cotiledone
Nervature fogliari
Nervature quasi
sempre parallele
Fusti
Fasci vascolari
sparsi
Elementi fiorali
in genere
multipli di 3
Fasci vascolari
ad anello
Elementi fiorali
in genere
multipli di 4 o di
5
DICOTILEDONI
Figura 35.1
Copyright © 2006 Zanichelli editore
Due
cotiledoni
Nervature quasi
sempre ramificate
Fiori
Radici
Sistema radicale
fascicolato
Sistema radicale
a fittone
35.2 Il corpo di una pianta è costituito da un sistema
aereo e un sistema radicale
• Le radici formano il sistema radicale della pianta.
• L’apparato radicale àncora la pianta al suolo, assorbe
e trasporta i minerali e l’acqua, e immagazzina le
sostanze nutritive.
• Il sistema aereo della pianta comprende i fusti, le
foglie e gli organi per la riproduzione, che nelle
angiosperme sono i fiori.
• Il fusto è la parte della pianta che in genere si trova al
di sopra del suolo e che porta le foglie e i fiori.
Copyright © 2006 Zanichelli editore
Piano strutturale di un’angiosperma (dicotiledone):
Gemma apicale
Lamina
Foglia
Picciolo
Gemma ascellare
Sistema
aereo
Fiore
Fusto
Nodo
Internodo
Sistema
radicale
Figura 35.2
Copyright © 2006 Zanichelli editore
Fittone
Pelo
radicale
Pelo
radicale
Cellule
dell’epidermide
35.3 Molte piante presentano radici, fusti e foglie
modificati
Alcune piante (per esempio le carote, le rape, le
barbabietole da zucchero e i ravanelli) sono radici a fittone
particolarmente ingrossate, che contengono sostanze
nutritive sotto forma di carboidrati, come l’amido.
Figura 35.3A
Copyright © 2006 Zanichelli editore
In alcune piante, come le fragole, il fusto principale
produce fusti supplementari chiamati stoloni, che
crescono orizzontalmente sulla superficie del suolo e
permettono alla pianta di
Pianta di
riprodursi per via asessuata.
fragola
Pianta di
patata
Stolone
Pianta di zenzero
Fittone
Rizoma
Rizoma
Tubero
Radice
Figura 35.3B
Copyright © 2006 Zanichelli editore
Alcune piante hanno altri tipi di foglia modificata: come il
viticcio (nel pisello e nella vite) che permette alla pianta
di arrampicarsi e le spine (nei cactus) che proteggono la
pianta dagli erbivori.
Figura 35.3C
Copyright © 2006 Zanichelli editore
35.4 Le piante hanno cellule organizzate in tessuti e
specializzate per struttura e funzione
La maggior parte delle cellule vegetali presenta tre
strutture uniche tra le cellule eucariotiche:
• i cloroplasti, sede della fotosintesi;
• un vacuolo centrale ripieno di liquido, che
contribuisce a mantenere il turgore cellulare;
• una parete cellulare che circonda esternamente la
membrana plasmatica.
Copyright © 2006 Zanichelli editore
Struttura di una cellula vegetale:
Nucleo
Cloroplasto
Reticolo
endoplasmatico
Vacuolo
centrale
Pareti cellulari
Parete cellulare primaria
Parete
cellulare
secondaria
Lamella
mediana
Mitocondrio
Apparato di Golgi
Ribosomi
Microtubuli
Membrana plasmatica
Figura 35.4A
Copyright © 2006 Zanichelli editore
Pareti cellulari
di due cellule
Membrana adiacenti
plasmatica
Plasmodesmi
Punteggiatura
I cinque tipi principali di cellule vegetali sono:
• le cellule parenchimatiche;
• le cellule collenchimatiche;
• le cellule parenchimatiche;
• le cellule conduttrici della linfa grezza;
• le cellule conduttrici della linfa elaborata.
Copyright © 2006 Zanichelli editore
Le cellule parenchimatiche svolgono la maggior parte
delle funzioni metaboliche
LM 270
Parete
primaria
(sottile)
Punteggiatura
Vescicole contenenti amido
Figura 35.4B
Copyright © 2006 Zanichelli editore
Le cellule collenchimatiche forniscono sostegno alle
parti in crescita della pianta.
LM 270
Parete
primaria
(spessa)
Figura 35.4C
Copyright © 2006 Zanichelli editore
Le cellule sclerenchimatiche hanno una parete
cellulare secondaria resa rigida dalla presenza di lignina,
la principale componente chimica del legno, e
comprendono le fibre e le sclereidi.
Parete cellulare
secondaria
Scleridi
Punteggiature
Parete
cellulare
secondaria
Fibra
Figura 35.4D
Copyright © 2006 Zanichelli editore
Parete
cellulare
primaria
Punteggiature
Sclereide
LM 200
Parete cellulare
primaria
LM 266
Cellule
delle fibre
Le cellule conduttrici della linfa grezza (cioè acqua e
sali minerali) che comprendono le tracheidi (cellule
lunghe e affusolate) e gli elementi dei vasi (cellule più
larghe, più corte e meno affusolate).
Tracheidi
Punteggiature
Elemento dei
vasi
Punteggiature
Aperture nella
parete cellulare
Figura 35.4E
Copyright © 2006 Zanichelli editore
Colorizzata SEM 150
Le cellule conduttrici della linfa elaborata, i tubi cribosi,
hanno pareti primarie sottili, non possiedono parete
secondaria e restano vive anche a maturità.
Placca cribrosa
Cellula
compagna
Parete
cellulare
primaria
Figura 35.4F
Copyright © 2006 Zanichelli editore
Citoplasma
I due tipi di tessuto vascolare sono:
• lo xilema, che trasporta la linfa grezza (acqua e sali
minerali);
• Il floema, che trasporta la linfa elaborata (contenente
gli zuccheri).
Copyright © 2006 Zanichelli editore
35.5 Il corpo di una pianta è costituito da tre sistemi
di tessuti
Le componenti anatomiche di una pianta (radici, fusto
e foglie) sono costituite da tre sistemi di tessuti:
• il sistema tegumentale;
• il sistema del tessuto vascolare;
•
il sistema del tessuto fondamentale.
Copyright © 2006 Zanichelli editore
• Il tessuto tegumentale riveste e protegge la pianta.
• Il tessuto vascolare, contiene lo xilema e il floema;
le sue funzioni principali sono trasportare l’acqua e le
sostanze nutritive e fornire sostegno strutturale alla
pianta.
• Il sistema del tessuto fondamentale è costituito da
cellule parenchimatiche, cellule collenchimatiche di
sostegno e cellule sclerenchimatiche.
Copyright © 2006 Zanichelli editore
I sistemi
tegumentale,
fondamentale e
vascolare
Figura 35.5
Copyright © 2006 Zanichelli editore
L’accrescimento primario e secondario
35.6 L’accrescimento primario determina
l’allungamento delle radici e dei germogli
• In tutte le piante la crescita indeterminata è
possibile grazie alla presenza di particolari tessuti
detti meristemi.
• I meristemi sono regioni costituite da cellule
indifferenziate in divisione attiva, responsabili
dell’accrescimento primario della pianta.
Copyright © 2006 Zanichelli editore
• I meristemi apicali, posti all’apice radicale e nelle
gemme apicali e ascellari, crescono in lunghezza
producendo nuove cellule.
• L’allungamento continua nella radice o nel sistema
aereo mano a mano che le cellule si allungano e si
Gemma apicale
differenziano.
Gemme ascellari
Le frecce indicano la
direzione di crescita
Figura 35.6A
Copyright © 2006 Zanichelli editore
Apici radicali
L’estremità della radice
è rivestita dalla cuffia
radicale, una struttura
cellulare conica che
protegge le delicate
cellule del meristema
apicale in divisione
attiva.
Cilindro vascolare
Pelo radicale
Epidermide
Zona di differenziamento
Zona di allungamento
Fibre di
cellulosa
Regione del
meristema apicale
Legenda
Sistema tegumentale
Sistema fondamentale
Figura 35.6B
Copyright © 2006 Zanichelli editore
Corteccia
Sistema vascolare
Zona di
divisione
cellulare
Cuffia radicale
Foglie
Meristema
apicale
Meristemi delle
gemme ascellari
1
Figura 35.6C
Copyright © 2006 Zanichelli editore
2
LM 103
Sezione longitudinale dell’apice
di un germoglio in accrescimento:
gemma apicale tagliata fino a
incontrare la prima coppia di
gemme ascellari.
35.7 L’accrescimento secondario fa aumentare il
diametro delle piante legnose
L’aumento in diametro delle piante è invece dovuto
all’accrescimento secondario, che deriva dalla divisione
delle cellule di un meristema cilindrico chiamato cambio
vascolare.
Copyright © 2006 Zanichelli editore
Il cambio vascolare ispessisce il fusto aggiungendo strati
di xilema secondario (o legno) all’interno della sua
superficie.
Primo anno:
inizio della primavera
Secondo anno:
tarda estate
Primo anno:
tarda estate
Legenda
Sistema tegumentale
Sistema fondamentale
Sfaldamento
dell’epidermide
Xilema
primario
Epidermide
Cambio
vascolare
Corteccia
Floema
primario
Figura 35.7A
Copyright © 2006 Zanichelli editore
Xilema
secondario
(legno)
Floema
secondario
Sughero
Cambio del
sughero
Corteccia
esterna
Sistema vascolare
Xilema secondario
(accrescimento del
secondo anno)
• Il durame e l’alburno sono formati da diversi strati di xilema
secondario (più vecchi il primo, più giovani il secondo).
• Gli strati che si trovano all’esterno del cambio vascolare
(cioè il floema secondario, il cambio del sughero e il
sughero) costituiscono la corteccia esterna.
Alburno
Anelli
Raggi
midollari
Durame
Alburno
Cambio vascolare
Durame
Figura 35.7B
Copyright © 2006 Zanichelli editore
Corteccia
esterna
Floema secondario
Cambio del sughero
Sughero
L’assorbimento e il trasporto della linfa
35.8 Le piante prelevano sostanze nutritive dal suolo
e dall’aria
La respirazione e la liberazione di ossigeno
• Le piante assorbono dal suolo,
attraverso le radici, acqua e sali
minerali (ioni inorganici) e parte
dell’ossigeno.
• Le foglie prelevano invece
CO2 dall’aria.
CO2
Minerali
Figura 35.8A
Copyright © 2006 Zanichelli editore
H2 O
O2
La membrana plasmatica delle cellule radicali controlla
l’ingresso dei soluti.
I peli radicali aumentano grandemente la superficie di
assorbimento.
Figura 35.8B
Copyright © 2006 Zanichelli editore
L’acqua e i soluti
(linfa grezza) si
spostano
attraverso
l’epidermide e la
corteccia radicali
sia passando tra
le cellule, sia
attraversandole.
Peli
radicali
Epidermid
e
Floema
Corteccia
Banda di
Caspary
Xilema
Endoderma
Via extracellulare
Banda di
Caspary
Xilema
Pelo radicale
Legenda
Sistema tegumentale
Sistema fondamentale
Sistema vascolare
Plasmodesmi
Via intracellulare
Epidermid
e
Figura 35.8C
Copyright © 2006 Zanichelli editore
Endoderma
Corteccia
In tutti i casi, la linfa grezza deve passare attraverso la
membrana selettivamente permeabile delle cellule
dell’endoderma per penetrare nello xilema e andare
verso l’alto.
Copyright © 2006 Zanichelli editore
35.9 Lo xilema trasporta acqua e sali minerali dalle
radici al resto della pianta
Linfa grezza
Cellule del mesofillo
Spazi aerei nella foglia
Stoma
Aria esterna
La traspirazione aspira la
alle foglie più alte.
Traspirazione
Flusso di acqua
linfa grezza dalle radici
Adesione
Parete
cellulare
Molecole
d’acqua
Cellule
Coesione e adesione dello
xilema
nello xilema
Coesione
dovuta ai
legami
idrogeno
Pelo radicale
Particella
di terreno
Acqua
Figura 35.9A
Copyright © 2006 Zanichelli editore
Assorbimento di
acqua dal suolo
• Le cellule di guardia controllano la traspirazione.
• L’apertura e la chiusura degli stomi delle foglie
sono il risultato di un adattamento che consente
alle piante di regolare il loro contenuto di acqua,
adeguandosi alle variazioni delle condizioni
ambientali.
Copyright © 2006 Zanichelli editore
Ciascuno stoma è delimitato da una coppia di cellule di
guardia, che ne determinano l’apertura o la chiusura
cambiando forma.
Stoma
H2O
Cellule di guardia
K
Vacuolo
+
H2O
H2O
H2O
H2O
Lo stoma si apre
Copyright © 2006 Zanichelli editore
H2O
H2O
H2O
Figura 35.9B
H2O
H2O
Lo stoma si chiude
35.10 Il floema trasporta i prodotti della fotosintesi
nella linfa elaborata
Il floema è costituito da cellule, chiamate elementi dei
tubi cribosi, sovrapposte in modo continuo a formare
delle colonne.
Elemento
del tubo
criboso
Figura 35.10A
Copyright © 2006 Zanichelli editore
TEM 2700
Placca
cribosa
Il floema trasporta le molecole nutritive prodotte con la
fotosintesi per un meccanismo di flusso di pressione.
Floema
Concentrazione di zuccheri elevata
Pressione idrostatica elevata 1
Sorgente
Xilema
1
Zucchero
2
Acqua
2
Cellula della sorgente
Placca cribosa
Cellula del
pozzo
Pozzo
3
3
Zucchero
4
4
Bassa concentrazione di zuccheri
Figura 35.10B
Copyright © 2006 Zanichelli editore
Bassa pressioneLow
idrostatica
water
pressure
Acqua
• A livello della sorgente (per esempio, nelle foglie)
gli zuccheri passano nei tubi cribosi del floema per
trasporto attivo.
• Questo apporto di zuccheri fa aumentare la
concentrazione di soluti nel fleoma.
• L’alta concentrazione di soluti richiama per osmosi
acqua nei tubi cribosi.
• A livello del pozzo sia i soluti sia l’acqua escono dal
tubo criboso; gli zuccheri lasciano il floema.
Copyright © 2006 Zanichelli editore
• L’aumento della pressione idrostatica alla sorgente
e la sua diminuzione al pozzo fanno sì che l’acqua
scorra dalla sorgente al pozzo, lungo il gradiente.
• Poiché gli zuccheri sono sciolti nell’acqua vengono
trasportati dalla sorgente al pozzo alla stessa
velocità dell’acqua.
Copyright © 2006 Zanichelli editore
Le sostanze nutritive necessarie alle piante
35.11 Il benessere delle piante dipende da diverse
sostanze nutritive inorganiche
• Le piante vivono e crescono utilizzando
esclusivamente sostanze inorganiche.
• Un elemento chimico è da considerarsi essenziale
per le piante se è indispensabile per il
completamento del loro ciclo vitale.
Copyright © 2006 Zanichelli editore
Per scoprire se un determinato elemento, per esempio il
potassio, è indispensabile per la pianta, questa viene
coltivata su un terreno privo di quell’elemento.
Figura 35.11A
Copyright © 2006 Zanichelli editore
Soluzione nutritiva completa,
Soluzione priva di
contenente tutti i minerali
potassio (sperimentale)
(controllo)
• Per le piante sono macronutrienti quegli elementi
(azoto, fosforo e potassio) necessari in grandi
quantità, usati principalmente per la sintesi delle
molecole organiche.
• I micronutrienti (tra cui ferro e zinco) agiscono
principalmente come cofattori di enzimi.
Copyright © 2006 Zanichelli editore
Le carenze di nutrienti sono diagnosticabili secondo
segni di sofferenza mostrati nelle piante.
Figura 35.11B
Copyright © 2006 Zanichelli editore
35.12 La fertilità del suolo è fondamentale per la
crescita delle piante e per la vita umana
• Un suolo si definisce fertile quando consente la
crescita di un numero abbondante di piante.
• Il suolo fertile contiene una miscela di particelle
rocciose e argillose che trattiene acqua e ioni
minerali e che permette all’ O2 di diffondersi nelle
radici delle piante.
Copyright © 2006 Zanichelli editore
• Il suolo è costituito da strati distinti chiamati
orizzonti.
• Il suolo superficiale è una miscela di particelle di
roccia di vario calibro, organismi viventi e humus
(costituito da resti di materiale organico in
decomposizione, fornisce le sostanze nutritive e
sostiene la vita di organismi che contribuiscono alla
fertilità del terreno).
Figura 35.12A
Copyright © 2006 Zanichelli editore
Tra i peli radicali delle piante, l’acqua del suolo e le
particelle del suolo superficiale esiste una stretta
associazione.
Particella di suolo circondata
da una pellicola d’acqua
Pelo radicale
Acqua
Spazio contenente aria
Figura 35.12B
Copyright © 2006 Zanichelli editore
• I cationi (ioni a carica positiva), come il K+,
aderiscono alle particelle del suolo.
• Nel meccanismo dello scambio cationico, i peli
radicali rilasciano ioni H+, che si scambiano con
cationi del suolo, che possono quindi essere
assorbiti dalle radici.
K+
K+
K+
Particella di
argilla
H+
K+
K+
K+
K+
K+
Pelo radicale
Figura 35.12C
Copyright © 2006 Zanichelli editore
• Gli anioni (ioni a carica negativa), come i nitrati
NO3-, sono prontamente disponibili per le piante
poiché non sono legati alle particelle del suolo.
• Tuttavia, gli anioni tendono a essere dilavati
facilmente.
Copyright © 2006 Zanichelli editore
• Conservare la fertilità del suolo è importante per
l’umanità.
• L’irrigazione oculata, il controllo dell’erosione e
l’uso prudente di fertilizzanti e pesticidi sono aspetti
utili al mantenimento della fertilità dei suoli.
Figura 35.12D
Figura 35.12E
Copyright © 2006 Zanichelli editore
COLLEGAMENTI
35.13 Due approcci opposti: l’agricoltura biologica e
la genetica applicata
L’agricoltura biologica, o organica, applica i principi
dell’ecologia alle pratiche agronomiche e comprende
metodi colturali che tendono a escludere i prodotti
chimici di sintesi.
Figura 35.13A
Copyright © 2006 Zanichelli editore
• La ricerca biologica in agricoltura si pone come uno
dei principali obiettivi l’aumento qualitativo e
quantitativo del contenuto proteico dei raccolti.
• Con le biotecnologie i ricercatori creano varietà di
piante geneticamente modificate (GM), con le
caratteristiche desiderate.
Figura 35.13B
Copyright © 2006 Zanichelli editore
Simbiosi e modalità nutritive delle piante
35.14 I funghi aiutano la maggior parte delle piante
ad assorbire le sostanze nutritive del suolo
• Diversi tipi di relazioni con altri organismi
contribuiscono alla nutrizione delle piante.
• Molte piante formano micorrize, cioè associazioni
reciprocamente benefiche coni funghi.
Figura 35.14
Copyright © 2006 Zanichelli editore
SEM 158
• Una rete di filamenti fungini circonda le radici,
aumentandone le capacità di assorbimento d’acqua e
nutrienti. Il fungo, in cambio, ottiene nutrienti dalla
pianta.
35.15 La maggior parte delle piante dipende dai
batteri per l’approvvigionamento di azoto
I batteri azotofissatori del suolo convertono l’azoto
atmosferico N2 in forme utilizzabili dalle piante, in un
processo metabolico chiamato fissazione dell’azoto.
ATMOSFERA
N2
Aminoacidi
N2
Batteri
azotofissatori
NH4
H+
+
Suolo
NH3
Materia
organica
Figura 35.15A
Copyright © 2006 Zanichelli editore
Batteri
ammonificanti
NH4
+
(ioni ammonio) Batteri
nitrificanti
NO3–
(nitrato)
Radice
• Le radici delle leguminose presentano noduli radicali
contenenti batteri azotofissatori del genere Rhizobium.
• Tra e piante e i batteri esiste una relazione di reciproco
vantaggio: la pianta fornisce ai batteri carboidrati e altri
composti organici, i batteri producono enzimi che catalizzano la
trasformazione dell’azoto atmosferico in ioni ammonio.
Batteri all’interno
di vescicole
Noduli
Radici
Figura 35.15B
Copyright © 2006 Zanichelli editore
Figura 35.15C
TEM 5850
Fusto
35.16 Metodi di nutrizione alternativi: le piante
parassite e le piante carnivore
• Le piante parassite succhiano la linfa elaborata da
altre piante.
• Le piante carnivore completano
la loro nutrizione azotata
catturando insetti e altri piccoli
animali con vari sistemi.
Figura 35.16B
Figura 35.16A
Copyright © 2006 Zanichelli editore
Figura 35.16C
Figura 35.16D
I sistemi di controllo ormonale
35.17 Alcuni storici esperimenti sui movimenti delle
piante verso la luce hanno portato alla scoperta del
primo ormone vegetale
Il fototropismo è una risposta adattativa grazie alla quale i
germogli in crescita e i fusti delle piante adulte si dirigono
verso la luce del Sole, di cui hanno bisogno per effettuare la
fotosintesi.
Figura 35.17A
Copyright © 2006 Zanichelli editore
Esperimenti sulle piante hanno portato alla scoperta di
un messaggero chimico, l’ormone auxina, responsabile
del fototropismo che può essere dovuto agli spostamenti
dell’auxina dal lato illuminato al lato buio del fusto.
Lato al
buio del
germoglio
Luce
Lato illuminato
del germoglio
Figura 35.17B
Copyright © 2006 Zanichelli editore
35.18 Gli ormoni vegetali sono sostanze che
regolano la crescita e lo sviluppo delle piante
• Gli ormoni vegetali coordinano le attività delle
cellule e dei tessuti della pianta.
• Questi ormoni sono prodotti in piccole quantità in
certi distretti della pianta e conducono segnali ad
altri distretti, regolando la crescita e lo sviluppo.
Copyright © 2006 Zanichelli editore
Le piante producono cinque classi principali di ormoni:
auxina, citochinine, giberelline, acido abscissico ed
etilene.
Tabella 35.18
Copyright © 2006 Zanichelli editore
35.19 Le auxine stimolano l’allungamento delle
cellule nei giovani germogli
• Il termine auxina viene utilizzato per descrivere
una classe di composti la cui funzione principale è
quella di promuovere l’allungamento dei germogli
in via di sviluppo.
• L’auxina (acido indolacetico, IAA) è prodotta agli
apici meristematici radicali e delle gemme apicali.
Copyright © 2006 Zanichelli editore
Allungamento
Stimolazione
Inibizione
A varie concentrazioni, l’auxina stimola o inibisce
l’allungamento dei fusti e delle radici.
Fusti
0
Radici
0,9 g/L
10–8
10–6
10–4
10–2
1
Aumento della concentrazione di auxina (g/L)
Figure 35.19A, B
Copyright © 2006 Zanichelli editore
102
• L’auxina può agire indebolendo le pareti cellulari,
stimolando alcune proteine di membrana a pompare
ioni idrogeno all’interno della parete stessa.
• Tali ioni attivano altri enzimi che rompono i legami tra
le molecole di cellulosa della parete, facendo
assorbire acqua alla cellula.
• L’auxina stimola anche lo sviluppo dei tessuti
vascolari e la divisione cellulare nel cambio
vascolare, promuovendo la crescita in diametro del
fusto.
Copyright © 2006 Zanichelli editore
35.20 Le citochinine stimolano la divisione cellulare
• Le citochinine sono regolatori della crescita che
promuovono la divisione cellulare, detta anche
citodieresi (o citochinesi).
• Le citochinine naturali sono prodotte nei tessuti in
crescita attiva, in particolare nelle radici, negli
embrioni e nei frutti.
Copyright © 2006 Zanichelli editore
Le citochinine di origine radicale bilanciano gli effetti
dell’auxina prodotta dai meristemi apicali, provocando lo
sviluppo delle gemme laterali e quindi la ramificazione.
Gemma apicale presente
Gemma apicale assente
Figura 35.20
Copyright © 2006 Zanichelli editore
35.21 Le gibberelline influiscono sull’allungamento
del fusto
Le gibberelline stimolano l’allungamento dei fusti e delle
foglie.
Figura 35.21A
Copyright © 2006 Zanichelli editore
• Le gibberelline stimolano anche lo sviluppo dei
frutti.
• Quelle rilasciate dagli embrioni sono coinvolte negli
eventi che accompagnano le prime fasi della
germinazione dei semi.
Figura 35.21B
Copyright © 2006 Zanichelli editore
35.22 L’acido abscissico inibisce molti processi
vegetali
• L’acido abscissico (AB) inibisce la germinazione
dei semi.
• Il rapporto ABA/gibberelline spesso determina la
quiescenza o la germinazione dei semi.
Copyright © 2006 Zanichelli editore
• L’ABA funge anche da ormone dello stress, causando
la chiusura degli stomi in caso di disidratazione.
• I semi di alcune piante restano quiescenti finché il
contenuto in ABA non viene dilavato dall’acqua o non
viene inattivato.
Figura 35.22
Copyright © 2006 Zanichelli editore
35.23 L’etilene induce la maturazione dei frutti e
controlla i processi di invecchiamento
• Quando le cellule di un frutto maturano, producono
etilene, un gas che favorisce la maturazione.
• Agendo come un ormone, l’etilene induce diversi
tipi di risposte connesse con l’invecchiamento tra
cui la morte programmata delle cellule.
Copyright © 2006 Zanichelli editore
La maturazione dei frutti
Un aumento della produzione di etilene in un frutto ne
innesca la maturazione.
1
3
2
Figura 35.23A
Copyright © 2006 Zanichelli editore
Gambo
della foglia
La caduta delle foglie
Ramo
LM 20
Un cambiamento del rapporto
tra auxina ed etilene,
innescato soprattutto
dall’accorciarsi del numero di
ore di luce, probabilmente
provoca i cambiamenti tipici
dell’autunno negli alberi
decidui.
Stato protettivo
Figura 35.23B
Copyright © 2006 Zanichelli editore
Ramo
Stato di abscissione
Picciolo
della foglia
I tropismi e i ritmi biologici
35.24 I tropismi sono meccanismi che permettono
alle piante di rispondere agli stimoli dell’ambiente
• Le piante sono in grado di percepire e di
rispondere ai cambiamenti ambientali in diversi
modi.
• I tropismi sono risposte di crescita che mutano la
forma di una pianta o la fanno crescere verso uno
stimolo o lontano da esso.
Copyright © 2006 Zanichelli editore
La risposta alla luce
• Il fototropismo dà luogo a una diversa velocità di
allungamento delle cellule sui lati opposti del fusto.
• A causa della diversa distribuzione di auxina, le
cellule sul lato non illuminato del fusto si allungano
più velocemente di quelle poste sul lato che riceve
luce diretta.
Copyright © 2006 Zanichelli editore
La risposta alla gravità
• Il geotropismo è una risposta alla gravità e può
essere causato dagli spostamenti di certi organuli
sul lato inferiore del fusto e delle radici.
• Lo spostamento degli organuli può causare un
cambiamento nella distribuzione degli ormoni.
Figura 35.24A
Copyright © 2006 Zanichelli editore
La risposta al contatto
Il tigmotropismo è la risposta al contatto ed è
responsabile dell’avvolgimento dei viticci e dei
rampicanti intorno a oggetti.
Figura 35.24B
Copyright © 2006 Zanichelli editore
35.25 Le piante hanno orologi interni e percepiscono
le stagioni
I ritmi biologici innati e l’adeguamento ai ritmi ambientali
• Un orologio biologico interno aita le piante a controllare i
movimenti notturni e altri cicli giornalieri.
• Questi cicli, chiamati ritmi circadiani, durano 24 ore e
persistono anche in assenza di stimoli ambientali.
Figura 35.25
Mezzogiorno
Copyright © 2006 Zanichelli editore
Mezzanotte
Il fotoperiodo e i cicli stagionali
• Le piante percepiscono le stagioni misurando il
fotoperiodo, la lunghezza relativa delle ore di buio
e di luce.
• Il momento della fioritura è una delle risposte
stagionali al fotoperiodo.
Copyright © 2006 Zanichelli editore
Le piante in cui la fioritura è indotta dal fotoperiodo si
dividono in due gruppi:
• piante brevidiurne che generalmente fioriscono alla
fine dell’estate, in autunno o in inverno, quando il
numero delle ore di luce è minore di quelle di buio.
• piante longidiurne che fioriscono alla fine della
primavera o all’inizio dell’estate, quando il numero
delle ore di luce è maggiore.
Copyright © 2006 Zanichelli editore